
Fecal Microbiome Changes and Specific Anti-Bacterial Response in Patients with IBD during Anti-TNF Therapy

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. DNA Extraction, PCR Amplification, Sequencing, and Data Analysis
2.3. Preparation of Bacterial Lysates
2.4. Indirect Enzyme-Linked Immunosorbent Assays (ELISAs)
2.5. Peripheral Blood Mononuclear Cells (PBMCs)
2.6. Statistical Analysis
3. Results
3.1. Cohort Characteristics
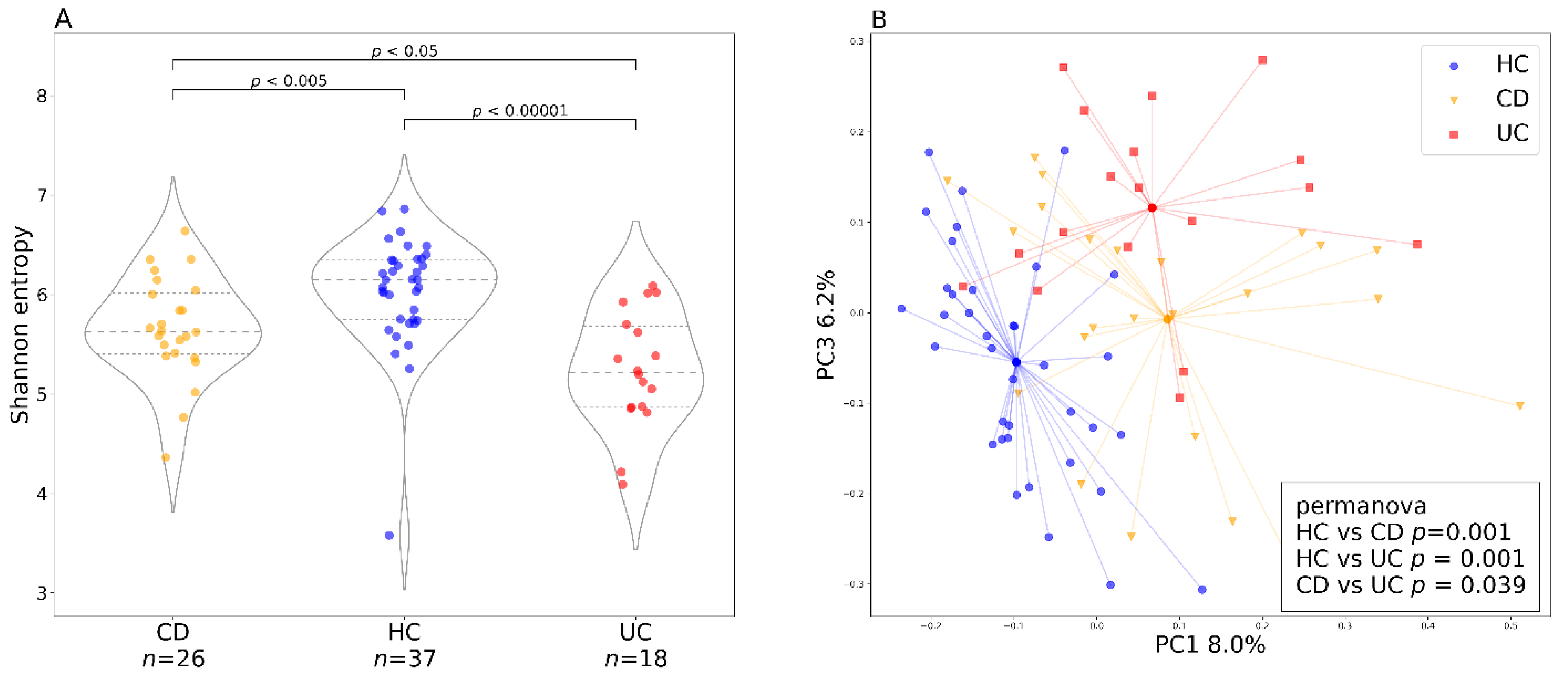
3.2. Individual and Baseline Differences in Gut Bacteriome of Healthy Controls and Patients with IBD
3.3. Factors Influencing Bacterial Diversity
3.4. Changes of the Gut Bacteriome during Anti-TNF Therapy
3.5. Metagenomic Predictions
3.6. Fungal Microbiome
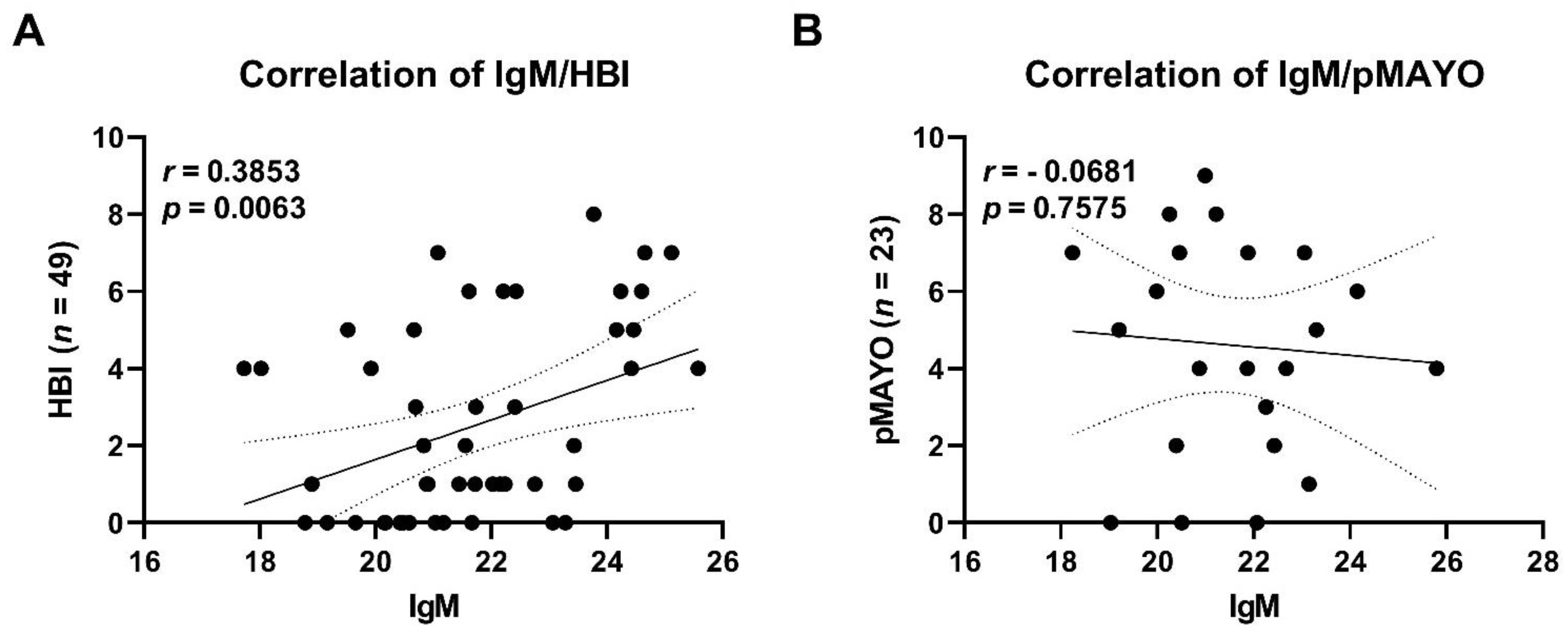
3.7. Serum Antibody Levels against Specific Commensals
3.8. Patients’ PBMCs Produce Increased Levels of IL-17 at the Endpoint of Therapy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Bortlík, M.; Ďuricová, D.; Douda, T.; Konečný, M.; Koželuhová, J.; Novotný, A.; Zbořil, V.; Prokopová, L.; Kohout, P.; Stehlík, J.; et al. Doporučení pro podávání biologické léčby pacientům s idiopatickými střevními záněty: Čtvrté, aktualizované vydání. Gastroenterol. Hepatol. 2019, 73, 11–24. [Google Scholar] [CrossRef]
- Braegger, C.P.; Nicholls, S.; Murch, S.H.; Stephens, S.; MacDonald, T.T. Tumour necrosis factor alpha in stool as a marker of intestinal inflammation. Lancet 1992, 339, 89–91. [Google Scholar] [CrossRef]
- Reimund, J.M.; Wittersheim, C.; Dumont, S.; Muller, C.D.; Kenney, J.S.; Baumann, R.; Poindron, P.; Duclos, B. Increased production of tumour necrosis factor-α, interleukin-1β, and interleukin-6 by morphologically normal intestinal biopsies from patients with Crohn’s disease. Gut 1996, 39, 684–689. [Google Scholar] [CrossRef] [Green Version]
- Marini, M.; Bamias, G.; Rivera-Nieves, J.; Moskaluk, C.A.; Hoang, S.B.; Ross, W.G.; Pizarro, T.T.; Cominelli, F. TNF-a neutralization ameliorates the severity of murine Crohn’s-like ileitis by abrogation of intestinal epithelial cell apoptosis. Proc. Natl. Acad. Sci. USA 2003, 100, 8366–8371. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, H.; Fromm, M.; Bentzel, C.J.; Scholz, P.; Detjen, K.; Mankertz, J.; Bode, H.; Epple, H.J.; Riecken, E.O.; Schulzke, J.D. Tumor necrosis factor-alpha (TNFα) regulates the epithelial barrier in the human intestinal cell line HT-29/B6. J. Cell Sci. 1999, 112, 137–146. [Google Scholar] [CrossRef]
- Friedrich, M.; Pohin, M.; Powrie, F. Cytokine networks in the pathophysiology of inflammatory bowel disease. Immunity 2019, 50, 992–1006. [Google Scholar] [CrossRef] [Green Version]
- Caruso, R.; Lo, B.C.; Núñez, G. Host–microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Aldars-García, L.; Chaparro, M.; Gisbert, J.P. Systematic review: The gut microbiome and its potential clinical application in inflammatory bowel disease. Microorganisms 2021, 9, 977. [Google Scholar] [CrossRef]
- Rajca, S.; Grondin, V.; Louis, E.; Vernier-Massouille, G.; Grimaud, J.C.; Bouhnik, Y.; Laharie, D.; Dupas, J.L.; Pillant, H.; Picon, L.; et al. Alterations in the intestinal microbiome (Dysbiosis) as a predictor of relapse after infliximab withdrawal in Crohn’s disease. Inflamm. Bowel Dis. 2014, 20, 978–986. [Google Scholar] [CrossRef]
- Busquets, D.; Mas-de-Xaxars, T.; López-Siles, M.; Martínez-Medina, M.; Bahí, A.; Sàbat, M.; Louvriex, R.; Miquel-Cusachs, J.O.; Garcia-Gil, J.L.; Aldeguer, X. Anti-tumour necrosis factor treatment with Adalimumab induces changes in the microbiota of Crohn’s disease. J. Crohn’s Colitis 2015, 9, 899–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnusson, M.K.; Strid, H.; Sapnara, M.; Lasson, A.; Bajor, A.; Ung, K.A.; Öhman, L. Anti-TNF therapy response in patients with ulcerative colitis is associated with colonic antimicrobial peptide expression and microbiota composition. J. Crohn’s Colitis 2016, 10, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Juillerat, P.; Øyås, O.; Ramon, C.; Bravo, F.D.; Franc, Y.; Fournier, N.; Michetti, P.; Mueller, C.; Geuking, M.; et al. Microbial network disturbances in relapsing refractory Crohn’s disease. Nat. Med. 2019, 25, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Ribaldone, D.G.; Caviglia, G.P.; Abdulle, A.; Pellicano, R.; Ditto, M.C.; Morino, M.; Fusaro, E.; Saracco, G.M.; Bugianesi, E.; Astegiano, M. Adalimumab therapy improves intestinal dysbiosis in Crohn’s disease. J. Clin. Med. 2019, 8, 1646. [Google Scholar] [CrossRef] [Green Version]
- Aden, K.; Rehman, A.; Waschina, S.; Pan, W.H.; Walker, A.; Lucio, M.; Nunez, A.M.; Bharti, R.; Zimmerman, J.; Bethge, J.; et al. Metabolic functions of gut microbes associate with efficacy of tumor necrosis factor antagonists in patients with inflammatory bowel diseases. Gastroenterology 2019, 157, 1279–1292.e11. [Google Scholar] [CrossRef] [Green Version]
- Kowalska-Duplaga, K.; Kapusta, P.; Gosiewski, T.; Sroka-Oleksiak, A.; Ludwig-Słomczyńska, A.H.; Wołkow, P.P.; Fyderek, K. Changes in the intestinal microbiota are seen following treatment with Infliximab in children with Crohn’s disease. J. Clin. Med. 2020, 9, 687. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Raimondi, S.; Amaretti, A.; Gozzoli, C.; Simone, M.; Righini, L.; Candeliere, F.; Brun, P.; Ardizzoni, A.; Colombari, B.; Paulone, S.; et al. Longitudinal survey of fungi in the human gut: ITS profiling, phenotyping, and colonization. Front. Microbiol. 2019, 10, 1575. [Google Scholar] [CrossRef] [Green Version]
- Ott, S.J.; Kühbacher, T.; Musfeldt, M.; Rosenstiel, P.; Hellmig, S.; Rehman, A.; Drews, O.; Weichert, W.; Timmis, K.N.; Schreiber, S. Fungi and inflammatory bowel diseases: Alterations of composition and diversity. Scand. J. Gastroenterol. 2008, 43, 831–841. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Li, Q.; Wang, C.; Tang, C.; He, Q.; Li, N.; Li, J. Dysbiosis of gut fungal microbiota is associated with mucosal inflammation in Crohn’s disease. J. Clin. Gastroenterol. 2014, 48, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwolinska-Wcislo, M.; Brzozowski, T.; Budak, A.; Kwiecien, S.; Sliwowski, Z.; Drozdowicz, D.; Trojanowska, D.; Rudnicka-Sosin, L.; Mach, T.; Konturek, S.J.; et al. Effect of Candida colonization on human ulcerative colitis and the healing of inflammatory changes of the colon in the experimental model of colitis ulcerosa. J. Physiol. Pharmacol. 2009, 60, 107–118. [Google Scholar] [PubMed]
- Nishino, K.; Nishida, A.; Inoue, R.; Kawada, Y.; Ohno, M.; Sakai, S.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Kawahara, M.; et al. Analysis of endoscopic brush samples identified mucosa-associated dysbiosis in inflammatory bowel disease. J. Gastroenterol. 2018, 53, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A microbial signature for Crohn’s disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Bonovas, S.; Doherty, G.; Kucharzik, T.; Gisbert, J.P.; Raine, T.; Adamina, M.; Armuzzi, A.; Bachmann, O.; Bager, P.; et al. ECCO guidelines on therapeutics in Crohn’s disease: Medical treatment. J. Crohn’s Colitis 2020, 14, 4–22. [Google Scholar] [CrossRef]
- Coufal, S.; Galanova, N.; Bajer, L.; Gajdarova, Z.; Schierova, D.; Jiraskova Zakostelska, Z.; Kostovcikova, K.; Jackova, Z.; Stehlikova, Z.; Drastich, P.; et al. Inflammatory bowel disease types differ in markers of inflammation, gut barrier and in specific anti-bacterial response. Cells 2019, 8, 719. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.F.; Bradshaw, J.M. Index of Crohn’S disease activity. Lancet 1980, 1, 514. [Google Scholar] [CrossRef]
- Schroeder, K.W.; Tremaine, W.J.; Ilstrup, D.M. Coated oral 5-ASA for mildely to moderately active ulcerative colitis. N. Engl. J. Med. 1987, 317, 1625–1629. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Douglas, G.; Maffei, V.; Zaneveld, J.; Yurgel, S.; Brown, J.; Taylor, C.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: An improved and customizable approach for metagenome inference. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Stehlikova, Z.; Tlaskal, V.; Galanova, N.; Roubalova, R.; Kreisinger, J.; Dvorak, J.; Prochazkova, P.; Kostovcikova, K.; Bartova, J.; Libanska, M.; et al. Oral microbiota composition and antimicrobial antibody response in patients with recurrent aphthous stomatitis. Microorganisms 2019, 7, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakostelska, Z.; Kverka, M.; Klimesova, K.; Rossmann, P.; Mrazek, J.; Kopecny, J.; Hornova, M.; Srutkova, D.; Hudcovic, T.; Ridl, J.; et al. Lysate of probiotic Lactobacillus casei DN-114 001 ameliorates colitis by strengthening the gut barrier function and changing the gut microenvironment. PLoS ONE 2011, 6, e27961. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, J.J.; Guruge, J.L.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman, A.L.; Clemente, J.C.; Knight, R.; Heath, A.C.; Leibel, R.L.; et al. The long-term stability of the human gut microbiota. Science 2013, 341, 1237439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, R.; Abu-Ali, G.; Drew, D.; Lloyd-Price, J.; Subramanian, A.; Lochhead, P.; Joshi, A.; Ivey, K.; Khalili, H.; Brown, G.; et al. Stability of the human faecal microbiome in a cohort of adult men. Nat. Microbiol. 2018, 3, 347–355. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Costello, E.K.; Berg-Lyons, D.; Gonzalez, A.; Stombaugh, J.; Knights, D.; Gajer, P.; Ravel, J.; Fierer, N.; et al. Moving pictures of the human microbiome. Genome Biol. 2011, 12, R50. [Google Scholar] [CrossRef] [Green Version]
- Yun, Y.; Kim, H.N.; Kim, S.E.; Heo, S.G.; Chang, Y.; Ryu, S.; Shin, H.; Kim, H.L. Comparative analysis of gut microbiota associated with body mass index in a large Korean cohort. BMC Microbiol. 2017, 17, 151. [Google Scholar] [CrossRef] [Green Version]
- Ventin-Holmberg, R.; Eberl, A.; Saqib, S.; Korpela, K.; Virtanen, S.; Sipponen, T.; Salonen, A.; Saavalainen, P.; Nissilä, E. Bacterial and fungal profiles as markers of infliximab drug response in inflammatory bowel disease. J. Crohn’s Colitis 2021, 15, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2015, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Wirth, U.; Garzetti, D.; Jochum, L.M.; Spriewald, S.; Kühn, F.; Ilmer, M.; Lee, S.M.; Niess, H.; Bazhin, A.V.; Andrassy, J.; et al. Microbiome analysis from paired mucosal and fecal samples of a colorectal cancer biobank. Cancers 2020, 12, 3702. [Google Scholar] [CrossRef] [PubMed]
- Osterman, M.T.; Lichtenstein, G.R. Infliximab vs. Adalimumab for UC: Is there a difference? Clin. Gastroenterol. Hepatol. 2017, 15, 1197–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorlund, K.; Druyts, E.; Mills, E.J.; Fedorak, R.N.; Marshall, J.K. Adalimumab versus infliximab for the treatment of moderate to severe ulcerative colitis in adult patients naïve to anti-TNF therapy: An indirect treatment comparison meta-analysis. J. Crohn’s Colitis 2014, 8, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doecke, J.D.; Hartnell, F.; Bampton, P.; Bell, S.; Mahy, G.; Grover, Z.; Lewindon, P.; Jones, L.V.; Sewell, K.; Krishnaprasad, K.; et al. Infliximab vs. adalimumab in Crohn’s disease: Results from 327 patients in an Australian and New Zealand observational cohort study. Aliment. Pharmacol. Ther. 2017, 45, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Heien, H.C.; Sangaralingham, L.R.; Schilz, S.R.; Kappelman, M.D.; Shah, N.D.; Loftus Jr., E. V Comparative effectiveness and safety of anti-tumor necrosis factor agents in biologic-naïve patients with Crohn’s disease. Clin. Gastroenterol. Hepatol. 2016, 14, 1120–1129. [Google Scholar] [CrossRef] [Green Version]
- Elvers, K.T.; Wilson, V.J.; Hammond, A.; Duncan, L.; Huntley, A.L.; Hay, A.D.; Werf, E.T. van der Antibiotic-induced changes in the human gut microbiota for the most commonly prescribed antibiotics in primary care in the UK: A systematic review. BMJ Open 2020, 10, e035677. [Google Scholar] [CrossRef]
- Agnes, A.; Puccioni, C.; D’Ugo, D.; Gasbarrini, A.; Biondi, A.; Persiani, R. The gut microbiota and colorectal surgery outcomes: Facts or hype? A narrative review. BMC Surg. 2021, 21, 83. [Google Scholar] [CrossRef]
- Sakurai, T.; Nishiyama, H.; Sakai, K.; De Velasco, M.A.; Nagai, T.; Komeda, Y.; Kashida, H.; Okada, A.; Kawai, I.; Nishio, K.; et al. Mucosal microbiota and gene expression are associated with long-term remission after discontinuation of adalimumab in ulcerative colitis. Sci. Rep. 2020, 10, 19186. [Google Scholar] [CrossRef]
- Sanchis-Artero, L.; Martínez-Blanch, J.F.; Manresa-Vera, S.; Cortés-Castell, E.; Valls-Gandia, M.; Iborra, M.; Paredes-Arquiola, J.M.; Boscá-Watts, M.; Huguet, J.M.; Gil-Borrás, R.; et al. Evaluation of changes in intestinal microbiota in Crohn’s disease patients after anti-TNF alpha treatment. Sci. Rep. 2021, 11, 10016. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Hall, A.B.; Yassour, M.; Sauk, J.; Garner, A.; Jiang, X.; Arthur, T.; Lagoudas, G.K.; Vatanen, T.; Fornelos, N.; Wilson, R.; et al. A novel Ruminococcus gnavus clade enriched in inflammatory bowel disease patients. Genome Med. 2017, 9, 103. [Google Scholar] [CrossRef]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus gnavus, a member of the human gut microbiome associated with Crohn’s disease, produces an inflammatory polysaccharide. Proc. Natl. Acad. Sci. USA 2019, 116, 12672–12677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Zhao, W.; Lan, P.; Mou, X. The microbiome in inflammatory bowel diseases: From pathogenesis to therapy. Protein Cell 2021, 12, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Sankarasubramanian, J.; Ahmad, R.; Avuthu, N.; Singh, A.B.; Guda, C. Gut microbiota and metabolic specificity in ulcerative colitis and Crohn’s disease. Front. Med. 2020, 7, 606298. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, X.; Ghozlane, A.; Hu, H.; Li, X.; Xiao, Y.; Li, D.; Yu, G.; Zhang, T. Characteristics of faecal microbiota in paediatric Crohn’s disease and their dynamic changes during infliximab therapy. J. Crohn’s Colitis 2018, 12, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of human gut microbiota and short-chain fatty acids in response to dietary interventions with three fermentable fibers. mBio 2019, 10, e02566-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.; Giudici, F.; Fiorindi, C.; Ficari, F.; Scaringi, S.; Amedei, A. Immunomodulating activity and therapeutic effects of short chain fatty acids and tryptophan post-biotics in inflammatory bowel disease. Front. Immunol. 2019, 10, 2754. [Google Scholar] [CrossRef] [Green Version]
- Venegas, D.P.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Jayawardena, D.; Dudeja, P.K. Micronutrient deficiency in inflammatory bowel diseases: Cause or effect? Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 707–708. [Google Scholar] [CrossRef]
- Skupsky, J.; Sabui, S.; Hwang, M.; Nakasaki, M.; Cahalan, M.D.; Said, H.M. Biotin supplementation ameliorates murine colitis by preventing NF-κB activation. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 557–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skupsky, J.; Sabui, S.; Nakasaki, M.; Chen, J.; Cahalan, M.; Said, H. P130 biotin supplementation ameliorates murine colitis by maintaining intestinal mucosal integrity. Gastroenterology 2019, 156, S90. [Google Scholar] [CrossRef]
- Singh, V.; Yeoh, B.S.; Xiao, X.; Kumar, M.; Bachman, M.; Borregaard, N.; Joe, B.; Vijay-Kumar, M. Interplay between enterobactin, myeloperoxidase and lipocalin 2 regulates E. Coli survival in the inflamed gut. Nat. Commun. 2015, 6, 7113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, A.K.; Mitchell, A.M. Enterobacterial common antigen: Synthesis and function of an enigmatic molecule. mBio 2020, 11, e01914-20. [Google Scholar] [CrossRef] [PubMed]
- Bull, D.M.; Ignaczak, T.F. Enterobacterial common antigen-inducedlymphocyte reactivity in inflammatory bowel disease. Gastroenterology 1973, 64, 43–50. [Google Scholar] [CrossRef]
- Makala, H.P.; Mayer, H. Enterobacterial common antigen. Bacteriol. Rev. 1976, 40, 591–632. [Google Scholar] [CrossRef]
- Auchtung, T.A.; Fofanova, T.Y.; Stewart, C.J.; Nash, A.K.; Wong, M.C.; Gesell, J.R.; Auchtung, J.M.; Ajami, N.J.; Petrosino, J.F. Investigating colonization of the healthy adult gastrointestinal tract by fungi. mSphere 2018, 3, e00092-18. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, C.; Dollive, S.; Grunberg, S.; Chen, J.; Li, H.; Wu, G.D.; Lewis, J.D.; Bushman, F.D. Archaea and fungi of the human gut microbiome: Correlations with diet and bacterial residents. PLoS ONE 2013, 8, e0066019. [Google Scholar] [CrossRef] [Green Version]
- Huseyin, C.E.; O’Toole, P.W.; Cotter, P.D.; Scanlan, P.D. Forgotten fungi-the gut mycobiome in human health and disease. FEMS Microbiol. Rev. 2017, 41, 479–511. [Google Scholar] [CrossRef] [Green Version]
- Kakoullis, L.; Pantzaris, N.D.; Platanaki, C.; Lagadinou, M.; Papachristodoulou, E.; Velissaris, D. The use of IgM-enriched immunoglobulin in adult patients with sepsis. J. Crit. Care 2018, 47, 30–35. [Google Scholar] [CrossRef]
- Daien, C.I.; Gailhac, S.; Mura, T.; Combe, B.; Hahne, M.; Morel, J. High levels of memory B cells are associated with response to a first tumor necrosis factor inhibitor in patients with rheumatoid arthritis in a longitudinal prospective study. Arthritis Res. Ther. 2014, 16, R95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmermans, W.M.C.; Van Laar, J.A.M.; Van Der Houwen, T.B.; Kamphuis, L.S.J.; Bartol, S.J.W.; Lam, K.H.; Ouwendijk, R.J.; Sparrow, M.P.; Gibson, P.R.; Van Hagen, P.M.; et al. B-cell dysregulation in Crohn’s disease is partially restored with infliximab therapy. PLoS ONE 2016, 11, e0160103. [Google Scholar] [CrossRef] [PubMed]
- Tabaqchali, S.; O’Donoghue, D.P.; Bettelheim, K.A. Escherichia coli antibodies in patients with inflammatory bowel disease. Gut 1978, 19, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Alexander, K.L.; Zhao, Q.; Reif, M.; Rosenberg, A.F.; Mannon, P.J.; Duck, L.W.; Elson, C.O. Human microbiota flagellins drive adaptive immune responses in Crohn’s disease. Gastroenterology 2021, 161, 522–535.e6. [Google Scholar] [CrossRef]
- Palm, N.W.; De Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut microbiota-induced immunoglobulin G controls systemic infection by symbiotic bacteria and pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Dubé, P.E.; Punit, S.; Brent Polk, D. Redeeming an old foe: Protective as well as pathophysiological roles for tumor necrosis factor in inflammatory bowel disease. Am. J. Physiol.-Gastrointest. Liver Physiol. 2015, 308, G161–G170. [Google Scholar] [CrossRef] [Green Version]
- Gálvez, J. Role of Th17 cells in the pathogenesis of human IBD. ISRN Inflamm. 2014, 2014, 928461. [Google Scholar] [CrossRef] [Green Version]
- Kojouharoff, G.; Hans, W.; Obermeler, F.; Männel, D.N.; Andus, T.; Schölmerich, J.; Gross, V.; Falk, W. Neutralization of tumour necrosis factor (TNF) but not of IL-1 reduces inflammation in chronic dextran sulphate sodium-induced colitis in mice. Clin. Exp. Immunol. 1997, 107, 353–358. [Google Scholar] [CrossRef]
- Noti, M.; Corazza, N.; Mueller, C.; Berger, B.; Brunner, T. TNF suppresses acute intestinal inflammation by inducing local glucocorticoid synthesis. J. Exp. Med. 2010, 207, 1057–1066. [Google Scholar] [CrossRef]
- Hueber, W.; Sands, B.E.; Lewitzky, S.; Vandemeulebroecke, M.; Reinisch, W.; Higgins, P.D.R.; Wehkamp, J.; Feagan, B.G.; Yao, M.D.; Karczewski, M.; et al. Secukinumab, a human anti-IL-17A monoclonal antibody, for moderate to severe Crohn’s disease: Unexpected results of a randomised, double-blind placebo-controlled trial Correspondence. Gut 2012, 61, 1693–1700. [Google Scholar] [CrossRef] [Green Version]
- Vernero, M.; Astegiano, M.; Ribaldone, D.G. New onset of inflammatory bowel disease in three patients undergoing IL-17A inhibitor secukinumab: A case series. Am. J. Gastroenterol. 2019, 114, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Walters, H.M.; Pan, N.; Lehman, T.J.A.; Adams, A.; Kalliolias, G.D.; Zhu, Y.S.; Santiago, F.; Nguyen, J.; Sitaras, L.; Cunningham-Rundles, S.; et al. The impact of disease activity and tumour necrosis factor-α inhibitor therapy on cytokine levels in juvenile idiopathic arthritis. Clin. Exp. Immunol. 2016, 184, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, A.; Calhan, T.; Cengiz, M.; Kahraman, R.; Aydin, K.; Ozdil, K.; Korachi, M.; Sokmen, H.M. Serum interleukin 17 levels in patients with Crohn’s disease: Real life data. Dis. Markers 2014, 2014, 690853. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Monin, L.; Castillo, P.; Elsegeiny, W.; Horne, W.; Eddens, T.; Vikram, A.; Good, M.; Schoenborn, A.A.; Bibby, K.; et al. Intestinal interleukin-17 receptor signaling mediates reciprocal control of the gut microbiota and autoimmune inflammation. Immunity 2016, 44, 659–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 37) | CD (n = 17) Week 0 | CD (n = 17) Week 38 | ∆ CD (p-Value) | CD vs. HC (p-Value) | UC (n = 10) Week 0 | UC (n = 10) Week 38 | ∆ UC (p-Value) | UC vs. HC (p-Value) | |
|---|---|---|---|---|---|---|---|---|---|
| Male: female | 15:22 | 5:12 | 0.8042 | 2:8 | - | 0.8684 | |||
| Age | 36.5 (28.8, 41.3) | 35.0 (26.5, 44.0) | - | 0.7800 | 31.0 (26.0, 41.3) | - | 0.4228 | ||
| BMI | 21.8 (20.5, 26.2) | 22.2 (20.8, 26.9) | - | 0.8274 | 21.3 (19.2, 24.4) | - | 0.1901 | ||
| Disease duration in years | - | 4.0 (2.0, 13.5) | - | - | 2.5 (1.0, 7.5) | - | - | ||
| Age at diagnosis | - | 26 (22.5, 35) | - | - | 27 (21.5, 31) | - | - | ||
| Remitters | - | 11/17 | 13/17 | - | - | 0/10 | 5/10 | - | - |
| HBI/pMayo | - | 3 (0, 6) | 1 (0.5, 5) | 0.3931 | - | 7 (3.8, 8.3) | 2.5 (0, 4.3) | 0.0039 | - |
| CRP (mg/L) | 0.9 (0.4, 1.9) | 4.2 (1.0, 8.9) | 1.3 (0.5, 5) | 0.0225 | 0.0037 | 4.1 (0.9, 22.3) | 0.95 (0.6, 2.8) | 0.1641 | 0.0124 |
| WBC | 5.9 (4.9, 6.9) | 8.0 (6.2, 11.0) | 5.9 (5, 7.8) | 0.0089 | 0.0135 | 10.4 (6.1, 12.3) | 5.9 (4.4, 7.3) | 0.0371 | 0.0071 |
| PLT | 247 (218, 272) | 328 (269, 416) | 283 (220, 388) | 0.0697 | 0.0007 | 276 (251, 387) | 278 (222, 310) | 0.4160 | 0.1136 |
| Ferritin (µg/L) | NA | 48.1 (19.8, 80.0) | 30.9 (12.3, 56.8) | 0.3778 | NA | 28.0 (11.7, 52.2) | 41.0 (9.5, 78.6) | 0.9219 | NA |
| Hb (g/L) | 141 (134, 158) | 130 (123, 150) | 134 (127, 144) | 0.4411 | 0.0495 | 128 (109, 1367) | 134 (121, 146) | 0.0918 | 0.0067 |
| FC (μg/g) | NA | 407 (148, 1028) | 162 (38, 284) | 0.0181 | NA | 661 (424, 2838) | 229 (59, 828) | 0.0039 | NA |
| Week 0 | Week 38 | HC | Week 0 vs. HC | Week 38 vs. HC | Week 0 vs. Week 38 | ||
|---|---|---|---|---|---|---|---|
| IgM | CD | 2.1 (1.3; 3.4) | 2.4 (1.4; 7.6) | 1.1 (0.8; 1.8) | 0.020 | <0.001 | <0.001 |
| UC | 2.6 (0.9; 4.0) | 1.3 (1.0; 3.2) | ns | ns | ns | ||
| IgA | CD | 1.9 (0.5; 5.0) | 2.2 (0.6; 6.6) | 0.7 (0.5; 1.4) | 0.009 | 0.010 | ns |
| UC | 2.5 (1.3; 4.4) | 3.2 (1.3; 5.7) | 0.039 | 0.024 | ns | ||
| IgG | CD | 15.9 (8.1; 35.1) | 11.2 (5.3; 20.4) | 1.1 (0.8; 1.8) | <0.001 | 0.002 | ns |
| UC | 10.5 (6.5; 18.1) | 10.6 (5.7; 59.8) | 0.019 | <0.001 | ns |
| CD w0 vs. HC | CD w38 vs. HC | CD w0 vs. w38 | UC w0 vs. HC | UC w38 vs. HC | UC w0 vs. w38 | |
|---|---|---|---|---|---|---|
| Lactobacillus | ns | ns | ↗ p = 0.005 | ns | ns | ns |
| Bifidobacterium | ns | ns | ↗ p = 0.010 | ns | ns | ns |
| Blautia | ns | ns | ↗ p = 0.005 | ns | ns | ns |
| Roseburia | ns | ns | ↗ p < 0.001 | ns | ns | ns |
| Faecalibacterium | ns | ns | ns | ns | ns | ns |
| Bacteroides | ns | ns | ↗ p = 0.005 | ns | ns | ns |
| Escherichia | ns | ns | ↗ p = 0.003 | ns | ns | ns |
| Prevotella | ns | ns | ↗ p = 0.005 | ns | ns | ns |
| Ruminococcus | ns | ns | ↗ p = 0.001 | ns | ns | ns |
| Eubacterium | ns | ns | ↗ p < 0.001 | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schierova, D.; Roubalova, R.; Kolar, M.; Stehlikova, Z.; Rob, F.; Jackova, Z.; Coufal, S.; Thon, T.; Mihula, M.; Modrak, M.; et al. Fecal Microbiome Changes and Specific Anti-Bacterial Response in Patients with IBD during Anti-TNF Therapy. Cells 2021, 10, 3188. https://doi.org/10.3390/cells10113188
Schierova D, Roubalova R, Kolar M, Stehlikova Z, Rob F, Jackova Z, Coufal S, Thon T, Mihula M, Modrak M, et al. Fecal Microbiome Changes and Specific Anti-Bacterial Response in Patients with IBD during Anti-TNF Therapy. Cells. 2021; 10(11):3188. https://doi.org/10.3390/cells10113188
Chicago/Turabian StyleSchierova, Dagmar, Radka Roubalova, Martin Kolar, Zuzana Stehlikova, Filip Rob, Zuzana Jackova, Stepan Coufal, Tomas Thon, Martin Mihula, Martin Modrak, and et al. 2021. "Fecal Microbiome Changes and Specific Anti-Bacterial Response in Patients with IBD during Anti-TNF Therapy" Cells 10, no. 11: 3188. https://doi.org/10.3390/cells10113188