
SCF Ligases and Their Functions in Oogenesis and Embryogenesis—Summary of the Most Important Findings throughout the Animal Kingdom


Abstract
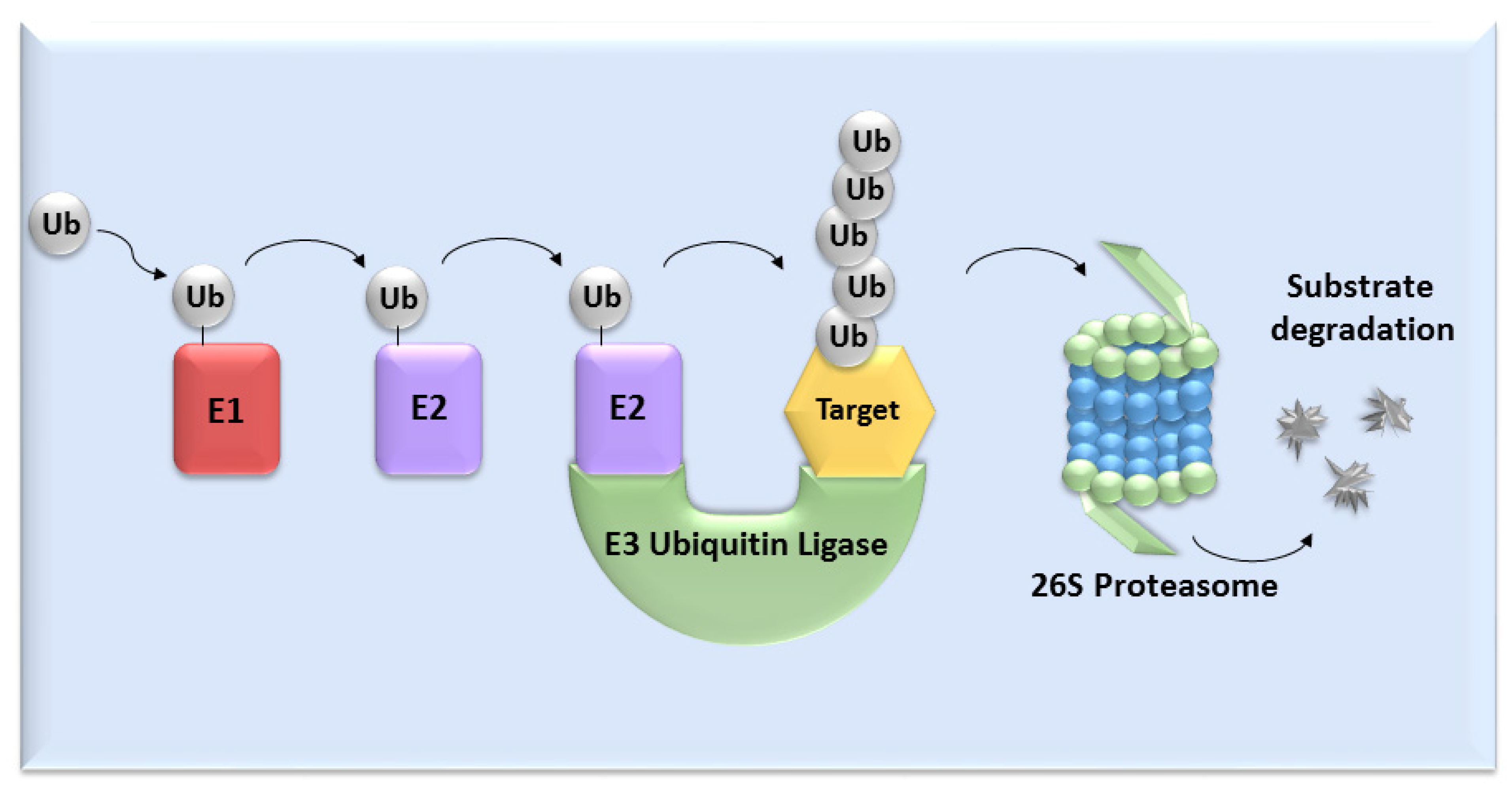
:1. Introduction
2. SCF Ligases Composition
2.1. Cullins
2.2. RBX1 (Ring-Box 1)
2.3. SKP1 (S-Phase Kinase-Associated Protein 1)
2.4. F-Box Proteins
3. SCF Ligases in Oocyte Maturation
4. SCF Ligases in Embryogenesis
5. Conclusions/Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez, F.; Smitz, J. Molecular Control of Oogenesis. Biochim. Biophys. Acta 2012, 1822, 1896–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swain, J.E.; Pool, T.B. ART Failure: Oocyte Contributions to Unsuccessful Fertilization. Hum. Reprod. Update 2008, 14, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Kline, D.; Kline, J.T. Repetitive Calcium Transients and the Role of Calcium in Exocytosis and Cell Cycle Activation in the Mouse Egg. Dev. Biol. 1992, 149, 80–89. [Google Scholar] [CrossRef]
- Laurincík, J.; Hyttel, P.; Baran, V.; Eckert, J.; Lucas-Hahn, A.; Pivko, J.; Niemann, H.; Brem, G.; Schellander, K. A Detailed Analysis of Pronucleus Development in Bovine Zygotes in Vitro: Cell-Cycle Chronology and Ultrastructure. Mol. Reprod. Dev. 1998, 50, 192–199. [Google Scholar] [CrossRef]
- Jukam, D.; Shariati, S.A.M.; Skotheim, J.M. Zygotic Genome Activation in Vertebrates. Dev. Cell 2017, 42, 316–332. [Google Scholar] [CrossRef]
- Minami, N.; Suzuki, T.; Tsukamoto, S. Zygotic Gene Activation and Maternal Factors in Mammals. J. Reprod. Dev. 2007, 53, 707–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, F.; Worrad, D.M.; Schultz, R.M. Regulation of Transcriptional Activity during the First and Second Cell Cycles in the Preimplantation Mouse Embryo. Dev. Biol. 1997, 181, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanka, J.; Kepková, K.; Nemcová, L. Gene Expression during Minor Genome Activation in Preimplantation Bovine Development. Theriogenology 2009, 72, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Barckmann, B.; Simonelig, M. Control of Maternal MRNA Stability in Germ Cells and Early Embryos. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2013, 1829, 714–724. [Google Scholar] [CrossRef] [PubMed]
- Glickman, M.H.; Ciechanover, A. The Ubiquitin-Proteasome Proteolytic Pathway: Destruction for the Sake of Construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef] [PubMed]
- Zheng, N.; Shabek, N. Ubiquitin Ligases: Structure, Function, and Regulation. Annu. Rev. Biochem 2017, 86, 129–157. [Google Scholar] [CrossRef]
- Petroski, M.D.; Deshaies, R.J. Function and Regulation of Cullin-RING Ubiquitin Ligases. Nat. Rev. Mol. Cell Biol. 2005, 6, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.; Wu, K.; Yang, Y.; Guerrero, C.; Nillegoda, N.; Pan, Z.-Q.; Huang, L. A Targeted Proteomic Analysis of the Ubiquitin-like Modifier Nedd8 and Associated Proteins. J. Proteome Res. 2008, 7, 1274–1287. [Google Scholar] [CrossRef] [Green Version]
- Bennett, E.J.; Rush, J.; Gygi, S.P.; Harper, J.W. Dynamics of Cullin-RING Ubiquitin Ligase Network Revealed by Systematic Quantitative Proteomics. Cell 2010, 143, 951–965. [Google Scholar] [CrossRef] [Green Version]
- Kepkova, K.V.; Vodicka, P.; Toralova, T.; Lopatarova, M.; Cech, S.; Dolezel, R.; Havlicek, V.; Besenfelder, U.; Kuzmany, A.; Sirard, M.-A.; et al. Transcriptomic Analysis of in Vivo and in Vitro Produced Bovine Embryos Revealed a Developmental Change in Cullin 1 Expression during Maternal-to-Embryonic Transition. Theriogenology 2011, 75, 1582–1595. [Google Scholar] [CrossRef]
- Benesova, V.; Kinterova, V.; Kanka, J.; Toralova, T. Characterization of SCF-Complex during Bovine Preimplantation Development. PLoS ONE 2016, 11, e0147096. [Google Scholar] [CrossRef] [Green Version]
- Ying, C.; Yangsheng, W.; Jiapeng, L.; Liqin, W.; Xiaolin, L.; Mingjun, L.; Juncheng, H. Transcriptome Profiles of Pre-Pubertal and Adult in Vitro Matured Ovine Oocytes Obtained from FSH-Stimulated Animals. Reprod. Domest. Anim. 2021, 56, 1085–1094. [Google Scholar] [CrossRef]
- Dealy, M.J.; Nguyen, K.V.; Lo, J.; Gstaiger, M.; Krek, W.; Elson, D.; Arbeit, J.; Kipreos, E.T.; Johnson, R.S. Loss of Cul1 Results in Early Embryonic Lethality and Dysregulation of Cyclin E. Nat. Genet. 1999, 23, 245–248. [Google Scholar] [CrossRef]
- Wang, Y.; Penfold, S.; Tang, X.; Hattori, N.; Riley, P.; Harper, J.W.; Cross, J.C.; Tyers, M. Deletion of the Cul1 Gene in Mice Causes Arrest in Early Embryogenesis and Accumulation of Cyclin E. Curr. Biol. 1999, 9, 1191–1194. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Luo, T.; He, L.; Tan, Y.; Zhang, Q. New Insights into the Roles of CUL1 in Mouse Placenta Development. Biochem. Biophys. Res. Commun. 2021, 559, 70–77. [Google Scholar] [CrossRef]
- Sun, X.; Tong, X.; Hao, Y.; Li, C.; Zhang, Y.; Pan, Y.; Dai, Y.; Liu, L.; Zhang, T.; Zhang, S. Abnormal Cullin1 Neddylation-Mediated P21 Accumulation Participates in the Pathogenesis of Recurrent Spontaneous Abortion by Regulating Trophoblast Cell Proliferation and Differentiation. Mol. Hum. Reprod 2020, 26, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yu, S.; Huang, X.; Tan, Y.; Zhu, C.; Wang, Y.-L.; Wang, H.; Lin, H.-Y.; Fu, J.; Wang, H. New Insights into the Function of Cullin 3 in Trophoblast Invasion and Migration. Reproduction 2015, 150, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Tsunematsu, R.; Nishiyama, M.; Kotoshiba, S.; Saiga, T.; Kamura, T.; Nakayama, K.I. Fbxw8 Is Essential for Cul1-Cul7 Complex Formation and for Placental Development. Mol. Cell. Biol. 2006, 26, 6157–6169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, M.; Nishida, T.; Nagayama, Y.; Yasuda, H. Nedd8-Modification of Cul1 Is Promoted by Roc1 as a Nedd8-E3 Ligase and Regulates Its Stability. Biochem. Biophys. Res. Commun. 2003, 301, 392–398. [Google Scholar] [CrossRef]
- Ohta, T.; Michel, J.J.; Schottelius, A.J.; Xiong, Y. ROC1, a Homolog of APC11, Represents a Family of Cullin Partners with an Associated Ubiquitin Ligase Activity. Mol. Cell 1999, 3, 535–541. [Google Scholar] [CrossRef]
- Jia, L.; Sun, Y. RBX1/ROC1-SCF E3 Ubiquitin Ligase Is Required for Mouse Embryogenesis and Cancer Cell Survival. Cell Div. 2009, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Sasagawa, Y.; Urano, T.; Kohara, Y.; Takahashi, H.; Higashitani, A. Caenorhabditis Elegans RBX1 Is Essential for Meiosis, Mitotic Chromosomal Condensation and Segregation, and Cytokinesis. Genes Cells 2003, 8, 857–872. [Google Scholar] [CrossRef]
- Jia, L.; Bickel, J.S.; Wu, J.; Morgan, M.A.; Li, H.; Yang, J.; Yu, X.; Chan, R.C.; Sun, Y. RBX1 (RING Box Protein 1) E3 Ubiquitin Ligase Is Required for Genomic Integrity by Modulating DNA Replication Licensing Proteins. J. Biol. Chem. 2011, 286, 3379–3386. [Google Scholar] [CrossRef] [Green Version]
- Noureddine, M.A.; Donaldson, T.D.; Thacker, S.A.; Duronio, R.J. Drosophila Roc1a Encodes a RING-H2 Protein with a Unique Function in Processing the Hh Signal Transducer Ci by the SCF E3 Ubiquitin Ligase. Dev. Cell 2002, 2, 757–770. [Google Scholar] [CrossRef] [Green Version]
- Bai, C.; Sen, P.; Hofmann, K.; Ma, L.; Goebl, M.; Harper, J.W.; Elledge, S.J. SKP1 Connects Cell Cycle Regulators to the Ubiquitin Proteolysis Machinery through a Novel Motif, the F-Box. Cell 1996, 86, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.W.; Eletsky, A.; Gonzalez, K.J.; van der Wel, H.; Strauch, E.-M.; Prestegard, J.H.; West, C.M. Skp1 Dimerization Conceals Its F-Box Protein Binding Site. Biochemistry 2020, 59, 1527–1536. [Google Scholar] [CrossRef]
- Guan, Y.; Leu, N.A.; Ma, J.; Chmátal, L.; Ruthel, G.; Bloom, J.C.; Lampson, M.A.; Schimenti, J.C.; Luo, M.; Wang, P.J. SKP1 Drives the Prophase I to Metaphase I Transition during Male Meiosis. Sci. Adv. 2020, 6, eaaz2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandel, S.A.; Fishman-Jacob, T.; Youdim, M.B.H. Targeting SKP1, an Ubiquitin E3 Ligase Component Found Decreased in Sporadic Parkinson’s Disease. Neurodegener. Dis. 2012, 10, 220–223. [Google Scholar] [CrossRef]
- Piva, R.; Liu, J.; Chiarle, R.; Podda, A.; Pagano, M.; Inghirami, G. In Vivo Interference with Skp1 Function Leads to Genetic Instability and Neoplastic Transformation. Mol. Cell. Biol. 2002, 22, 8375–8387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, P.K.; Eldridge, A.G.; Freed, E.; Furstenthal, L.; Hsu, J.Y.; Kaiser, B.K.; Reimann, J.D. The Lore of the RINGs: Substrate Recognition and Catalysis by Ubiquitin Ligases. Trends Cell Biol. 2000, 10, 429–439. [Google Scholar] [CrossRef]
- Galan, J.M.; Peter, M. Ubiquitin-Dependent Degradation of Multiple F-Box Proteins by an Autocatalytic Mechanism. Proc. Natl. Acad. Sci. USA 1999, 96, 9124–9129. [Google Scholar] [CrossRef] [Green Version]
- Kisielnicka, E.; Minasaki, R.; Eckmann, C.R. MAPK Signaling Couples SCF-Mediated Degradation of Translational Regulators to Oocyte Meiotic Progression. Proc. Natl. Acad. Sci. USA 2018, 115, E2772–E2781. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Yang, M.; Gao, C.; Yue, W.; Liang, X.; Xie, B.; Zhu, X.; Fan, S.; Li, R.; Li, M. Fbxo30 Regulates Chromosome Segregation of Oocyte Meiosis. Cell. Mol. Life Sci. 2019, 76, 2217–2229. [Google Scholar] [CrossRef]
- Zhao, B.-W.; Sun, S.-M.; Xu, K.; Li, Y.-Y.; Lei, W.-L.; Li, L.; Liu, S.-L.; Ouyang, Y.-C.; Sun, Q.-Y.; Wang, Z.-B. FBXO34 Regulates the G2/M Transition and Anaphase Entry in Meiotic Oocytes. Front. Cell Dev. Biol. 2021, 9, 647103. [Google Scholar] [CrossRef]
- Margottin-Goguet, F.; Hsu, J.Y.; Loktev, A.; Hsieh, H.M.; Reimann, J.D.R.; Jackson, P.K. Prophase Destruction of Emi1 by the SCF(BetaTrCP/Slimb) Ubiquitin Ligase Activates the Anaphase Promoting Complex to Allow Progression beyond Prometaphase. Dev. Cell 2003, 4, 813–826. [Google Scholar] [CrossRef] [Green Version]
- Marangos, P.; Verschuren, E.W.; Chen, R.; Jackson, P.K.; Carroll, J. Prophase I Arrest and Progression to Metaphase I in Mouse Oocytes Are Controlled by Emi1-Dependent Regulation of APC(Cdh1). J. Cell Biol. 2007, 176, 65–75. [Google Scholar] [CrossRef]
- Hansen, D.V.; Loktev, A.V.; Ban, K.H.; Jackson, P.K. Plk1 Regulates Activation of the Anaphase Promoting Complex by Phosphorylating and Triggering SCFbetaTrCP-Dependent Destruction of the APC Inhibitor Emi1. Mol. Biol. Cell 2004, 15, 5623–5634. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.; Rauh, N.R.; Nigg, E.A.; Mayer, T.U. Cytostatic Factor: An Activity That Puts the Cell Cycle on Hold. J. Cell Sci. 2006, 119, 1213–1218. [Google Scholar] [CrossRef] [Green Version]
- Tung, J.J.; Hansen, D.V.; Ban, K.H.; Loktev, A.V.; Summers, M.K.; Adler, J.R.; Jackson, P.K. A Role for the Anaphase-Promoting Complex Inhibitor Emi2/XErp1, a Homolog of Early Mitotic Inhibitor 1, in Cytostatic Factor Arrest of Xenopus Eggs. Proc. Natl. Acad. Sci. USA 2005, 102, 4318–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sako, K.; Suzuki, K.; Isoda, M.; Yoshikai, S.; Senoo, C.; Nakajo, N.; Ohe, M.; Sagata, N. Emi2 Mediates Meiotic MII Arrest by Competitively Inhibiting the Binding of Ube2S to the APC/C. Nat. Commun. 2014, 5, 3667. [Google Scholar] [CrossRef] [Green Version]
- Setoyama, D.; Yamashita, M.; Sagata, N. Mechanism of Degradation of CPEB during Xenopus Oocyte Maturation. Proc. Natl. Acad. Sci. USA 2007, 104, 18001–18006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.-W.; Zhu, Y.-Z.; Chen, H.; Wu, Y.-W.; Pi, S.-B.; Chen, L.; Shen, L.; Fan, H.-Y. PABPN1L Mediates Cytoplasmic MRNA Decay as a Placeholder during the Maternal-to-Zygotic Transition. EMBO Rep. 2020, 21, e49956. [Google Scholar] [CrossRef] [PubMed]
- Daldello, E.M.; Le, T.; Poulhe, R.; Jessus, C.; Haccard, O.; Dupré, A. Correction: Control of Cdc6 Accumulation by Cdk1 and MAPK Is Essential for Completion of Oocyte Meiotic Divisions in Xenopus. J. Cell Sci. 2018, 131, jcs215293. [Google Scholar] [CrossRef] [Green Version]
- Spike, C.A.; Huelgas-Morales, G.; Tsukamoto, T.; Greenstein, D. Multiple Mechanisms Inactivate the LIN-41 RNA-Binding Protein To Ensure a Robust Oocyte-to-Embryo Transition in Caenorhabditis Elegans. Genetics 2018, 210, 1011–1037. [Google Scholar] [CrossRef] [Green Version]
- Chesnaye, E.D.L.; Kerr, B.; Paredes, A.; Merchant-Larios, H.; Méndez, J.P.; Ojeda, S.R. Fbxw15/Fbxo12J Is an F-Box Protein-Encoding Gene Selectively Expressed in Oocytes of the Mouse Ovary. Biol. Reprod. 2008, 78, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Moshe, Y.; Boulaire, J.; Pagano, M.; Hershko, A. Role of Polo-like Kinase in the Degradation of Early Mitotic Inhibitor 1, a Regulator of the Anaphase Promoting Complex/Cyclosome. Proc. Natl. Acad. Sci. USA 2004, 101, 7937–7942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, N.; Arai, H.; Iwasaki, J.-I.; Shiina, M.; Ogata, K.; Hunter, T.; Osada, H. Cyclin-Dependent Kinase (CDK) Phosphorylation Destabilizes Somatic Wee1 via Multiple Pathways. Proc. Natl. Acad. Sci. USA 2005, 102, 11663–11668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauh, N.R.; Schmidt, A.; Bormann, J.; Nigg, E.A.; Mayer, T.U. Calcium Triggers Exit from Meiosis II by Targeting the APC/C Inhibitor XErp1 for Degradation. Nature 2005, 437, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Mendez, R.; Barnard, D.; Richter, J.D. Differential MRNA Translation and Meiotic Progression Require Cdc2-Mediated CPEB Destruction. EMBO J. 2002, 21, 1833–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welcker, M.; Clurman, B.E. FBW7 Ubiquitin Ligase: A Tumour Suppressor at the Crossroads of Cell Division, Growth and Differentiation. Nat. Rev. Cancer 2008, 8, 83–93. [Google Scholar] [CrossRef]
- Spike, C.A.; Coetzee, D.; Eichten, C.; Wang, X.; Hansen, D.; Greenstein, D. The TRIM-NHL Protein LIN-41 and the OMA RNA-Binding Proteins Antagonistically Control the Prophase-to-Metaphase Transition and Growth of Caenorhabditis Elegans Oocytes. Genetics 2014, 198, 1535–1558. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.R.; Francis, R.; Schedl, T. GLD-1, a Cytoplasmic Protein Essential for Oocyte Differentiation, Shows Stage- and Sex-Specific Expression during Caenorhabditis Elegans Germline Development. Dev. Biol. 1996, 180, 165–183. [Google Scholar] [CrossRef]
- Cheng, X.; Pei, P.; Yu, J.; Zhang, Q.; Li, D.; Xie, X.; Wu, J.; Wang, S.; Zhang, T. F-Box Protein FBXO30 Mediates Retinoic Acid Receptor γ Ubiquitination and Regulates BMP Signaling in Neural Tube Defects. Cell Death Dis. 2019, 10, 551. [Google Scholar] [CrossRef]
- Adhikari, D.; Zheng, W.; Shen, Y.; Gorre, N.; Ning, Y.; Halet, G.; Kaldis, P.; Liu, K. Cdk1, but Not Cdk2, Is the Sole Cdk That Is Essential and Sufficient to Drive Resumption of Meiosis in Mouse Oocytes. Hum. Mol. Genet. 2012, 21, 2476–2484. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Tripurani, S.K.; Wanna, W.; Rexroad, C.E.; Yao, J. Cloning and Characterization of a Novel Oocyte-Specific Gene Encoding an F-Box Protein in Rainbow Trout (Oncorhynchus Mykiss). Reprod. Biol. Endocrinol. 2013, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Shimuta, K.; Nakajo, N.; Uto, K.; Hayano, Y.; Okazaki, K.; Sagata, N. Chk1 Is Activated Transiently and Targets Cdc25A for Degradation at the Xenopus Midblastula Transition. EMBO J. 2002, 21, 3694–3703. [Google Scholar] [CrossRef] [Green Version]
- Collart, C.; Smith, J.C.; Zegerman, P. Chk1 Inhibition of the Replication Factor Drf1 Guarantees Cell-Cycle Elongation at the Xenopus Laevis Mid-Blastula Transition. Dev. Cell 2017, 42, 82–96.e3. [Google Scholar] [CrossRef] [Green Version]
- Kinterova, V.; Kanka, J.; Petruskova, V.; Toralova, T. Inhibition of SCF Complexes during Bovine Oocyte Maturation and Preimplantation Development Leads to Delayed Development of Embryos. Biol. Reprod. 2018, 100, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Toralova, T.; Kinterova, V.; Chmelikova, E.; Kanka, J. The Neglected Part of Early Embryonic Development: Maternal Protein Degradation. Cell. Mol. Life Sci. 2020, 77, 3177–3194. [Google Scholar] [CrossRef]
- Benesova, V.; Kinterova, V.; Kanka, J.; Toralova, T. Potential Involvement of SCF-Complex in Zygotic Genome Activation During Early Bovine Embryo Development. Methods Mol. Biol. 2017, 1605, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kou, Z.; Jing, Z.; Zhang, Y.; Guo, X.; Dong, M.; Wilmut, I.; Gao, S. Proteome of Mouse Oocytes at Different Developmental Stages. Proc. Natl. Acad. Sci. USA 2010, 107, 17639–17644. [Google Scholar] [CrossRef] [Green Version]
- Knowles, B.B.; Evsikov, A.V.; de Vries, W.N.; Peaston, A.E.; Solter, D. Molecular Control of the Oocyte to Embryo Transition. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 1381–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Ferreira, I.; Bento, I.; Pimenta-Marques, A.; Jana, S.C.; Lince-Faria, M.; Duarte, P.; Borrego-Pinto, J.; Gilberto, S.; Amado, T.; Brito, D.; et al. Regulation of Autophosphorylation Controls PLK4 Self-Destruction and Centriole Number. Curr. Biol. 2013, 23, 2245–2254. [Google Scholar] [CrossRef] [Green Version]
- Muzzopappa, M.; Wappner, P. Multiple Roles of the F-Box Protein Slimb in Drosophila Egg Chamber Development. Development 2005, 132, 2561–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peel, N.; Dougherty, M.; Goeres, J.; Liu, Y.; O’Connell, K.F. The C. Elegans F-Box Proteins LIN-23 and SEL-10 Antagonize Centrosome Duplication by Regulating ZYG-1 Levels. J. Cell Sci. 2012, 125, 3535–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.; He, S.; Xing, C.; Lu, K.; Wang, J.; Xing, G.; Meng, A.; Jia, S.; He, F.; Zhang, L. SCFFBXL15 Regulates BMP Signalling by Directing the Degradation of HECT-Type Ubiquitin Ligase Smurf1. EMBO J. 2011, 30, 2675–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avilés-Pagán, E.E.; Kang, A.S.W.; Orr-Weaver, T.L. Identification of New Regulators of the Oocyte-to-Embryo Transition in Drosophila. G3 Genes Genomes Genet. 2020, 10, 2989–2998. [Google Scholar] [CrossRef] [PubMed]
- Solc, P.; Saskova, A.; Baran, V.; Kubelka, M.; Schultz, R.M.; Motlik, J. CDC25A Phosphatase Controls Meiosis I Progression in Mouse Oocytes. Dev. Biol. 2008, 317, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Bertke, M.M.; Champion, M.M.; Zhu, G.; Huber, P.W.; Dovichi, N.J. Quantitative Proteomics of Xenopus Laevis Embryos: Expression Kinetics of Nearly 4000 Proteins during Early Development. Sci. Rep. 2014, 4, 4365. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Zhou, C.; Wang, Y.; Liu, W.; Liu, C.; Wang, L.; Liu, Y.; Shang, Y.; Li, M.; Zhou, S.; et al. The E3 Ubiquitin Ligase RNF114 and TAB1 Degradation Are Required for Maternal-to-Zygotic Transition. EMBO Rep. 2017, 18, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higuchi, C.; Yamamoto, M.; Shin, S.-W.; Miyamoto, K.; Matsumoto, K. Perturbation of Maternal PIASy Abundance Disrupts Zygotic Genome Activation and Embryonic Development via SUMOylation Pathway. Biol. Open 2019, 8, bio048652. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCF Ligase | Substrates | Animal | Reference |
|---|---|---|---|
| SCFβTrCP | Emi2/Erp1 | Xenopus laevis | [44] |
| Emi1 | [45] | ||
| CPEB | [46] | ||
| BTG4 | Mouse | [47] | |
| SCFSEL-10/FBXW7/CDC4 | GLD-1,CPB-3 | Caenorhabditis elegans | [37] |
| CDC6 | [48] | ||
| LIN-41 | [49] | ||
| SCFFBXO30 | SLBP | Mouse | [38] |
| SCFFBXO34 | Not identified | Mouse | [39] |
| SCFFBXW15,FBXO12J | Not identified | Mouse | [50] |
| SCF Ligases and Their Substrates in Embryogenesis | |||
|---|---|---|---|
| SCF Ligase | Substrates | Animal | Reference |
| SCFβ-TrCP/Slimb/LIN-23 | DRF1 | Xenopus laevis | [62] |
| CDC25A | [61] | ||
| PLK4 | Drosophila melanogaster | [68] | |
| Medea | [69] | ||
| ZYG-1 | Caenorhabditis elegans | [70] | |
| SCFSEL-10/FBXW7/CDC4 | ZYG-1 | Caenorhabditis elegans | [70] |
| SCFFBXO30 | RARγ, BMP | Human | [58] |
| SCFFBXL15 | Smurf1, BMP | Zebrafish | [71] |
| SCFCG5003 | Not identified | Drosophila melanogaster | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kinterová, V.; Kaňka, J.; Bartková, A.; Toralová, T. SCF Ligases and Their Functions in Oogenesis and Embryogenesis—Summary of the Most Important Findings throughout the Animal Kingdom. Cells 2022, 11, 234. https://doi.org/10.3390/cells11020234
Kinterová V, Kaňka J, Bartková A, Toralová T. SCF Ligases and Their Functions in Oogenesis and Embryogenesis—Summary of the Most Important Findings throughout the Animal Kingdom. Cells. 2022; 11(2):234. https://doi.org/10.3390/cells11020234
Chicago/Turabian StyleKinterová, Veronika, Jiří Kaňka, Alexandra Bartková, and Tereza Toralová. 2022. "SCF Ligases and Their Functions in Oogenesis and Embryogenesis—Summary of the Most Important Findings throughout the Animal Kingdom" Cells 11, no. 2: 234. https://doi.org/10.3390/cells11020234