
Generation and Characterization of a Novel Angelman Syndrome Mouse Model with a Full Deletion of the Ube3a Gene
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of the Ube3agenedel Model
2.2. Mouse Husbandry, Breeding, and Experimental Cohorts
2.3. Western Blotting
2.4. Rotarod
2.5. Gait Analysis
2.6. Tail Suspension Test
2.7. Open Field
2.8. Elevated plus Maze
2.9. Novel Object Recognition
2.10. Nest Building
2.11. Barnes Maze
2.12. Testing in Home-Cage Environment IntelliCage
2.13. Statistical Analyses
3. Results
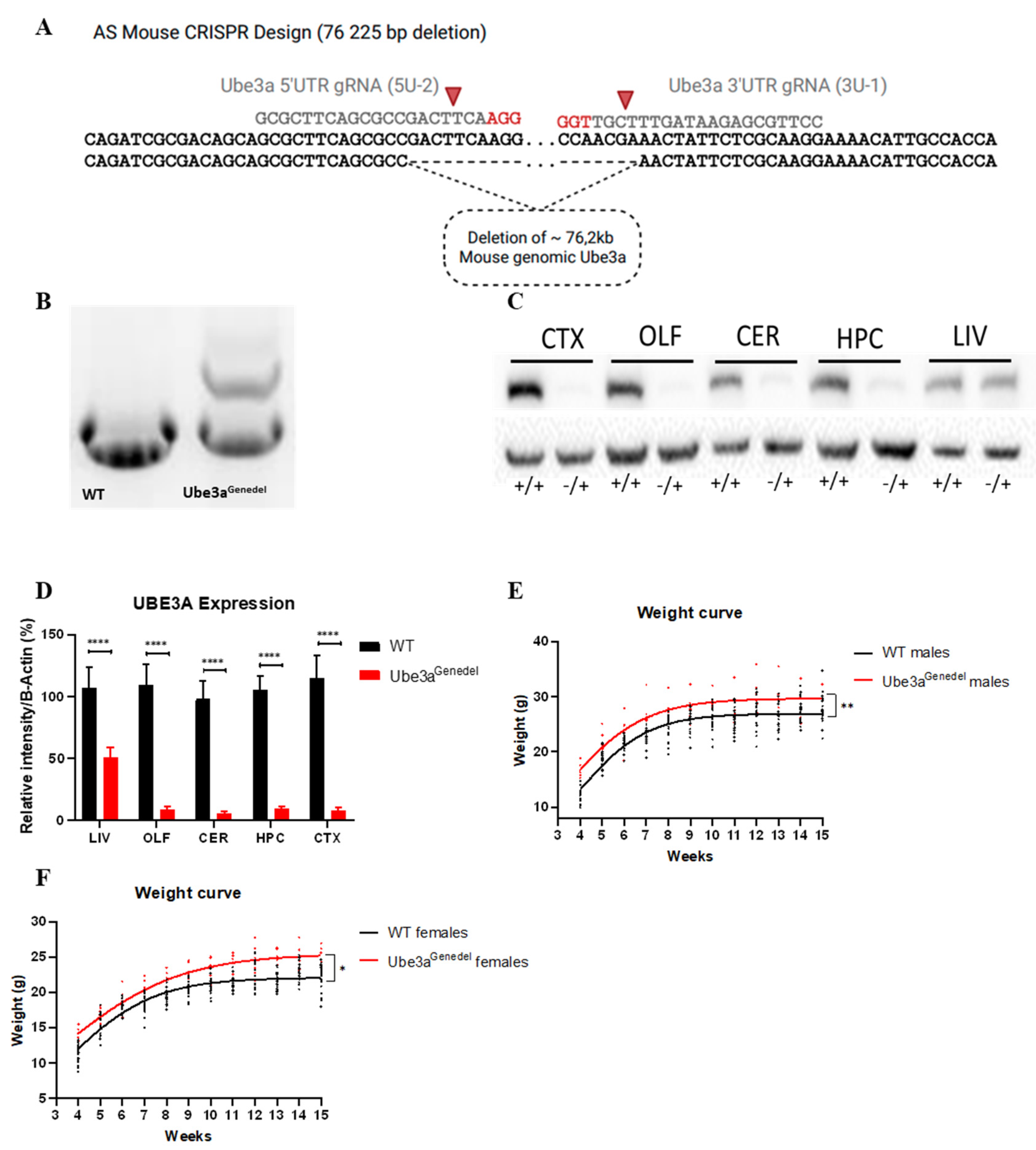
3.1. The New Ube3agenedel Model Harbors a Large Deletion Spanning the Entire Ube3a Gene
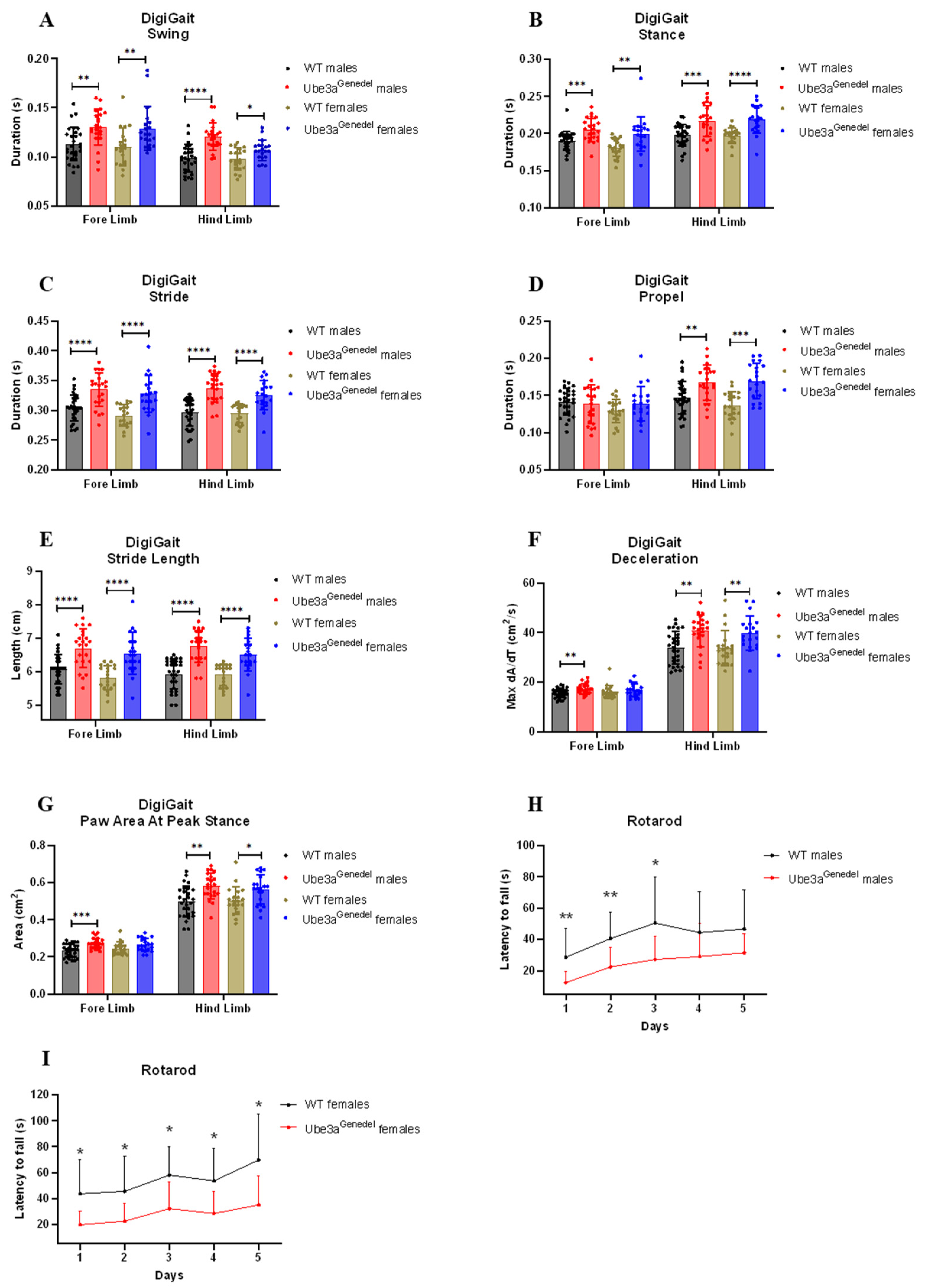
3.2. Gait Impairment and Impaired Motor Skills Are Recapitulated in The Novel Ube3agenedel Mouse
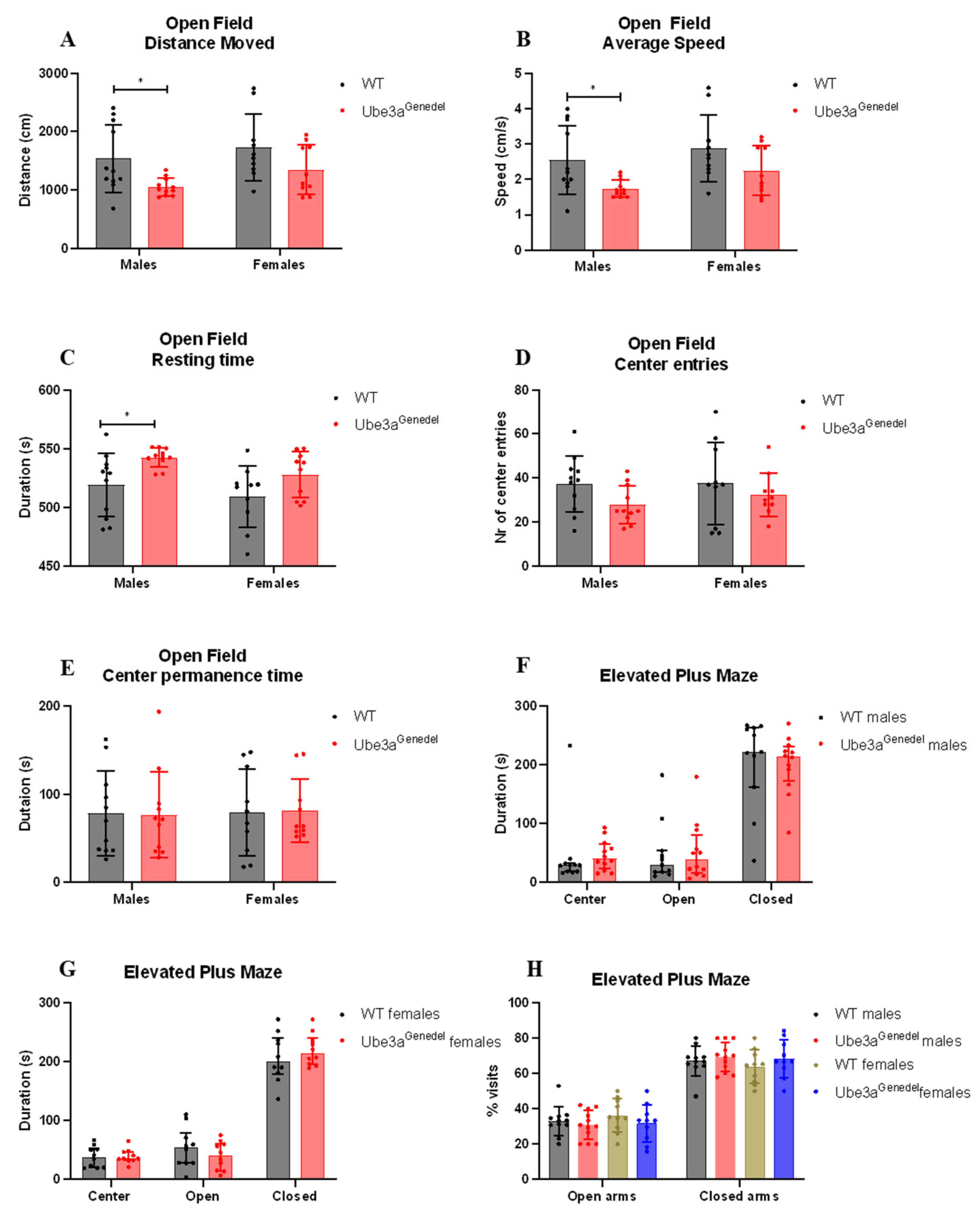
3.3. The Ube3agenedel Model Exhibits Robust Behavioral Impairment
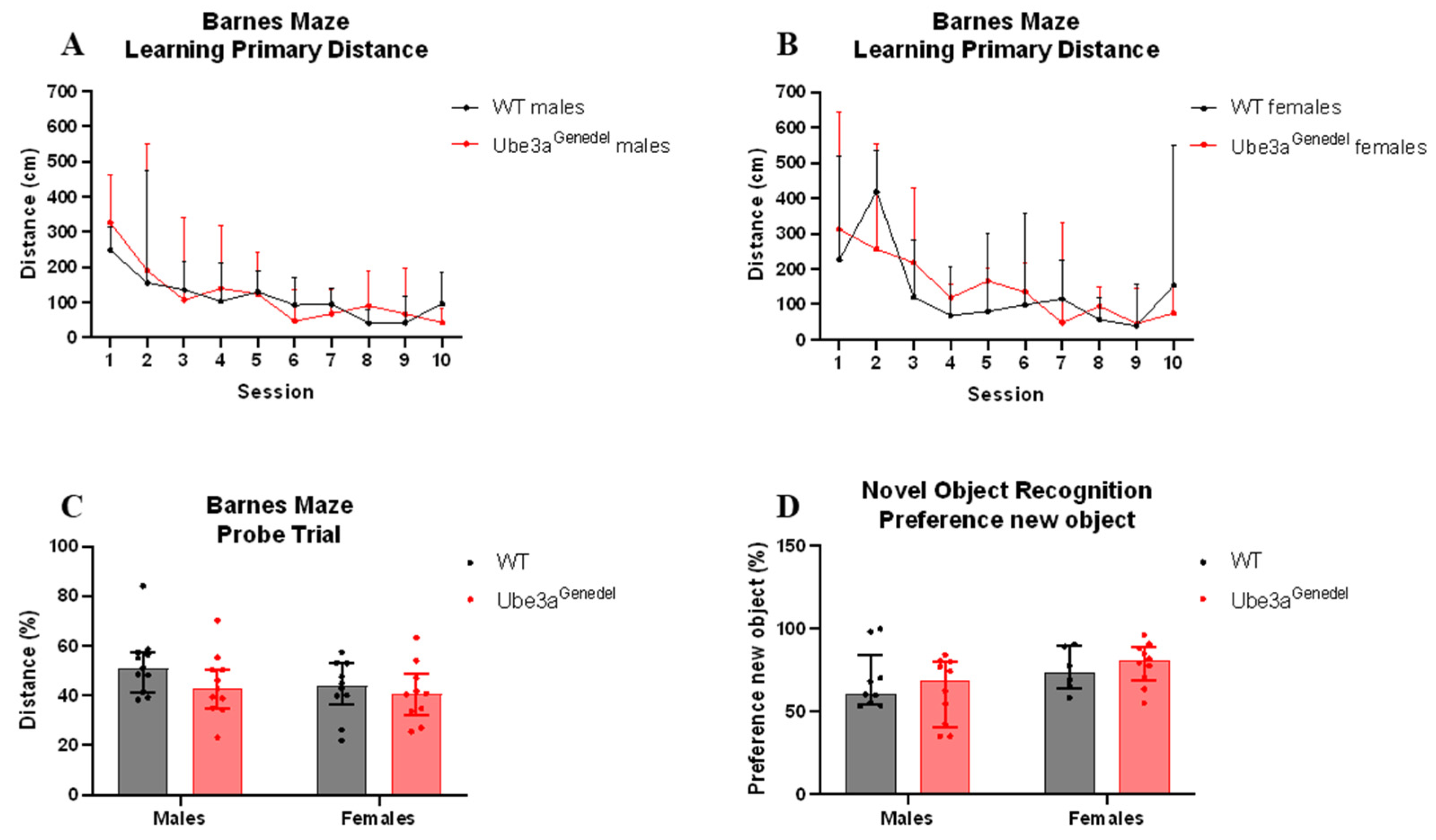
3.4. Memory and Learning Were Not Impaired in Ube3agenedel Mice in Barnes Maze and Novel Object Recognition Tests
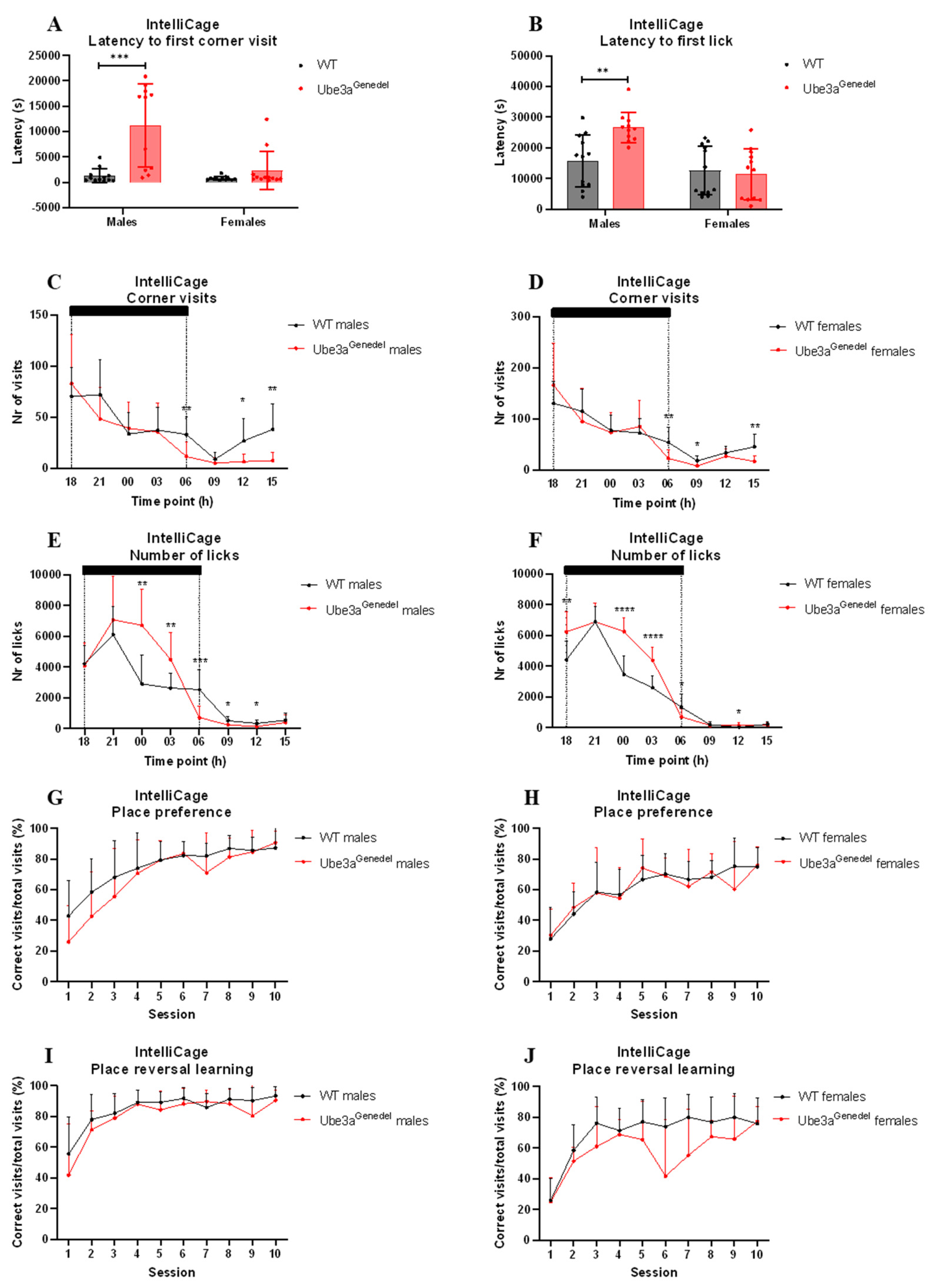
3.5. Home-Cage Circadian Activity, Response to Novelty, and Performance in Place Reversal Tasks Differ in Ube3agenedel Animals
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- layton-Smith, J.; Laan, L. Angelman syndrome: A review of the clinical and genetic aspects. J. Med. Genet. 2003, 40, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Jolleff, N.; Ryan, M.M. Communication development in Angelman’s syndrome. Arch. Dis. Child. 1993, 69, 148–150. [Google Scholar] [CrossRef]
- Viani, F.; Romeo, A.; Viri, M.; Mastrangelo, M.; Lalatta, F.; Selicorni, A.; Gobbi, G.; Lanzi, G.; Bettio, D.; Briscioli, V.; et al. Seizure and EEG Patterns in Angelman’s Syndrome. J. Child. Neurol. 1995, 10, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Person, R.E.; Beaudet, A.L. Ube3a-ATS is an atypical RNA polymerase II transcript that represses the paternal expression of Ube3a. Hum. Mol. Genet. 2012, 21, 3001–3012. [Google Scholar] [CrossRef] [PubMed]
- Sandanam, T.; Beange, H.; Robson, L.; Woolnough, H.; Buchholz, T.; Smith, A. Manifestations in institutionalised adults with Angelman syndrome due to deletion. Am. J. Med. Genet. 1997, 70, 415–420. [Google Scholar] [CrossRef]
- Malcolm, S.; Clayton-Smith, J.; Nichols, M.; Pembrey, M.; Armour, J.; Jeffreys, A.; Robb, S.; Webb, T. Uniparental paternal disomy in Angelman’s syndrome. Lancet 1991, 337, 694–697. [Google Scholar] [CrossRef]
- Buiting, K.; Gross, S.; Lich, C.; Gillessen-Kaesbach, G.; el-Maarri, O.; Horsthemke, B. Epimutations in Prader-Willi and Angelman syndromes: A molecular study of 136 patients with an im-printing defect. Am. J. Hum. Genet. 2003, 72, 571–577. [Google Scholar] [CrossRef]
- Matsuura, T.; Sutcliffe, J.S.; Fang, P.; Galjaard, R.-J.; Jiang, Y.-H.; Benton, C.S.; Rommens, J.M.; Beaudet, A.L. De novo truncating mutations in E6-AP ubiquitin-protein ligase gene (UBE3A) in Angelman syndrome. Nat. Genet. 1997, 15, 74–77. [Google Scholar] [CrossRef]
- Kishino, T.; Lalande, M.; Wagstaff, J. UBE3A/E6-AP mutations cause Angelman syndrome. Nat. Genet. 1997, 15, 70–73. [Google Scholar] [CrossRef]
- Syding, L.A.; Nickl, P.; Kasparek, P.; Sedlacek, R. CRISPR/Cas9 Epigenome Editing Potential for Rare Imprinting Diseases: A Review. Cells 2020, 9, 993. [Google Scholar] [CrossRef]
- Williams, C.A.; Beaudet, A.L.; Clayton-Smith, J.; Knoll, J.H.; Kyllerman, M.; Laan, L.A.; Magenis, R.E.; Moncla, A.; Schinzel, A.A.; Summers, J.A.; et al. Angelman syndrome 2005: Updated consensus for diagnostic criteria. Am. J. Med. Genet. Part A 2006, 140A, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Koyavski, L.; Panov, J.; Simchi, L.; Rayi, P.R.; Sharvit, L.; Feuermann, Y.; Kaphzan, H. Sex-Dependent Sensory Phenotypes and Related Transcriptomic Expression Profiles Are Differentially Affected by Angelman Syndrome. Mol. Neurobiol. 2019, 56, 5998–6016. [Google Scholar] [CrossRef] [PubMed]
- van Woerden, G.M.; Harris, K.D.; Hojjati, M.R.; Gustin, R.M.; Qiu, S.; de Avila Freire, R.; Jiang, Y.H.; Elgersma, Y.; Weeber, E.J. Rescue of neurological deficits in a mouse model for Angelman syndrome by reduction of alphaCaMKII inhibitory phosphorylation. Nat. Neurosci. 2007, 10, 280–282. [Google Scholar] [CrossRef]
- Sidorov, M.S.; Judson, M.C.; Kim, H.; Rougie, M.; Ferrer, A.I.; Nikolova, V.D.; Riddick, N.V.; Moy, S.S.; Philpot, B.D. Enhanced Operant Extinction and Prefrontal Excitability in a Mouse Model of Angelman Syndrome. J. Neurosci. 2018, 38, 2671–2682. [Google Scholar] [CrossRef] [PubMed]
- Sonzogni, M.; Wallaard, I.; Santos, S.S.; Kingma, J.; Du Mee, D.; Van Woerden, G.M.; Elgersma, Y. A behavioral test battery for mouse models of Angelman syndrome: A powerful tool for testing drugs and novel Ube3a mutants. Mol. Autism 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Albrecht, U.; Sutcliffe, J.S.; Cattanach, B.M.; Beechey, C.V.; Armstrong, D.; Eichele, G.; Beaudet, A.L. Imprinted expression of the murine Angelman syndrome gene, Ube3a, in hippocampal and Purkinje neurons. Nat. Genet. 1997, 17, 75–78. [Google Scholar] [CrossRef]
- Jiang, Y.-H.; Pan, Y.; Zhu, L.; Landa, L.; Yoo, J.; Spencer, C.; Lorenzo, I.; Brilliant, M.; Noebels, J.; Beaudet, A.L. Altered Ultrasonic Vocalization and Impaired Learning and Memory in Angelman Syndrome Mouse Model with a Large Maternal Deletion from Ube3a to Gabrb3. PLoS ONE 2010, 5, e12278. [Google Scholar] [CrossRef]
- Avagliano Trezza, R.; Sonzogni, M.; Bossuyt, S.N.V.; Zampeta, F.I.; Punt, A.M.; van den Berg, M.; Rotaru, D.C.; Koene, L.M.C.; Munshi, S.T.; Stedehouder, J. Loss of nuclear UBE3A causes electrophysiological and behavioral deficits in mice and is asso-ciated with Angelman syndrome. Nat. Neurosci. 2019, 22, 1235–1247. [Google Scholar] [CrossRef]
- Gabriel, J.M.; Gabriel, J.M.; Merchant, M.; Ohta, T.; Ji, Y.; Caldwell, R.G.; Ramsey, M.J.; Tucker, J.D.; Longnecker, R.; Nicholls, R.D. A transgene insertion creating a heritable chromosome deletion mouse model of Prader-Willi and angelman syn-dromes. Proc. Natl. Acad. Sci. USA 1999, 96, 9258–9263. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Armstrong, D.; Albrecht, U.; Atkins, C.M.; Noebels, J.L.; Eichele, G.; Sweatt, J.D.; Beaudet, A.L. Mutation of the Angelman ubiquitin ligase in mice causes increased cytoplasmic p53 and deficits of con-textual learning and long-term potentiation. Neuron 1998, 21, 799–811. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Van Woerden, G.M.; Elgersma, Y.; Borst, J.G.G. Enhanced Transmission at the Calyx of Held Synapse in a Mouse Model for Angelman Syndrome. Front. Cell. Neurosci. 2018, 11, 418. [Google Scholar] [CrossRef]
- Ube3a Mouse Gene Details | Ubiquitin Protein Ligase E3A | International Mouse Phenotyping Consortium. Available online: https://www.mousephenotype.org/ (accessed on 25 March 2022).
- Berrios, J.; Stamatakis, A.M.; Kantak, P.A.; McElligott, Z.A.; Judson, M.C.; Aita, M.; Rougie, M.; Stuber, G.D.; Philpot, B.D. Loss of UBE3A from TH-expressing neurons suppresses GABA co-release and enhances VTA-NAc optical self-stimulation. Nat. Commun. 2016, 7, 10702. [Google Scholar] [CrossRef] [PubMed]
- Silva-Santos, S.; Van Woerden, G.M.; Bruinsma, C.F.; Mientjes, E.; Jolfaei, M.A.; Distel, B.; Kushner, S.; Elgersma, Y. Ube3a reinstatement identifies distinct developmental windows in a murine Angelman syndrome model. J. Clin. Investig. 2015, 125, 2069–2076. [Google Scholar] [CrossRef] [PubMed]
- Skarnes, W.C.; Rosen, B.; West, A.P.; Koutsourakis, M.; Bushell, W.; Iyer, V.; Mujica, A.O.; Thomas, M.; Harrow, J.; Cox, T.; et al. A conditional knockout resource for the genome-wide study of mouse gene function. Nature 2011, 474, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.P.; Zhou, Y.-D.; Zhang, G.; Jin, Z.; Stoppel, D.C.; Anderson, M.P. Increased Gene Dosage of Ube3a Results in Autism Traits and Decreased Glutamate Synaptic Transmission in Mice. Sci. Transl. Med. 2011, 3, 103ra97. [Google Scholar] [CrossRef]
- Krishnan, V.; Stoppel, D.C.; Nong, Y.; Johnson, M.A.; Nadler, M.J.S.; Ozkaynak, E.; Teng, B.L.; Nagakura, I.; Mohammad, F.; Silva, M.A.; et al. Autism gene Ube3a and seizures impair sociability by repressing VTA Cbln1. Nature 2017, 543, 507–512. [Google Scholar] [CrossRef]
- Matsumoto, A.; Kumagai, T.; Miura, K.; Miyazaki, S.; Hayakawa, C.; Yamanaka, T. Epilepsy in Angelman Syndrome Associated with Chromosome 15q Deletion. Epilepsia 1992, 33, 1083–1090. [Google Scholar] [CrossRef]
- Dindot, S.V.; Antalffy, B.A.; Bhattacharjee, M.B.; Beaudet, A.L. The Angelman syndrome ubiquitin ligase localizes to the synapse and nucleus, and maternal deficiency results in abnormal dendritic spine morphology. Hum. Mol. Genet. 2007, 17, 111–118. [Google Scholar] [CrossRef]
- Copping, N.A.; Christian, S.G.B.; Ritter, D.G.; Islam, M.S.; Buscher, N.; Zolkowska, D.; Pride, M.C.; Berg, E.L.; LaSalle, J.M.; Ellegood, J.; et al. Neuronal overexpression of Ube3a isoform 2 causes behavioral impairments and neuroanatomical pathology relevant to 15q11.2-q13.3 duplication syndrome. Hum. Mol. Genet. 2017, 26, 3995–4010. [Google Scholar] [CrossRef]
- The Jackson Laboratory, I.o.f.T.J.L., Bar Harbor, ME. Unpublished. 2005–2017. Available online: https://www.jax.org/ (accessed on 25 March 2022).
- Miura, K.; Kishino, T.; Li, E.; Webber, H.; Dikkes, P.; Holmes, G.L.; Wagstaff, J. Neurobehavioral and Electroencephalographic Abnormalities in Ube3aMaternal-Deficient Mice. Neurobiol. Dis. 2002, 9, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Tsai, T.F.; Jiang, Y.H.; Bressler, J.; Armstrong, D.; Beaudet, A.L. Paternal deletion from Snrpn to Ube3a in the mouse causes hypotonia, growth retardation and partial le-thality and provides evidence for a gene contributing to Prader-Willi syndrome. Hum. Mol. Genet. 1999, 8, 1357–1364. [Google Scholar] [CrossRef]
- Russell, L.B.; Montgomery, C.S.; Cacheiro, N.L.; Johnson, D.K. Complementation analyses for 45 mutations encompassing the pink-eyed dilution (p) locus of the mouse. Genetics 1995, 141, 1547–1562. [Google Scholar] [CrossRef] [PubMed]
- Rotaru, D.C.; Mientjes, E.J.; Elgersma, Y. Angelman Syndrome: From Mouse Models to Therapy. Neuroscience 2020, 445, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Born, H.A.; Dao, A.T.; Levine, A.T.; Lee, W.L.; Mehta, N.M.; Mehra, S.; Weeber, E.J.; Anderson, A.E. Strain-dependence of the Angelman Syndrome phenotypes in Ube3a maternal deficiency mice. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Shiotsuki, H.; Yoshimi, K.; Shimo, Y.; Funayama, M.; Takamatsu, Y.; Ikeda, K.; Takahashi, R.; Kitazawa, S.; Hattori, N. A rotarod test for evaluation of motor skill learning. J. Neurosci. Methods 2010, 189, 180–185. [Google Scholar] [CrossRef]
- Hampton, T.G.; Kale, A.; Amende, I.; Tang, W.; McCue, S.; Bhagavan, H.N.; VanDongen, C.G. Gait Disturbances in Dystrophic Hamsters. J. Biomed. Biotechnol. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Bertin, A.; Jalfre, M. Behavioral despair in mice: A primary screening test for antidepressants. Arch. Int. Pharmacodyn. Ther. 1977, 229, 327–336. [Google Scholar] [PubMed]
- Kulesskaya, N.; Voikar, V. Assessment of mouse anxiety-like behavior in the light–dark box and open-field arena: Role of equipment and procedure. Physiol. Behav. 2014, 133, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Lister, R.G. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology 1987, 92, 180–185. [Google Scholar] [CrossRef]
- Chiba, A.A.; Kesner, R.P.; Reynolds, A.M. Memory for spatial location as a function of temporal lag in rats: Role of hip-pocampus and medial prefrontal cortex. Behav. Neural Biol. 1994, 61, 123–131. [Google Scholar] [CrossRef]
- Youn, J.; Ellenbroek, B.A.; van Eck, I.; Roubos, S.; Verhage, M.; Stiedl, O. Finding the right motivation: Genotype-dependent differences in effective reinforcements for spatial learning. Behav. Brain Res. 2012, 226, 397–403. [Google Scholar] [CrossRef]
- O’Leary, T.P.; Brown, R.E. The effects of apparatus design and test procedure on learning and memory performance of C57BL/6J mice on the Barnes maze. J. Neurosci. Methods 2012, 203, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.; Endo, T.; Endo, N.; Kakeyama, M.; Tohyama, C. Early deprivation induces competitive subordinance in C57BL/6 male mice. Physiol. Behav. 2014, 137, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Margolis, S.S.; Sell, G.L.; Zbinden, M.A.; Bird, L.M. Angelman Syndrome. Neurotherapeutics 2015, 12, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Jirkof, P. Burrowing and nest building behavior as indicators of well-being in mice. J. Neurosci. Methods 2014, 234, 139–146. [Google Scholar] [CrossRef]
- Yamasaki, K.; Joh, K.; Ohta, T.; Masuzaki, H.; Ishimaru, T.; Mukai, T.; Niikawa, N.; Ogawa, M.; Wagstaff, J.; Kishino, T. Neurons but not glial cells show reciprocal imprinting of sense and antisense transcripts of Ube3a. Hum. Mol. Genet. 2003, 12, 837–847. [Google Scholar] [CrossRef]
- Duca, D.G.; Craiu, D.; Boer, M.; Chirieac, S.M.; Arghir, A.; Tutulan-Cunita, A.; Barca, D.; Iliescu, C.; Lungeanu, A.; Magureanu, S.; et al. Diagnostic approach of angelman syndrome. Maedica (Bucur) 2013, 8, 321–327. [Google Scholar]
- Heck, D.; Zhao, Y.; Roy, S.; LeDoux, M.S.; Reiter, L.T. Analysis of cerebellar function in Ube3a-deficient mice reveals novel genotype-specific behaviors. Hum. Mol. Genet. 2008, 17, 2181–2189. [Google Scholar] [CrossRef]
- Berg, E.L.; Petkova, S.P.; Born, H.A.; Adhikari, A.; Anderson, A.E.; Silverman, J.L. Insulin-like growth factor-2 does not improve behavioral deficits in mouse and rat models of Angelman Syndrome. Mol. Autism 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Dodge, A.; Peters, M.M.; Greene, H.E.; Dietrick, C.; Botelho, R.; Chung, D.; Willman, J.; Nenninger, A.W.; Ciarlone, S.; Kamath, S.G.; et al. Generation of a Novel Rat Model of Angelman Syndrome with a Complete Ube3a Gene Deletion. Autism Res. 2020, 13, 397–409. [Google Scholar] [CrossRef]
- Rostosky, C.M.; Milošević, I. Gait Analysis of Age-dependent Motor Impairments in Mice with Neurodegeneration. J. Vis. Exp. 2018, 136, e57752. [Google Scholar] [CrossRef]
- Rinalduzzi, S.; Trompetto, C.; Marinelli, L.; Alibardi, A.; Missori, P.; Fattapposta, F.; Pierelli, F.; Currà, A. Balance dysfunction in Parkinson’s disease. Biomed. Res. Int. 2015, 2015, 434683. [Google Scholar] [CrossRef] [PubMed]
- Dan, B.; Pelc, K.; Cheron, G. Behavior and neuropsychiatric manifestations in Angelman syndrome. Neuropsychiatr. Dis. Treat. 2008, 4, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-S.; Burns, A.J.; Nonneman, R.J.; Baker, L.K.; Riddick, N.V.; Nikolova, V.D.; Riday, T.T.; Yashiro, K.; Philpot, B.D.; Moy, S.S. Behavioral deficits in an Angelman syndrome model: Effects of genetic background and age. Behav. Brain Res. 2013, 243, 79–90. [Google Scholar] [CrossRef]
- Maranga, C.; Fernandes, T.G.; Bekman, E.; Da Rocha, S.T. Angelman syndrome: A journey through the brain. FEBS J. 2020, 287, 2154–2175. [Google Scholar] [CrossRef] [PubMed]
- Moy, S.S.; Nadler, J.J.; Young, N.B.; Nonneman, R.J.; Segall, S.K.; Andrade, G.M.; Crawley, J.N.; Magnuson, T.R. Social approach and repetitive behavior in eleven inbred mouse strains. Behav. Brain Res. 2008, 191, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.L.; Yang, M.; Lord, C.; Crawley, J.N. Behavioural phenotyping assays for mouse models of autism. Nat. Rev. Neurosci. 2010, 11, 490–502. [Google Scholar] [CrossRef]
- Guariglia, S.R.; Chadman, K.K. Water T-maze: A useful assay for determination of repetitive behaviors in mice. J. Neurosci. Methods 2013, 220, 24–29. [Google Scholar] [CrossRef]
- Heinz, D.E.; Schöttle, V.A.; Nemcova, P.; Binder, F.P.; Ebert, T.; Domschke, K.; Wotjak, C.T. Exploratory drive, fear, and anxiety are dissociable and independent components in foraging mice. Transl. Psychiatry 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Sarkar, P.A.; Shigli, A.; Patidar, C. Happy Puppet syndrome. BMJ Case Rep. 2011, 2011, bcr0920114747. [Google Scholar] [CrossRef]
- Ter Horst, J.P.; De Kloet, E.R.; Schächinger, H.; Oitzl, M.S. Relevance of Stress and Female Sex Hormones for Emotion and Cognition. Cell. Mol. Neurobiol. 2011, 32, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, A.A.; Bevilaqua, M.C.D.N.; da Fonseca, A.M.P.; Nardi, A.E.; Thuret, S.; Dias, G.P. Gender Differences in the Neurobiology of Anxiety: Focus on Adult Hippocampal Neurogenesis. Neural Plast. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Strain | Genotype | Phenotype (Ube3a −/+) | Ref. |
|---|---|---|---|---|
| Deletions and substitutions | Ube3atm1Alb | Deletion of 100 N-terminal amino acids in the encoded protein and a frameshift inactivating all putative protein isoforms | Deficits in context-dependent learning (FC), impaired LTP (EP), increased abundance of p53 in PC (IHC), seizures, and motor disfunction (RRT, RCT) | [20] |
| Ube3atm2Yelg | Nucleotide substitutions in exon 3 result in a stop codon for glutamic acid at position 113 (E113X). | MNTB neurons decreased failure rate (IJWR), faster recovery after AP in neurons, elevated AP amplitude (IJWR), AIS length increased (IHC), reduced STD and fast recovery from STD (IJWR) | [21] | |
| Ube3aem2Yelg | The ATG codon (exon 3) encoding the start codon methionine of UBE3A isoform 2 was mutated into a TGA, resulting in expression of isoform 3 only. | Cytosolic Ube3a isoform, not critical for development of severe AS symptoms | [18] | |
| Ube3aem1Yelg | An ATG codon (Ube3a exon 4/5) encoding the initiating methionine of UBE3A isoform 3 was mutated into an alanine (GCG). Therefore, only isoform 2 is expressed in these mice. | Nuclear Ube3a isoform, crucial for development. Deficiency leads to synaptic changes and impacts excitation and inhibition balance (VCR). Neurobehavioral phenotype confirmed by RRT NB, MB, FST, OF. | [18] | |
| Ube3aem1(IMPC)Hmgu (C57BL/6NCrl) | Intra-exon deletion (exon 3) | Assessed by IMPC pipeline, decreased locomotor activity (OF), decreased food intake (IC), decreased respiratory quotient (IC), increased hematocrit (HEM) | [22] | |
| Floxed alleles | Ube3atm1.1Bdph | A floxed allele, exon 5 flanked by loxP sites. | Enhanced reward-seeking behavior (OS) due to lack of Ube3a in TH neurons (EP, VCR, IHC) | [23] |
| Ube3atm1Yelg | A stop cassette with loxP sites inserted in intron CRE-mediated recombination reinstates gene expression. | Non-CRE recombined mouse recapitulates murine AS phenotype. Recombination leads to partial rescue of the phenotype on the protein (WB), neuronal (EP), and motor behavior levels (OF, MB, RRT, NB, FST) | [24] | |
| Ube3atm1a(KOMP)Wtsi | The critical exon(s) is/are flanked by loxP sites. FLP recombination generates a conditional allele. Subsequent CRE expression results in a knockout mouse. If CRE expression occurs without FLP expression, a reporter knockout mouse is created. | N/A | [25] | |
| Inducible isoforms | Tg(tetO-Ube3a*2)884Svd | The transgene under control of a modified Tet response element (TRE or tetO), transgene of mouse ubiquitin protein ligase E3A (Ube3a) cDNA sequence encoding transcript variant 2 (NM_011668.2) with FLAG tag, and an SV40 polyA signal | Anxiety-like behavior (EPMT, LD) autism (TCSIT), contextual learning deficit (FC), lower seizure threshold (EEG), reduced brain volume (MRI) | [30] unpublisde |
| Tg(tetO-Ube3a*1)1Svd | The transgene under control of a modified Tet response element (TRE or tetO), transgene of mouse ubiquitin protein ligase E3A (Ube3a) cDNA sequence encoding transcript variant 1 (NM_173010.3) with FLAG tag, and an SV40 polyA signal | N/A | [31] unpublished | |
| Modified Ube3a | Tg(Ube3a)1Mpan | Overexpression of Ube3a gene with three FLAG tags | Overexpression leads to development of autistic symptoms, impaired social behavior (TCSIT, RRT, OF, EMPT), decreased communication and repetitive behavior, impaired glutamatergic synaptic transmission, and glutamate release (EP). | [26] |
| Tg(Ube3a)5Mpan | Extra copy of Ube3a transgene in the genome | Increased seizures, impaired social behavior (TCSIT, VT) | [27] | |
| Ube3atm1Jwf | A part of exon 15 and all of exon 16 are fused to IRES-lacZ-neo cassette, resulting in functional impairment of the C-terminal region responsible for ubiquitin protein ligase activity. | Allows Ube3a expression tracing based on LacZ staining, motor disfunction (RRT, BCT, FST), abnormal EEG), increased abundance of p53 in PC (IHC). | [28,32] | |
| Ube3atm2.1Alb/Ube3atm2Alb | Fusion of yellow fluorescent protein (YFP) to exon expression of YFP is through inheritance of the maternal allele and recapitulates endogenous expression. | Phenotype not analyzed; the strain is mainly used to track Ube3a expression. | [29] | |
| Large deletions | Del(7Gabra3-Ube3a)1Yhj | The deletion extending from Gabra3 to Ube3a gene including Atp10a. | Homozygotes exhibit cleft palate and perinatal lethality. In AS mice, impaired behavior (LD, HP, USV, MWM, PPI), motor function (RRT, OF), and seizures | [17] |
| Dp(7Herc2-Mkrn3)1Taku | Insertion of selection cassettes and loxP sites proximal to Herc2 and distal to Mkrn3. CRE-mediated recombination in ES cells led to balanced duplication and deletion of 6.3 Mb region between Herc2 and Mkrn3. | Duplication of the paternal allele results in poor social interaction, behavioral inflexibility, and abnormal ultrasonic vocalizations and correlates with anxiety (TCSIT, MWM, BMT, USV, FC) and altered 5-HT2c receptor signaling (EP). | [15] | |
| Del(7Herc2-Mkrn3)13FRdni | 5Mb deletion of entire AS/PWS locus spanning from Herc2 to Mkrn3 genes via Lmp2a transgene insertion | Neurophysiological and behavioral phenotype, cellular morphology, impaired homeostasis and metabolism, increased mortality, aging, and respiratory problems. Analyzed for PWS only. | [19] | |
| Del(7Ube3a-Snrpn)1Alb | Deletion of genomic DNA from the loxP site within Snrpn to the loxP site within Ube3a | Neurophysiological and behavioral phenotype, impaired growth, increased mortality, aging, and muscle hypotonia. Analyzed for PWS only. | [33] | |
| Oca2p−30PUb | This deletion expands distally from the p locus to Gabrb3, Ube3a, and Ipw. This deletion includes Atp10a. | Used for PWS and Atp10c research, homozygosity lethal, impaired modulation of body fat and/or affecting lipid metabolism (increased total fat) | [34] | |
| C57BL/6NCrl-Ube3a<em1(IMPC)Ccpcz>/Ph | Gene deletion | Impaired motor functions (RRT, GB, TST) and altered behavior (OF, EPMT, NB, BMT InteliCage) | This article |
| Cohort | WT Males | Ube3aGenedel Males | WT Females | Ube3aGenedel Females |
|---|---|---|---|---|
| 1 | 14 | 11 | 10 | 10 |
| 2 | 10 | 11 | 9 | 10 |
| 3 | 12 | 12 | 12 | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Syding, L.A.; Kubik-Zahorodna, A.; Nickl, P.; Novosadova, V.; Kopkanova, J.; Kasparek, P.; Prochazka, J.; Sedlacek, R. Generation and Characterization of a Novel Angelman Syndrome Mouse Model with a Full Deletion of the Ube3a Gene. Cells 2022, 11, 2815. https://doi.org/10.3390/cells11182815
Syding LA, Kubik-Zahorodna A, Nickl P, Novosadova V, Kopkanova J, Kasparek P, Prochazka J, Sedlacek R. Generation and Characterization of a Novel Angelman Syndrome Mouse Model with a Full Deletion of the Ube3a Gene. Cells. 2022; 11(18):2815. https://doi.org/10.3390/cells11182815
Chicago/Turabian StyleSyding, Linn Amanda, Agnieszka Kubik-Zahorodna, Petr Nickl, Vendula Novosadova, Jana Kopkanova, Petr Kasparek, Jan Prochazka, and Radislav Sedlacek. 2022. "Generation and Characterization of a Novel Angelman Syndrome Mouse Model with a Full Deletion of the Ube3a Gene" Cells 11, no. 18: 2815. https://doi.org/10.3390/cells11182815