
Clover Species Specific Influence on Microbial Abundance and Associated Enzyme Activities in Rhizosphere and Non-Rhizosphere Soils

 , , ,
, , ,  , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
- The biological properties of rhizosoil would exhibit generally stronger relation or correlation among each other (due to less diverse and more specified microbial community) than the biological properties of bulk soil (which likely exert much higher microbial diversity).
- The nutrient content of plant biomass would correlate significantly more positively with the biological properties of rhizosoil (due to the mutual interaction) than with biological properties of bulk soil (because of weak or no interaction).
- There would be a significant species-specific effect of tested types of legumes on both biological properties of soil (enzyme activity, microbial abundance) and nutrient content in the plant biomass.
2. Materials and Methods
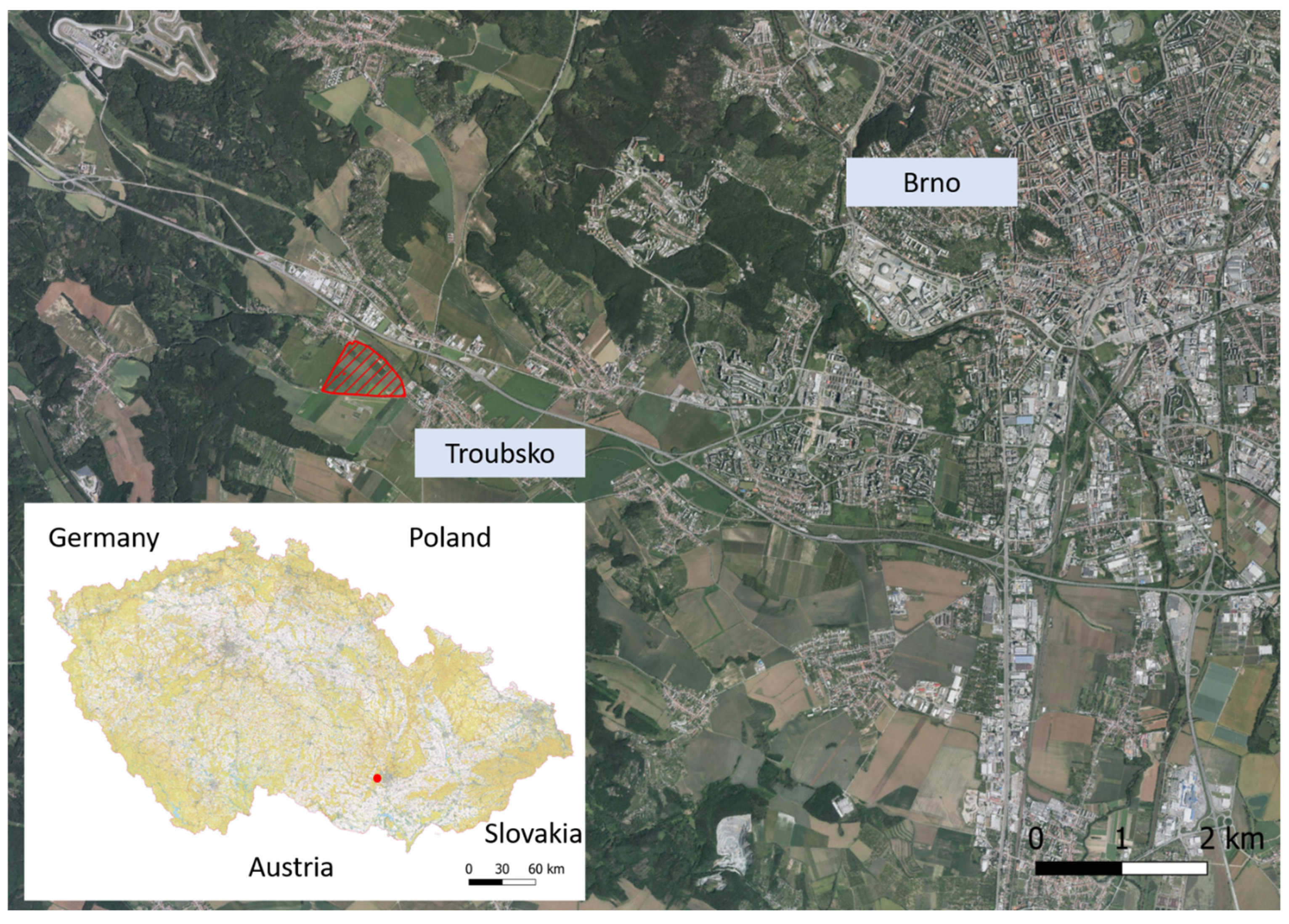
2.1. Site Description and Experimental Setup
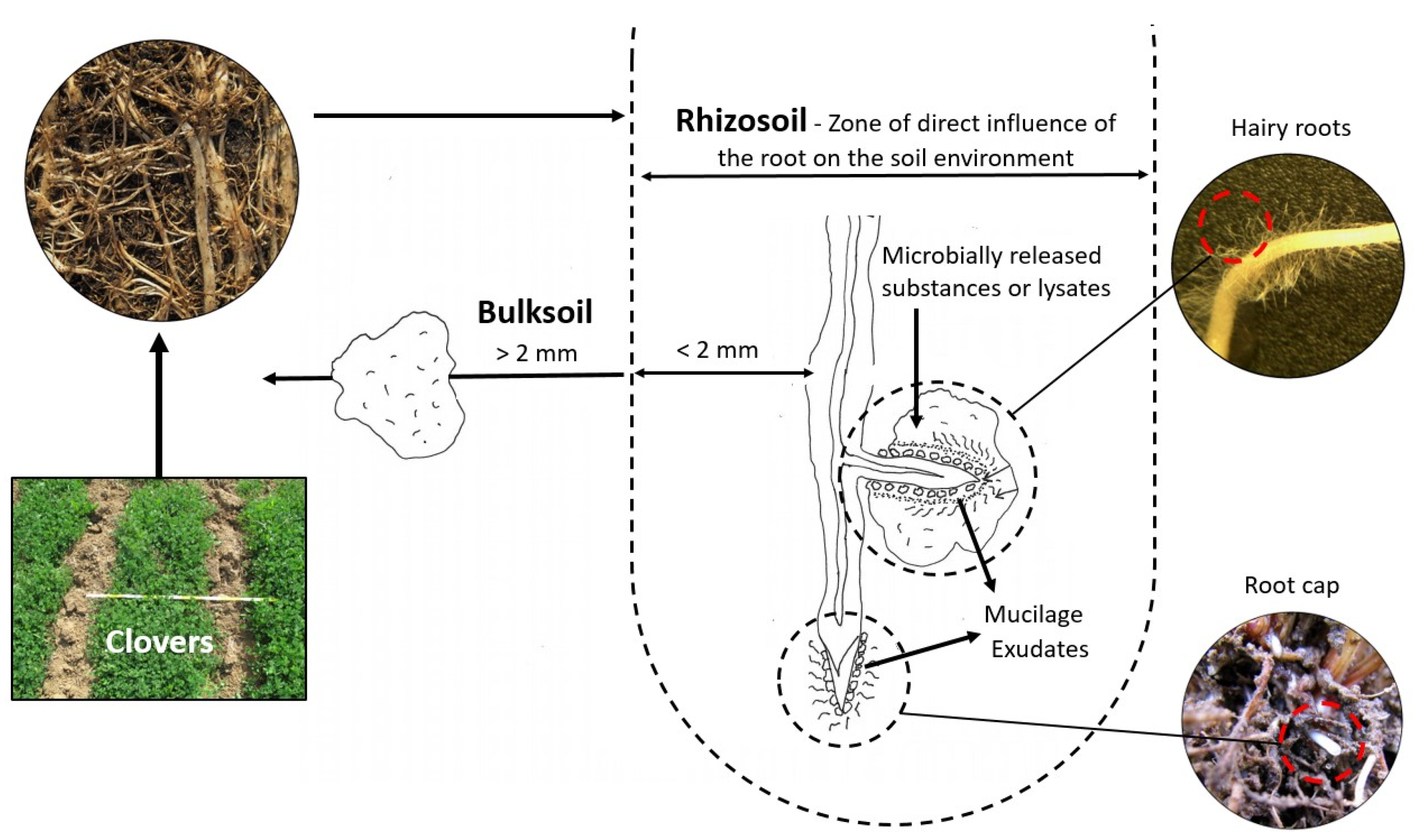
2.2. Soil Sampling and Processing
2.3. Plant Biomass
2.4. Soil Enzyme Activities
2.5. DNA Extraction and Real-Time qPCR
2.6. Statistical Analyses
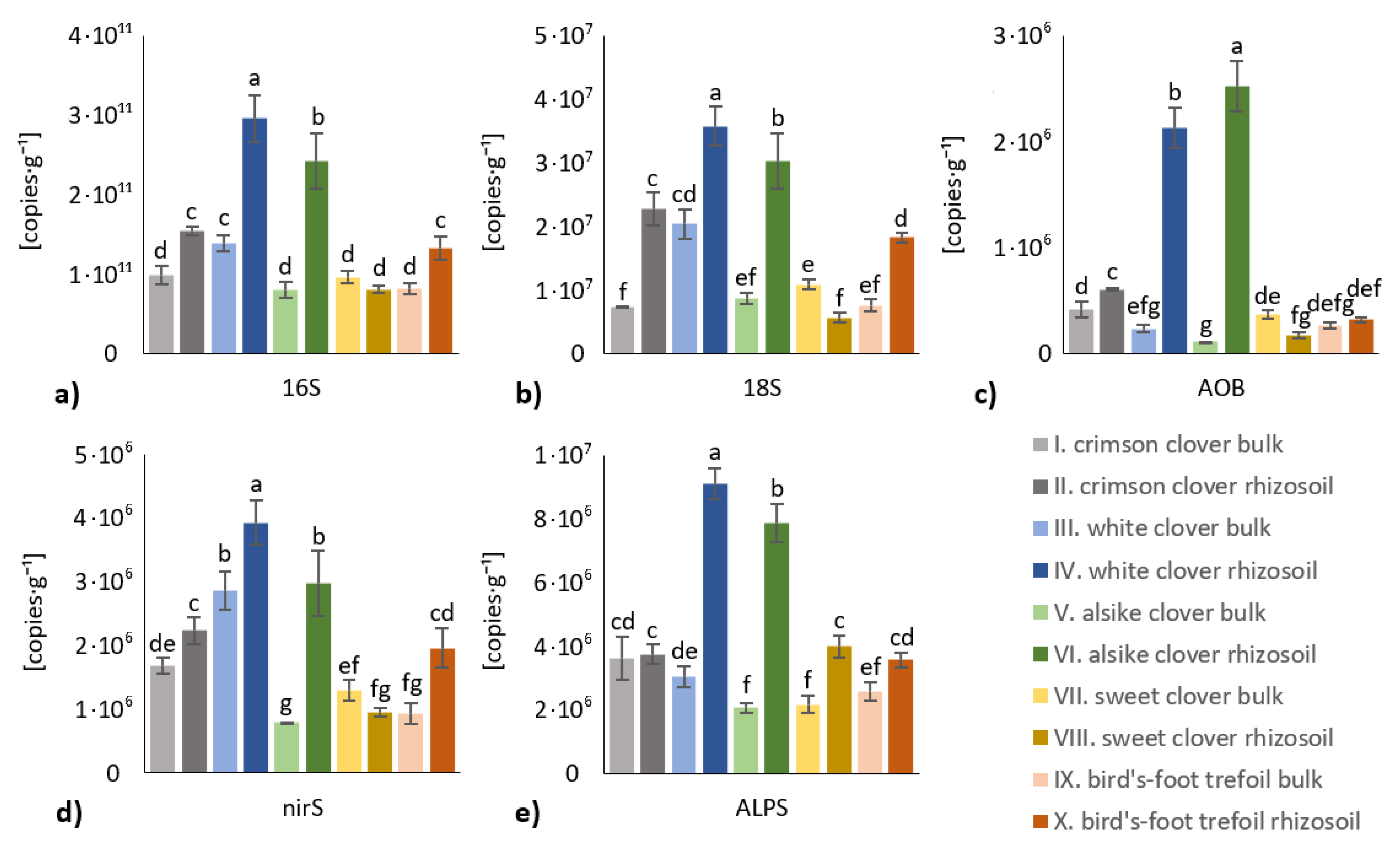
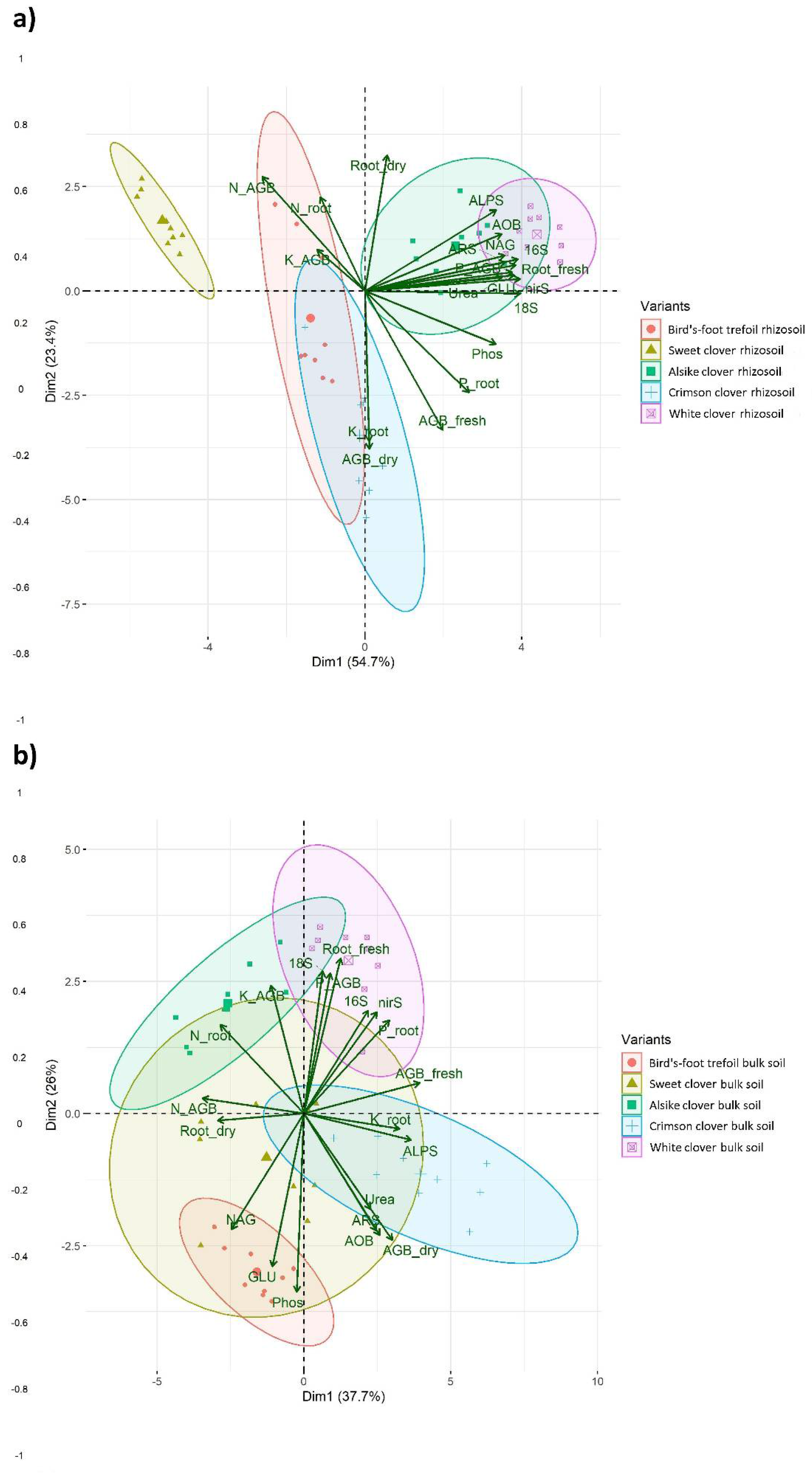
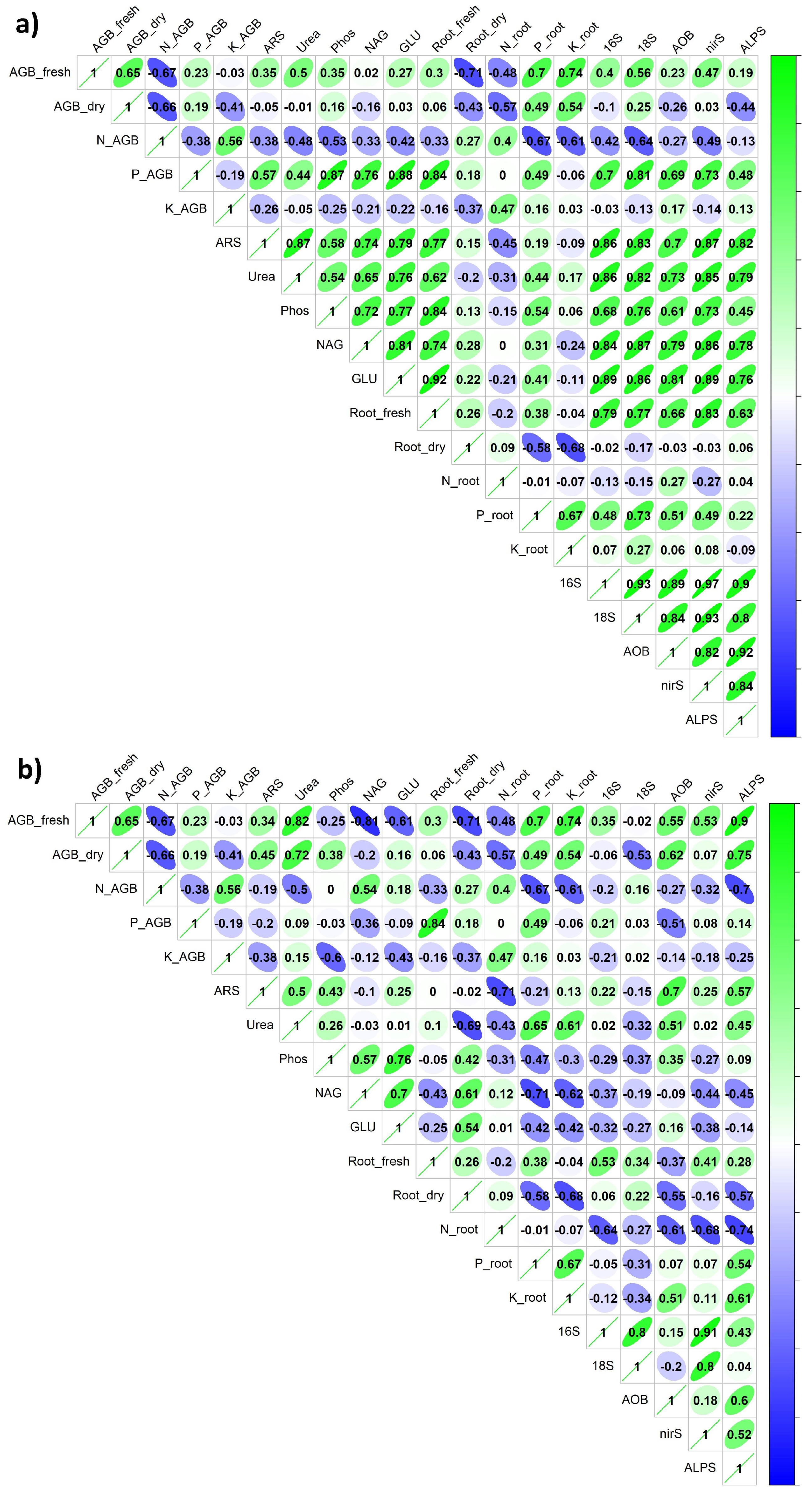
3. Results
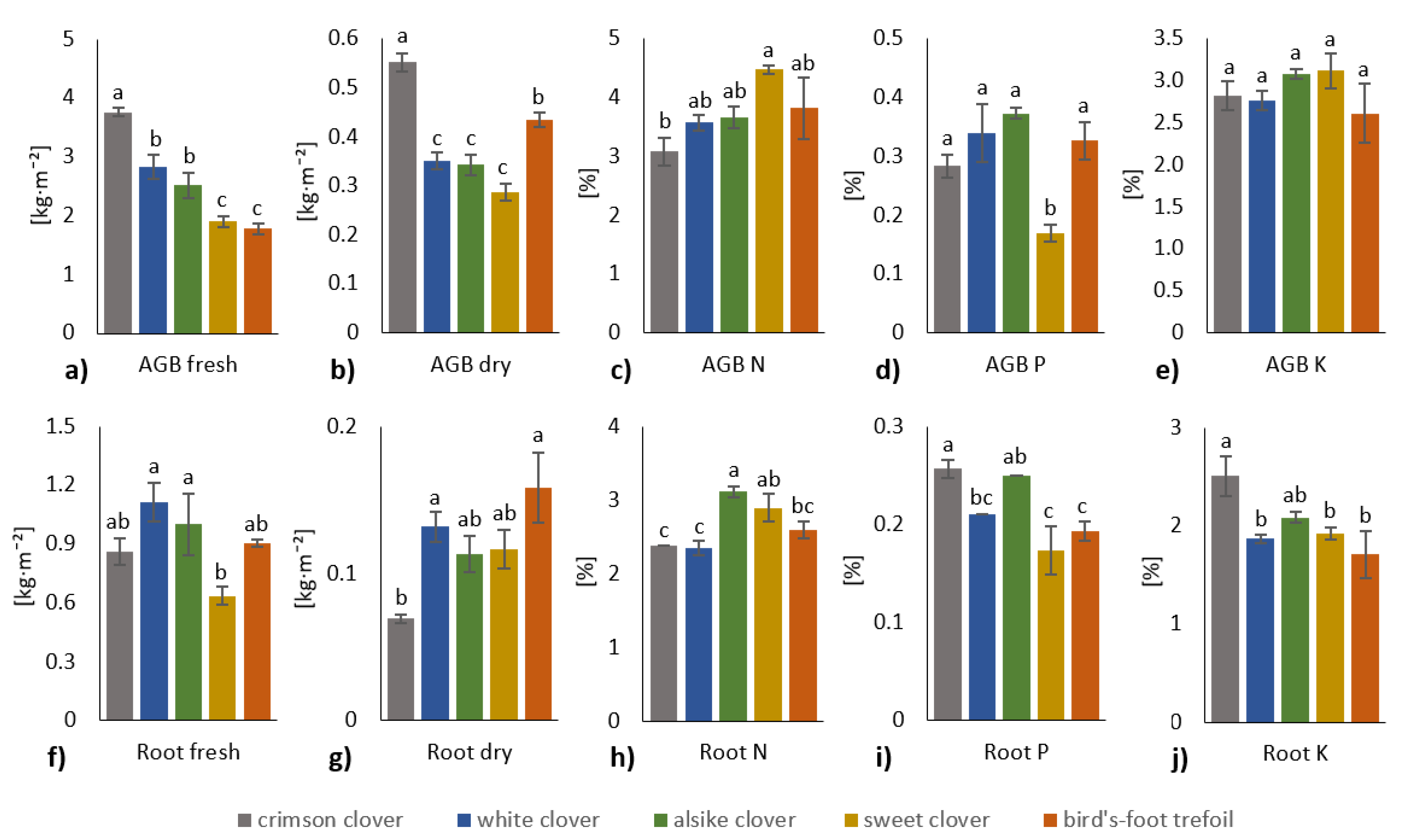
3.1. Plant Biomass and Nutrient Content Relation to Rhizosoil Microbial Abundance and Activity
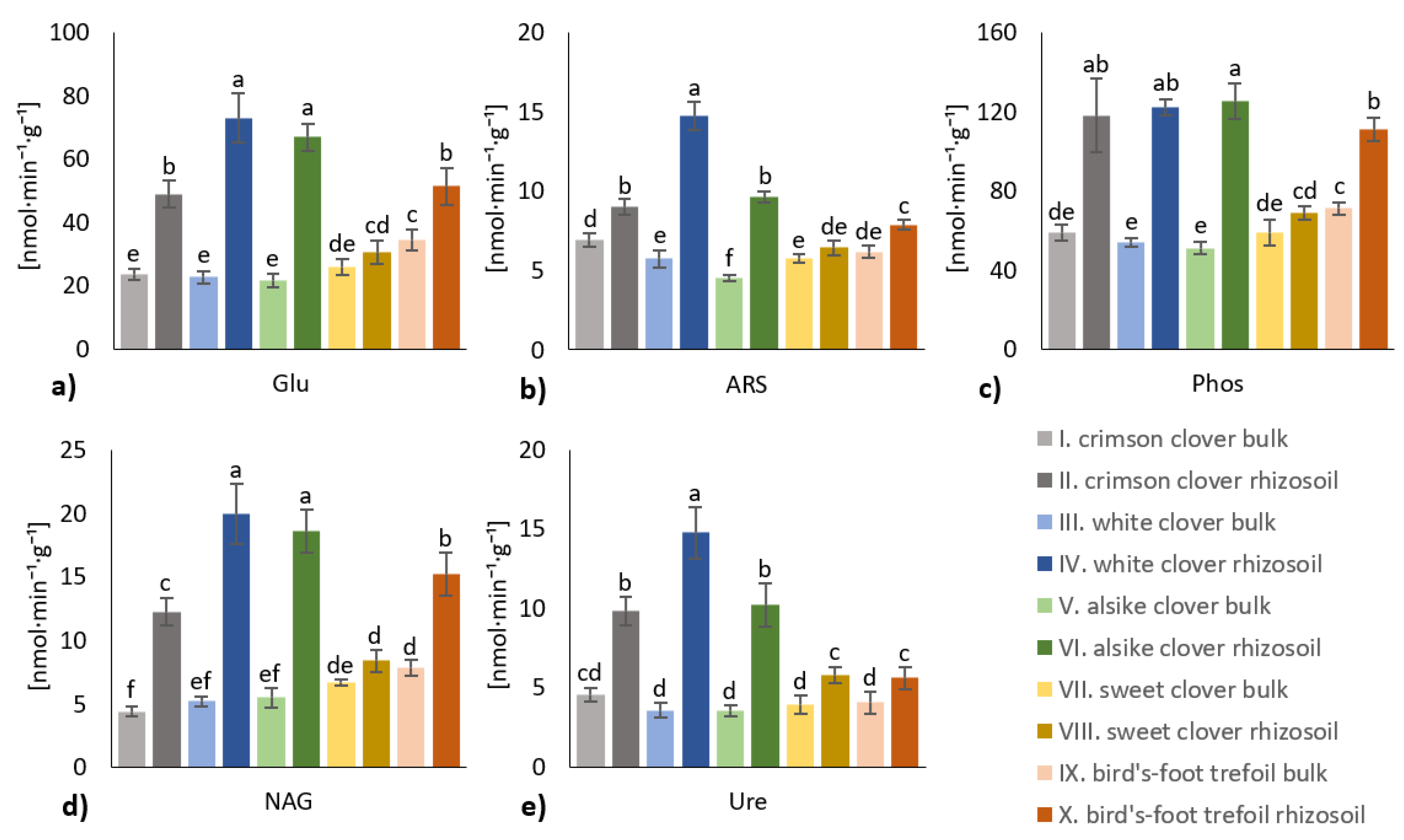
3.2. Enzymes Activities in Rhizosoil and Bulk Soil
3.3. Microbial Biomass and Abundance in Rhizosoil and Bulk Soil
4. Discussion
4.1. Plant Biomass and Nutrient Content Relation to Rhizosoil Microbial Abundance and Activity
4.2. Microbial Soil Propertied in Rhizosoil and Bulk Soil, and Their Relation to Plant Traits
5. Conclusions
- The soil biological properties were generally more affected in the rhizosoil than in the bulk soil, although the individual legume crop variants differed in the rate and significance of the differential impact on either rhizosoil or bulk soil. The most significantly affected properties were ammonium oxidizing bacteria (AOB) and P-solubilizing microbiota in the rhizosoil of white clover and alsike clover variants.
- The biological properties of rhizosoil showed generally high synergism among each other and a significant effect on the plant qualitative and quantitative properties.
- However, the antagonism among N and P + K transfer in the rhizosoil to plants in the term of nutrient uptake and acquisition by either shoots or roots of tested legume crops, and this antagonism also influenced AGB and root biomass.
- Species-specific effect of tested legume crops was evaluated: the most significant species-specific effect showed white clover and alsike clover, whereas the least significant species-specific impact exerted sweet clover.
- The experimental design may be applied as a suitable approach for the investigation of the plant species-specific effect on the soil properties.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AGB | Aboveground biomass |
| AGB N | nitrogen content in dry AGB |
| AGB P | phosphorus content in dry AGB |
| AGB K | potassium content in dry AGB |
| AHL | N-acyl homoserine lactone |
| AOB | ammonia oxidizing bacteria |
| ALPS | phosphorus solubilizing microbes |
| ARS | arylsulfatase |
| FAO | Food and Agriculture Organization of the United Nations |
| GLU | β-glucosidase |
| NAG | N-acetyl-β-D-glucosaminidase |
| nirS | nitrate reductase/denitrifiers |
| PCA | principal component analysis |
| Phos | phosphatase |
| r | Pearson’s correlation coefficient |
| root N | nitrogen content in dry root biomass |
| root P | phosphorus content in dry root biomass |
| root K | potassium content in dry root biomass |
| Ure | urease |
Appendix A


References
- West, P.C.; Gerber, J.S.; Engstrom, P.M.; Mueller, N.D.; Brauman, K.A.; Carlson, K.M.; Cassidy, E.S.; Johnston, M.; MacDonald, G.K.; Ray, D.K.; et al. Leverage points for improving global food security and the environment. Science 2014, 345, 325–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, M.; Naseer, I.; Hussain, A.; Zahid Mumtaz, M.; Mustafa, A.; Hilger, T.H.; Zahir, Z.A.; Minggang, X. Appraising Endophyte—Plant Symbiosis for Improved Growth, Nodulation, Nitrogen Fixation and Abiotic Stress Tolerance: An Experimental Investigation with Chickpea (Cicer arietinum L.). Agronomy 2019, 9, 621. [Google Scholar] [CrossRef] [Green Version]
- Rafique, M.; Naveed, M.; Mustafa, A.; Akhtar, S.; Munawar, M.; Kaukab, S.; Ali, H.M.; Siddiqui, M.H.; Salem, M.Z.M. The Combined Effects of Gibberellic Acid and Rhizobium on Growth, Yield and Nutritional Status in Chickpea (Cicer arietinum L.). Agronomy 2021, 11, 105. [Google Scholar] [CrossRef]
- Bationo, A.; Lompo, F.; Koala, S. Research on nutrient flows and balances in west Africa: State-of-the-art1Paper contributes to EC INCO-DC project IC18-CT96-0092 (Spatial and temporal variation of soil nutrient stocks and management in sub-Saharan African farming systems).1. Agric. Ecosyst. Environ. 1998, 71, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Wik, M.; Pingali, P.; Broca, S. Global Agricultural Performance: Past Trends and Future Prospects; World Development Report; World Bank: Washington, DC, USA, 2008. [Google Scholar]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Elucidating the diversity and plant growth promoting attributes of wheat (Triticum aestivum) associated acid tolerant bacteria from southern hills zone of India. Natl. J. Life Sci. 2013, 10, 219–226. [Google Scholar]
- Refsgaard, K.; Halberg, N.; Kristensen, E.S. Energy utilization in crop and dairy production in organic and conventional livestock production systems. Agric. Syst. 1998, 57, 599–630. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.; Kiel, P. Integrated utilisation of green biomass in the green biorefinery. Ind. Crop. Prod. 2000, 11, 129–137. [Google Scholar] [CrossRef]
- Snapp, S.S.; Swinton, S.M.; Labarta, R.; Mutch, D.; Black, J.R.; Leep, R.; Nyiraneza, J.; O’Neil, K. Evaluating cover crops for benefits, costs and performance within cropping system niches. Agron. J. 2005, 97, 322–332. [Google Scholar] [CrossRef]
- Fustec, J.; Lesuffleur, F.; Mahieu, S.; Cliquet, J.-B. Nitrogen rhizodeposition of legumes. A review. Agron. Sustain. Dev. 2010, 30, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Naveed, M.; Mustafa, A.; Abbas, A. The Good, the Bad, and the Ugly of Rhizosphere Microbiome. In Probiotics and Plant Health; Springer: Singapore, 2017; pp. 253–290. [Google Scholar]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 2010, 34, 150–170. [Google Scholar] [CrossRef]
- Veliz-Vallejos, D.F.; Kawasaki, A.; Mathesius, U. The Presence of Plant-Associated Bacteria Alters Responses to N-acyl Homoserine Lactone Quorum Sensing Signals that Modulate Nodulation in Medicago Truncatula. Plants 2020, 9, 777. [Google Scholar] [CrossRef] [PubMed]
- Ortas, I. Determination of the extent of rhizosphere soil. Commun. Soil Sci. Plant Anal. 1997, 28, 1767–1776. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Augustin, J.; Merbach, W. Plant rhizodeposition—An important source for carbon turnover in soils. J. Plant Nutr. Soil Sci. 2002, 165, 397–407. [Google Scholar] [CrossRef]
- Elbl, J.; Zahora, J. The Comparison of Microbial Activity in Rhizosphere and Non-Rhizosphere Soil Stressed by Drought; Mendel Univ Brno, Fac Agronomy: Brno, Czech Republic, 2014. [Google Scholar]
- Walker, T.S.; Bais, H.P.; Grotewold, E.; Vivanco, J.M. Root exudation and rhizosphere biology. Plant. Physiol. 2003, 132, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.R.; Ramakrishnan, K.; Walshaw, J.; Heavens, D.; Alston, M.; Swarbreck, D.; Osbourn, A.; Grant, A.; Poole, P.S. Comparative metatranscriptomics reveals kingdom level changes in the rhizosphere microbiome of plants. ISME J. 2013, 7, 2248–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, J.; Lotter, D.; Meyer, A.; Kleinert, A.; Pérez-Fernández, M.; Valentine, A. Variation in rhizosphere nutrient cycling affects the source of nitrogen acquisition in wild and cultivated Aspalathus linearis (N.L.Burm.) R.Dahlgren plants. Appl. Soil Ecol. 2018, 130, 26–33. [Google Scholar] [CrossRef]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de los Reyes, A.M.M.; Ocampo, E.T.M.; Manuel, M.C.C.; Mendoza, B.C. Analysis of the Bacterial and Fungal Community Profiles in Bulk Soil and Rhizospheres of Three Mungbean [Vigna radiata (L.) R. Wilczek] Genotypes through PCR-DGGE. Int. Lett. Nat. Sci. 2020, 77, 1–26. [Google Scholar] [CrossRef]
- Di Cenzo, G.C.; Checcucci, A.; Bazzicalupo, M.; Mengoni, A.; Viti, C.; Dziewit, L.; Finan, T.M.; Galardini, M.; Fondi, M. Metabolic modelling reveals the specialization of secondary replicons for niche adaptation in Sinorhizobium meliloti. Nat. Commun. 2016, 7, 12219. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Negassa, W.C.; Guber, A.K.; Hildebrandt, B.; Marsh, T.L.; Rivers, M.L. Intra-aggregate Pore Structure Influences Phylogenetic Composition of Bacterial Community in Macroaggregates. Soil Sci. Soc. Am. J. 2014, 78, 1924–1939. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Grayston, S.J.; Wang, S.Q.; Campbell, C.D.; Edwards, A.C. Selective influence of plant species on microbial diversity in the rhizosphere. Soil Biol. Biochem. 1998, 30, 369–378. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Svenning, M. Competition for Nodule Occupancy Between Introduced Strains of Rhizobium leguminosarum Biovar trifolii and its Influence on Plant Production. Ann. Bot. 2001, 88, 781–787. [Google Scholar] [CrossRef]
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Jackson, R.W.; Mauchline, T.H.; Hirsch, P.R.; Shaw, L.J.; Doring, T.F.; Jones, H.E. Is there sufficient Ensifer and Rhizobium species diversity in UK farmland soils to support red clover (Trifolium pratense), white clover (T. repens), lucerne (Medicago sativa) and black medic (M. lupulina)? Appl. Soil Ecol. 2017, 120, 35–43. [Google Scholar] [CrossRef]
- de la Fuente Canto, C.; Simonin, M.; King, E.; Moulin, L.; Bennett, M.J.; Castrillo, G.; Laplaze, L. An extended root phenotype: The rhizosphere, its formation and impacts on plant fitness. Plant J. 2020, 103, 951–964. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Reference Base for Soil Resources. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Brtnicky, M.; Hammerschmiedt, T.J.; Elbl Kintl, A.; Skulcova, L.; Radziemska, M.; Latal, O.; Baltazar, T.; Kobzova, E.; Holatko, J. The Potential of Biochar Made from Agricultural Residues to Increase Soil Fertility and Microbial Activity: Impacts on Soils with Varying Sand Content. Agronomy 2021, 11, 1174. [Google Scholar] [CrossRef]
- Capolongo, A.; Barresi, A.A.; Rovero, G. Freeze-drying of lignin peroxidase: Influence of lyoprotectants on enzyme activity and stability. J. Chem. Technol. Biotechnol. 2003, 78, 56–63. [Google Scholar] [CrossRef]
- Iocoli, G.A.; Zabaloy, M.C.; Pasdevicelli, G.; Gomez, M.A. Use of biogas digestates obtained by anaerobic digestion and co-digestion as fertilizers: Characterization, soil biological activity and growth dynamic of Lactuca sativa L. Sci. Total. Environ. 2019, 647, 11–19. [Google Scholar] [CrossRef]
- ISO_20130. Soil Quality—Measurement of Enzyme Activity Patterns in Soil Samples Using Colorimetric Substrates in Micro-Well Plates; International Organization for Standardization: Geneva, Switzerland, 2018. [Google Scholar]
- Verchot, L.; Borelli, T. Application of -nitrophenol (NP) enzyme assays in degraded tropical soils. Soil Biol. Biochem. 2005, 37, 625–633. [Google Scholar] [CrossRef]
- Whalen, J.K.; Warman, P.R. Arylsulfatase activity in soil and soil extracts using natural and artificial substrates. Biol. Fertil. Soils 1996, 22, 373–378. [Google Scholar] [CrossRef]
- Jackson, C.R.; Tyler, H.L.; Millar, J.J. Determination of microbial extracellular enzyme activity in waters, soils, and sediments using high throughput microplate assays. J. Vis. Exp. 2013, 2013, e50399. [Google Scholar] [CrossRef] [Green Version]
- Cordero, I.; Snell, H.; Bardgett, R.D. High throughput method for measuring urease activity in soil. Soil Biol. Biochem. 2019, 134, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandeler, E.; Deiglmayr, K.; Tscherko, D.; Bru, D.; Philippot, L. Abundance of narG, nirS, nirK, and nosZ genes of denitrifying bacteria during primary successions of a glacier foreland. Appl. Environ. Microbiol. 2006, 72, 5957–5962. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Xia, Y.; Sun, Q.; Liu, K.; Chen, X.; Ge, T.; Zhu, B.; Zhu, Z.; Zhang, Z.; Su, Y. Effects of long-term fertilization on phoD-harboring bacterial community in Karst soils. Sci. Total. Environ. 2018, 628–629, 53–63. [Google Scholar] [CrossRef]
- R_CORE_TEAM. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Mendiburu, D.F. Agricolae: Statistical Procedures for Agricultural Research; R Package Version 1.2-3; 2015; Available online: http://CRAN.R-project.org/package=agricolae (accessed on 29 October 2021).
- Beaujean, A.A. R Package for Baylor University Educational Psychology Quantitative Courses; BaylorEdPsych; 2012; Available online: https://rdrr.io/cran/BaylorEdPsych/ (accessed on 29 October 2021).
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses; 2017; Available online: https://rpkgs.datanovia.com/factoextra/index.html (accessed on 29 October 2021).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.G.; Carl, P. Performance Analytics: Econometric Tools for Performance and Risk Analysis; 2020; Available online: https://rdrr.io/cran/PerformanceAnalytics/ (accessed on 29 October 2021).
- Thilakarathna, M.S.; Papadopoulos, Y.A. Nitrogen fixation and transfer of red clover genotypes under legume-grass forage based production systems. Nutr. Cycl. Agroecosyst. 2016, 106, 233–247. [Google Scholar] [CrossRef]
- Sekeran, U.; Loya, R.J. Intercropping of kura clover (Trifolium ambiguum M. Bieb) with prairie cordgrass (Spartina pectinata Link.) enhanced soils biochemical activities and microbial community structure. Appl. Soil Ecol. 2019, 147, 1–10. [Google Scholar] [CrossRef]
- Kiers, E.T.; West, S.K.; Denison, R.F. Maintaining cooperation in the legume-rhizobia symbiosis: Identifying selection pressures and mechanisms. In Nitrogen-Fixing Leguminous Symbioses; Springer: Dordrecht, The Netherlands, 2008; pp. 59–76. [Google Scholar]
- Silveira, M.L.; Kohmann, M.M. Maintaining soil fertility and health for sustainable pastures. In Management Strategies for Sustainable Cattle Production in Southern Pastures; Academic Press: London, UK, 2020; pp. 35–58. [Google Scholar]
- Chtouki, M.; Naciri, R.; Soulaimani, A.; Zeroual, Y.; El Gharous, M.; Oukarroum, A. Effect of Cadmium and Phosphorus Interaction on Tomato: Chlorophyll a Fluorescence, Plant Growth, and Cadmium Translocation. Water Air Soil Pollut. 2021, 232, 11. [Google Scholar] [CrossRef]
- Zhang, F.; Niu, J.; Zhang, W.; Chen, X.; Li, C.; Yuan, L.; Xie, J. Potassium nutrition of crops under varied regimes of nitrogen supply. Plant Soil 2010, 335, 21–34. [Google Scholar] [CrossRef]
- Lee, Y.; Hwanga, T.Y.; Lee, S.; Shinogi, Y.; Oh, T.K.; Sung, J. Nutrient-Specific Variation of C-N Metabolism in the Leaves and Roots of Bell Pepper (Capsicum annunm. L) in Response to Macronutrient Deficiency. J. Fac. Agric. Kyushu Univ. 2020, 65, 25–34. [Google Scholar] [CrossRef]
- Subbarao, G.V.; Yoshihashi, T.; Worthington, M.; Nakahara, K.; Ando, Y.; Sahrawat, K.L.; Rao, I.M.; Lata, J.-C.; Kishii, M.; Braun, H.-J. Suppression of soil nitrification by plants. Plant Sci. 2015, 233, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Dear, B.; Moore, G.; Hughes, S. Adaptation and potential contribution of temperate perennial legumes to the southern Australian wheatbelt: A review. Aust. J. Exp. Agric. 2003, 43, 1–18. [Google Scholar] [CrossRef]
- ABMI_Website, 3.8.2021. Alberta Biodiversity Monitoring Institute: Alsike Clover (Trifolium hybridum). Available online: https://abmi.ca/home/data-analytics/biobrowser-home/species-profile?tsn=99003745 (accessed on 29 October 2021).
- Rogers, M.E.; Colmer, T.D.; Frost, K.; Henry, D.; Cornwall, D.; Hulm, E.; Deretic, J.; Hughes, S.R.; Craig, A.D. Diversity in the genus Melilotus for tolerance to salinity and waterlogging. Plant Soil 2008, 304, 89–101. [Google Scholar] [CrossRef]
- ABMI_Website, 12.8.2021. Alberta Biodiversity Monitoring Institute: White Clover (Trifolium repens). Available online: https://abmi.ca/home/data-analytics/biobrowser-home/species-profile?year=2016&tsn=99003747 (accessed on 29 October 2021).
- IAEA_TECDOC. Improving Yield and Nitrogen Fixation of Grain Legumes in the Tropics and Sub-tropics of Asia; No. 1027; International Atomic Energy Agency: Vienna, Austria, 1998. [Google Scholar]
- Lynch, J.M.; Whipps, J.M. Substrate flow in the rhizosphere. Plant Soil 1990, 129, 1–10. [Google Scholar] [CrossRef]
- Roque-Malo, S.; Woo, D.K.; Kumar, P. Modeling the Role of Root Exudation in Critical Zone Nutrient Dynamics. Water Resour. Res. 2020, 56, 23. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Blesh, J. Functional traits in cover crop mixtures: Biological nitrogen fixation and multifunctionality. J. Appl. Ecol. 2018, 55, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Wang, E.; Xing, M.; Zhao, W.; Chen, X. Rhizosphere and non-rhizosphere bacterial community composition of the wild medicinal plant Rumex patientia. World J. Microbiol. Biotechnol. 2012, 28, 2257–2265. [Google Scholar] [CrossRef]
- Schlosse, G. On influence of external conditions on content of coumarin and related compounds in leaves of Melilotus albus. Z. Pflanzenzucht. 1968, 59, 327. [Google Scholar]
- Sugiyama, A.; Shitan, N.; Yazaki, K. Involvement of a Soybean ATP-Binding Cassette-Type Transporter in the Secretion of Genistein, a Signal Flavonoid in Legume-Rhizobium Symbiosis. Plant Physiol. 2007, 144, 2000–2008. [Google Scholar] [CrossRef] [Green Version]
- Barazani, O.; Friedman, J. Allelopathic bacteria and their impact on higher plants. Crit. Rev. Microbiol. 2001, 27, 41–55. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research Progress on the use of Plant Allelopathy in Agriculture and the Physiological and Ecological Mechanisms of Allelopathy. Front Plant Sci 2015, 6, 1020. [Google Scholar] [CrossRef]
- Li, Y.-P.; Feng, Y.-L.; Kang, Z.-L.; Zheng, Y.-L.; Zhang, J.-L.; Chen, Y.-J. Changes in soil microbial communities due to biological invasions can reduce allelopathic effects. J. Appl. Ecol. 2017, 54, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ge, X.; Fan, X.; Liu, H.; Gao, Y.; Ren, A. The Inhibitory Effect of Endophyte-Infected Tall Fescue on White Clover Can Be Alleviated by Glomus mosseae Instead of Rhizobia. Microorganisms 2021, 9, 109. [Google Scholar] [CrossRef]
- Korir, H.; Mungai, N.W.; Thuita, M.; Hamba, Y.; Masso, C. Co-inoculation Effect of Rhizobia and Plant Growth Promoting Rhizobacteria on Common Bean Growth in a Low Phosphorus Soil. Front. Plant Sci 2017, 8, 141. [Google Scholar] [CrossRef] [Green Version]
- Hack, C.M.; Porta, M.; Schäufele, R.; Grimoldi, A.A. Arbuscular mycorrhiza mediated effects on growth, mineral nutrition and biological nitrogen fixation of Melilotus alba Med. in a subtropical grassland soil. Appl. Soil Ecol. 2019, 134, 38–44. [Google Scholar] [CrossRef]
- Orhan, E.; Esitken, A.; Ercisli, S.; Turan, M.; Sahin, F. Effects of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient contents in organically growing raspberry. Sci. Hortic. 2006, 111, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ.-Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Keiluweit, M.; Wanzek, T.; Kleber, M.; Nico, P.; Fendorf, S. Anaerobic microsites have an unaccounted role in soil carbon stabilization. Nat. Commun. 2017, 8, 1771. [Google Scholar] [CrossRef]
- Freeman, C.; Ostle, N.J.; Fenner, N.; Kang, H. A regulatory role for phenol oxidase during decomposition in peatlands. Soil Biol. Biochem. 2004, 36, 1663–1667. [Google Scholar] [CrossRef]
- Brennan, E.B.; Acosta-Martinez, V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Ranells, N.N.; Wagger, M.G. Nitrogen-15 recovery and release by rye and crimson clover cover crops. Soil Sci. Soc. Am. J. 1997, 61, 943–948. [Google Scholar] [CrossRef]
- Tian, L.; Shi, W. Short-term effects of plant litter on the dynamics, amount, and stoichiometry of soil enzyme activity in agroecosystems. Eur. J. Soil Biol. 2014, 65, 23–29. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, W.; Liu, G.; He, P.; Liu, Z. Soil C/N and pH together as a comprehensive indicator for evaluating the effects of organic substitution management in subtropical paddy fields after application of high-quality amendments. Geoderma 2019, 337, 1116–1125. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, R.; Zhang, H.; Christie, P.; Feng, G.; Li, X.; Gai, J. Stoichiometric analysis of an arable crop-soil-microbe system after repeated fertilizer and compost application for 10 years. J. Soils Sediments 2021, 21, 1466–1475. [Google Scholar] [CrossRef]
- Li, Z.; Zu, C.; Wang, C.; Yang, J.; Yu, H.; Wu, H. Different responses of rhizosphere and non-rhizosphere soil microbial communities to consecutive Piper nigrum L. monoculture. Sci. Rep. 2016, 6, 35825. [Google Scholar] [CrossRef]
- Herman, D.J.; Johnson, K.K.; Jaeger, C.H.; Schwartz, E.; Firestone, M.K. Root Influence on Nitrogen Mineralization and Nitrification in Avena barbata Rhizosphere Soil. Soil Sci. Soc. Am. J. 2006, 70, 1504–1511. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, R.J.; Stevens, R.J. Evidence for Fungal Dominance of Denitrification and Codenitrification in a Grassland Soil. Soil Sci. Soc. Am. J. 2002, 66, 1540–1548. [Google Scholar] [CrossRef]
- Bedmar, E.; Bueno., E.; Correa., D.; Torres, M.; Delgado, M.; Mesa, S. Ecology of Denitrification in Soils and Plant-Associated Bacteria. In Benef. Plant-Microb. Interact.; Gonzalez, M.B.R., Gonzalez-Lopez, J., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 165–182. [Google Scholar]
- Li, X.; Petersen, S.O.; Sørensen, P.; Olesen, J.E. Effects of contrasting catch crops on nitrogen availability and nitrous oxide emissions in an organic cropping system. Agric. Ecosyst. Environ. 2015, 199, 382–393. [Google Scholar] [CrossRef]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkacz, A.; Cheema, J.; Chandra, G.; Grant, A.; Poole, P.S. Stability and succession of the rhizosphere microbiota depends upon plant type and soil composition. ISME J. 2015, 9, 2349–2359. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.X.; Kuzyakov, Y. Root effects on soil organic matter decomposition. In Roots and Soil Management: Interactions between Roots and the Soil; Zobel, R.W., Wright, S.F., Eds.; Agronomy Monograph No. 48; ASA-CSSA-SSSA: Madison, WI, USA, 2005; pp. 119–143. [Google Scholar]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Blagodatskaya, E.; Kuzyakov, Y. Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol. Fertil. Soils 2008, 45, 115–131. [Google Scholar] [CrossRef]
- Mason-Jones, K.; Kuzyakov, Y. Soil Biology & Biochemistry “Non-metabolizable” glucose analogue shines new light on priming mechanisms: Triggering of microbial metabolism. Soil Biol. Biochem. 2017, 107, 68–76. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (lat.) | Species | Variety | Sowing [g·m−2] |
|---|---|---|---|
| Trifolium incarnatum L. | crimson clover | Kardinál | 2.74 |
| Trifolium repens L. | white clover | Rivendel (small-leafed) | 0.60 |
| Trifolium hybridum L. | alsike clover | Táborský | 1.78 |
| Melilotus albus Medik. | sweet clover | Běla (2-year) | 1.71 |
| Lotus corniculatus L. | bird’s-foot trefoil | Lotar | 1.60 |
| Number | Samples | Species | Description |
|---|---|---|---|
| I. | crimson clover bulk | Trifolium incarnatum | soil >2 mm from the root |
| II. | crimson clover rhizosoil | Trifolium incarnatum | soil ≤2 mm close the root |
| III. | white clover bulk | Trifolium repens | soil >2 mm from the root |
| IV. | white clover rhizosoil | Trifolium repens | soil ≤2 mm close the root |
| V. | alsike clover bulk | Trifolium hybridum | soil >2 mm from the root |
| VI. | alsike clover rhizosoil | Trifolium hybridum | soil ≤2 mm close the root |
| VII. | sweet clover bulk | Melilotus albus | soil >2 mm from the root |
| VIII. | sweet clover rhizosoil | Melilotus albus | soil ≤2 mm close the root |
| IX. | bird’s-foot trefoil bulk | Lotus corniculatus | soil >2 mm from the root |
| X. | bird’s-foot trefoil rhizosoil | Lotus corniculatus | soil ≤2 mm close the root |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brtnicky, M.; Kintl, A.; Hammerschmiedt, T.; Mustafa, A.; Elbl, J.; Kucerik, J.; Vyhnanek, T.; Skladanka, J.; Hunady, I.; Holatko, J. Clover Species Specific Influence on Microbial Abundance and Associated Enzyme Activities in Rhizosphere and Non-Rhizosphere Soils. Agronomy 2021, 11, 2214. https://doi.org/10.3390/agronomy11112214
Brtnicky M, Kintl A, Hammerschmiedt T, Mustafa A, Elbl J, Kucerik J, Vyhnanek T, Skladanka J, Hunady I, Holatko J. Clover Species Specific Influence on Microbial Abundance and Associated Enzyme Activities in Rhizosphere and Non-Rhizosphere Soils. Agronomy. 2021; 11(11):2214. https://doi.org/10.3390/agronomy11112214
Chicago/Turabian StyleBrtnicky, Martin, Antonin Kintl, Tereza Hammerschmiedt, Adnan Mustafa, Jakub Elbl, Jiri Kucerik, Tomas Vyhnanek, Jiri Skladanka, Igor Hunady, and Jiri Holatko. 2021. "Clover Species Specific Influence on Microbial Abundance and Associated Enzyme Activities in Rhizosphere and Non-Rhizosphere Soils" Agronomy 11, no. 11: 2214. https://doi.org/10.3390/agronomy11112214