
Hypothermic Storage of 3D Cultured Multipotent Mesenchymal Stromal Cells for Regenerative Medicine Applications


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection and Source of Chemicals
2.2. Cell Culture
2.3. Scaffolds, Cell Seeding, and Culture
2.4. Hypothermic Storage of 3D Constructs
2.5. Resazurin Assay
2.6. Immunofluorescent Staining
2.7. Total RNA Isolation
2.8. Real-Time PCR
2.9. Conditioned Medium (CM) Production and Proteomic Analysis
2.10. Statistical Analysis
3. Results
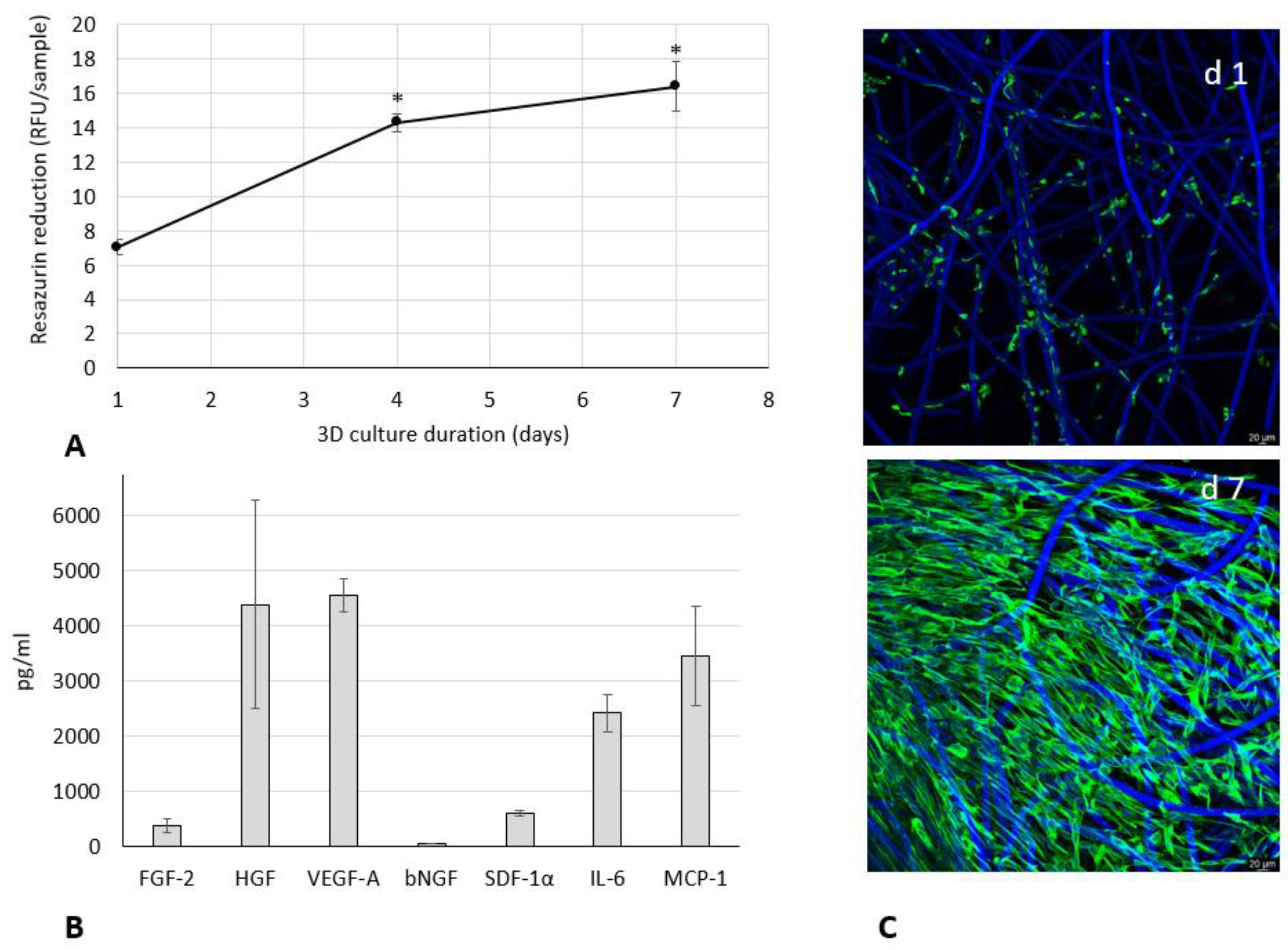
3.1. Growth and Paracrine Activity of WJ-MSCs within 3D HyaloFast®
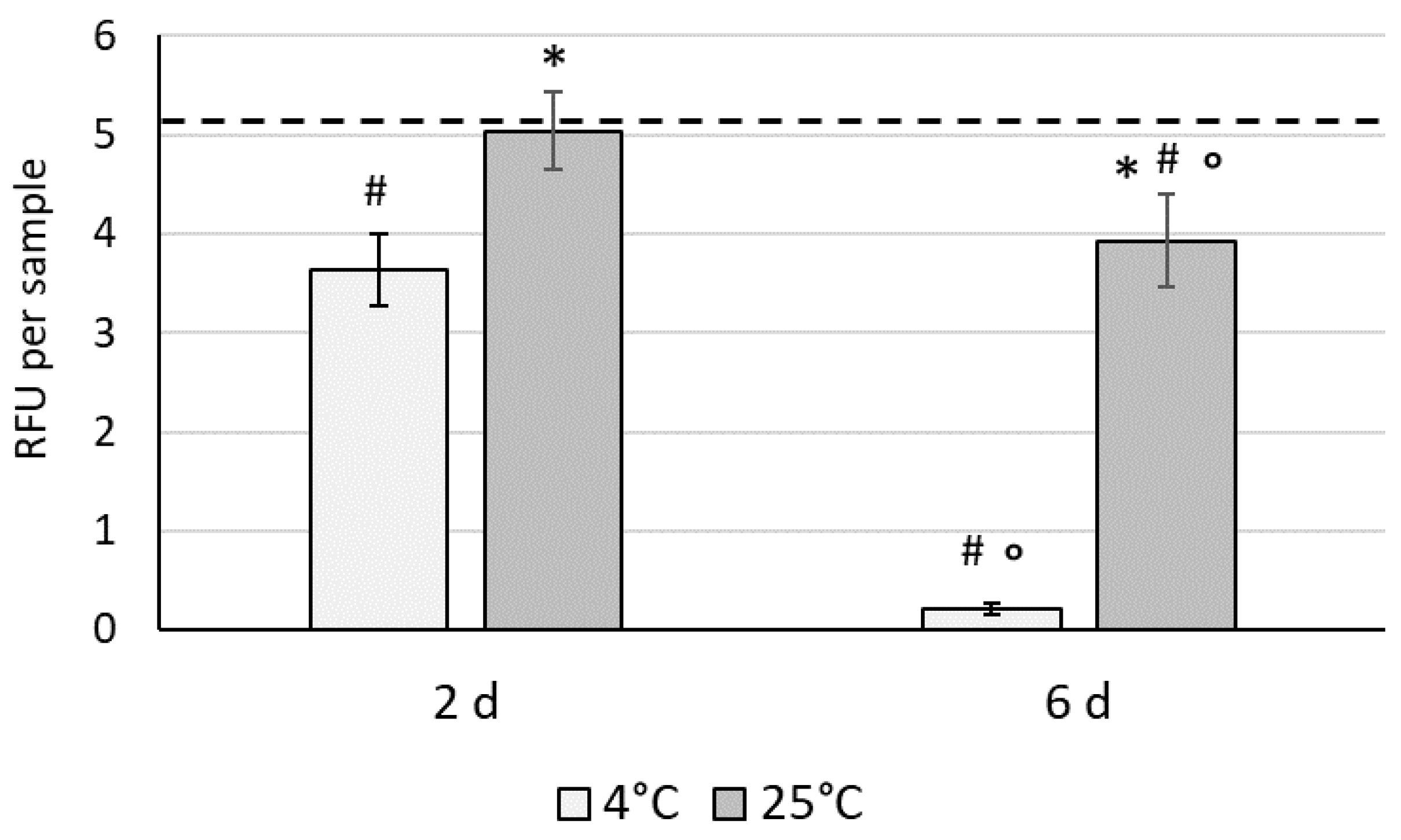
3.2. The Effect of Hypothermic Storage on the Recovery of WJ-MSCs in 3D Conditions
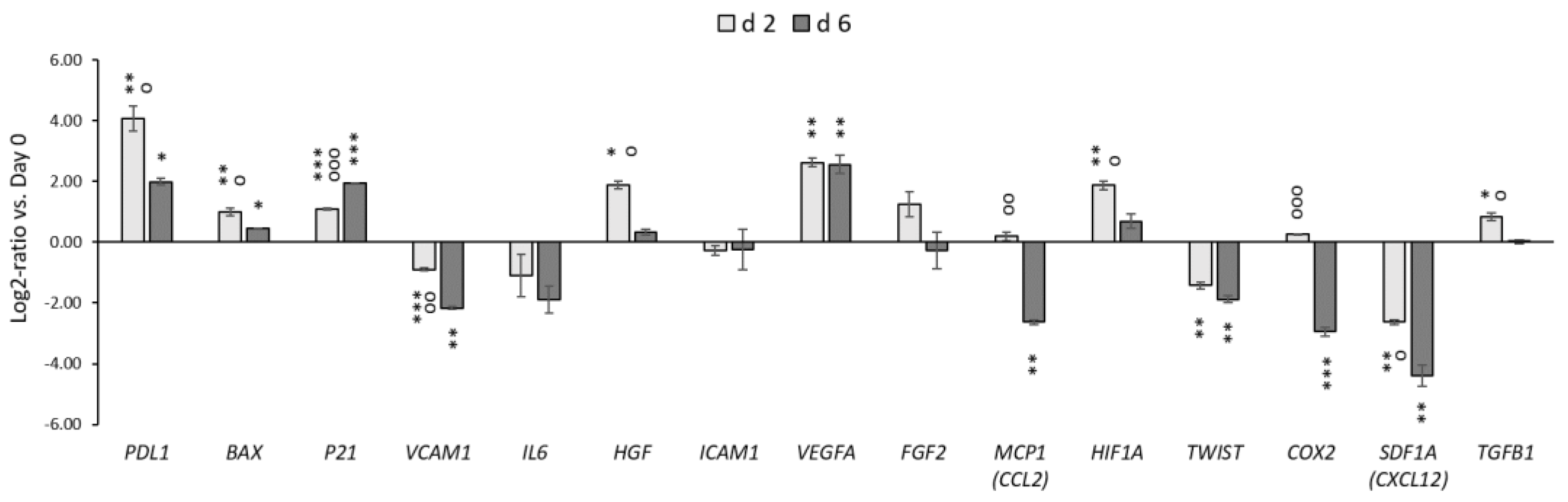
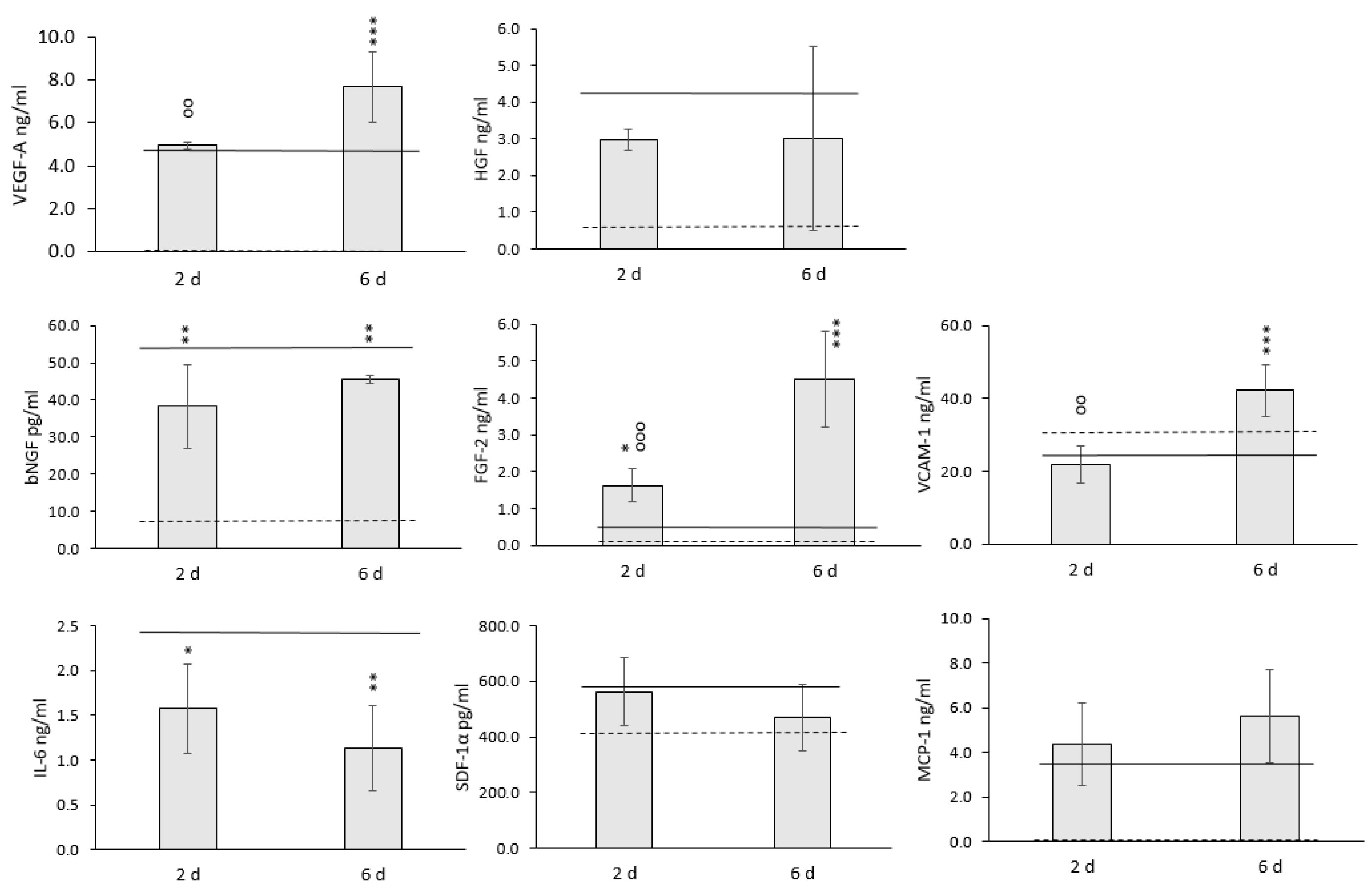
3.3. The Effect of Mild Hypothermic Storage under Ambient (25 °C) Conditions on the Gene Expression and Paracrine Secretion Profile of WJ-MSCs within 3D HyaloFast®
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise review: Mesenchymal stem cells: From roots to boost. Stem Cells 2019, 37, 855–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yamamoto, Y.; Xiao, Z.; Ochiya, T. The Immunomodulatory Functions of Mesenchymal Stromal/Stem Cells Mediated via Paracrine Activity. J. Clin. Med. 2019, 8, 1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrenko, Y.; Vackova, I.; Kekulova, K.; Chudickova, M.; Koci, Z.; Turnovcova, K.; Skalnikova, H.K.; Vodicka, P.; Kubinova, S. A Comparative Analysis of Multipotent Mesenchymal Stromal Cells derived from Different Sources, with a Focus on Neuroregenerative Potential. Sci. Rep. 2020, 10, 4290. [Google Scholar] [CrossRef] [PubMed]
- Kalaszczynska, I.; Ferdyn, K. Wharton’s Jelly Derived Mesenchymal Stem Cells: Future of Regenerative Medicine? Recent Findings and Clinical Significance. BioMed Res. Int. 2015, 2015, 1–11. [Google Scholar] [CrossRef]
- Wong, S.W.; Lenzini, S.; Giovanni, R.; Knowles, K.; Shin, J.-W. Matrix biophysical cues direct mesenchymal stromal cell functions in immunity. Acta Biomater. 2021, 133, 126–138. [Google Scholar] [CrossRef]
- Follin, B.; Juhl, M.; Cohen, S.; Perdersen, A.E.; Kastrup, J.; Ekblond, A. Increased Paracrine Immunomodulatory Potential of Mesenchymal Stromal Cells in Three-Dimensional Culture. Tissue Eng. Part B Rev. 2016, 22, 1–26. [Google Scholar] [CrossRef]
- Petrenko, Y.; Syková, E.; Kubinová, Š. The therapeutic potential of three-dimensional multipotent mesenchymal stromal cell spheroids. Stem Cell Res. Ther. 2017, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- De Cássia Noronha, N.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Park, I.S.; Rhie, J.-W.; Kim, S.-H. A novel three-dimensional adipose-derived stem cell cluster for vascular regeneration in ischemic tissue. Cytotherapy 2014, 16, 508–522. [Google Scholar] [CrossRef]
- Bhang, S.H.; Lee, S.; Shin, J.-Y.; Lee, T.-J.; Kim, B.-S. Transplantation of Cord Blood Mesenchymal Stem Cells as Spheroids Enhances Vascularization. Tissue Eng. Part A 2012, 18, 2138–2147. [Google Scholar] [CrossRef] [PubMed]
- Drzeniek, N.M.; Mazzocchi, A.; Schlickeiser, S.; Forsythe, S.D.; Moll, G.; Geissler, S.; Reinke, P.; Gossen, M.; Gorantla, V.S.; Volk, H.-D.; et al. Bio-instructive hydrogel expands the paracrine potency of mesenchymal stem cells. Biofabrication 2021, 13, 045002. [Google Scholar] [CrossRef] [PubMed]
- Qazi, T.H.; Mooney, D.; Duda, G.N.; Geissler, S. Niche-mimicking interactions in peptide-functionalized 3D hydrogels amplify mesenchymal stromal cell paracrine effects. Biomaterials 2020, 230, 119639. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.H.G.; McDevitt, T.C.; Cabral, J.M.; da Silva, C.L.; Ferreira, F.C. Tridimensional configurations of human mesenchymal stem/stromal cells to enhance cell paracrine potential towards wound healing processes. J. Biotechnol. 2017, 262, 28–39. [Google Scholar] [CrossRef]
- Vindigni, V.; Cortivo, R.; Iacobellis, L.; Abatangelo, G.; Zavan, B. Hyaluronan Benzyl Ester as a Scaffold for Tissue Engineering. Int. J. Mol. Sci. 2009, 10, 2972–2985. [Google Scholar] [CrossRef] [Green Version]
- Tsanaktsidou, E.; Kammona, O.; Kiparissides, C. Recent Developments in Hyaluronic Acid-Based Hydrogels for Cartilage Tissue Engineering Applications. Polymers 2022, 14, 839. [Google Scholar] [CrossRef]
- Abbas, M.; Alkaff, M.; Jilani, A.; Alsehli, H.; Damiati, L.; Kotb, M.; Abdelwahed, M.; Alghamdi, F.; Kalamegam, G. Combination of Mesenchymal Stem Cells, Cartilage Pellet and Bioscaffold Supported Cartilage Regeneration of a Full Thickness Articular Surface Defect in Rabbits. Tissue Eng. Regen. Med. 2018, 15, 661–671. [Google Scholar] [CrossRef]
- Gobbi, A.; Scotti, C.; Karnatzikos, G.; Mudhigere, A.; Castro, M.; Peretti, G. One-step surgery with multipotent stem cells and Hyaluronan-based scaffold for the treatment of full-thickness chondral defects of the knee in patients older than 45 years. Knee Surgery, Sports Traumatol. Arthrosc. 2017, 25, 2494–2501. [Google Scholar] [CrossRef] [Green Version]
- Gálvez, P.; Clares, B.; Bermejo, M.; Hmadcha, A.; Soria, B. Standard Requirement of a Microbiological Quality Control Program for the Manufacture of Human Mesenchymal Stem Cells for Clinical Use. Stem Cells Dev. 2014, 23, 1074–1083. [Google Scholar] [CrossRef] [Green Version]
- Coopman, K.; Medcalf, N. From production to patient: Challenges and approaches for delivering cell therapies. In StemBook; Harvard Stem Cell Institute Copyright: 2014 Karen Coopman and Nick Medcalf: Cambridge, MA, USA, 2008. Available online: https://www.ncbi.nlm.nih.gov/books/NBK208660/ (accessed on 25 May 2022).
- Günther, S.-K.; Geiss, C.; Kaiser, S.J.; Mutters, N.T.; Günther, F. Microbiological Control of Cellular Products: The Relevance of the Cellular Matrix, Incubation Temperature, and Atmosphere for the Detection Performance of Automated Culture Systems. Transfus. Med. Hemotherapy 2020, 47, 254–263. [Google Scholar] [CrossRef]
- Mutsenko, V.V.; Rogulska, O.Y.; Petrenko, Y.A.; Ehrlich, H.; Mazur, S.P.; Volkova, N.A.; Petrenko, A.Y. Cryosensitivity of Mesenchymal Stromal Cells Cryopreserved Within Marine Sponge Ianthella basta Skeleton-Based Carriers. Probl. Cryobiol. Cryomedicine 2016, 26, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Katsen-Globa, A.; Meiser, I.; Petrenko, Y.A.; Ivanov, R.V.; Lozinsky, V.I.; Zimmermann, H.; Petrenko, A. Towards ready-to-use 3-D scaffolds for regenerative medicine: Adhesion-based cryopreservation of human mesenchymal stem cells attached and spread within alginate-gelatin cryogel scaffolds. J. Mater. Sci. Mater. Med. 2014, 25, 857–871. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, Y.; Petrenko, A.; Martin, I.; Wendt, D. Perfusion bioreactor-based cryopreservation of 3D human mesenchymal stromal cell tissue grafts. Cryobiology 2017, 76, 150–153. [Google Scholar] [CrossRef] [PubMed]
- Arutyunyan, I.; Elchaninov, A.; Sukhikh, G.; Fatkhudinov, T. Cryopreservation of Tissue-Engineered Scaffold-Based Constructs: From Concept to Reality. Stem Cell Rev. Rep. 2022, 18, 1234–1252. [Google Scholar] [CrossRef] [PubMed]
- Koci, Z.; Výborný, K.; Dubišová, J.; Vacková, I.; Jäger, A.; Lunov, O.; Jiráková, K.; Kubinova, S. Extracellular Matrix Hydrogel Derived from Human Umbilical Cord as a Scaffold for Neural Tissue Repair and Its Comparison with Extracellular Matrix from Porcine Tissues. Tissue Eng. Part C Methods 2017, 23, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Rampersad, S.N. Multiple Applications of Alamar Blue as an Indicator of Metabolic Function and Cellular Health in Cell Viability Bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef] [PubMed]
- Bajuri, M.Y.; Sabri, S.; Mazli, N.; Sarifulnizam, F.A.; Apandi, H.M. Osteochondral Injury of the Talus Treated with Cell-Free Hyaluronic Acid-Based Scaffold (Hyalofast®)—A Reliable Solution. Cureus 2021, 13, e17928. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.; Pallamar, M.; Eder, T.; Ganger, R. Treatment of advanced stage osteochondrosis dissecans in the adolescent elbow using a hyaloronic acid-based scaffold: A case series of 5 patients. Arch. Orthop. Trauma. Surg. 2021, 141, 1541–1549. [Google Scholar] [CrossRef]
- Deszcz, I.; Lis-Nawara, A.; Grelewski, P.; Dragan, S.; Bar, J. Utility of direct 3D co-culture model for chondrogenic differentiation of mesenchymal stem cells on hyaluronan scaffold (Hyaff-11). Regen. Biomater. 2020, 7, 543–552. [Google Scholar] [CrossRef]
- Cristino, S.; Grassi, F.; Toneguzzi, S.; Piacentini, A.; Grigolo, B.; Santi, S.; Riccio, M.; Tognana, E.; Facchini, A.; Lisignoli, G. Analysis of mesenchymal stem cells grown on a three-dimensional HYAFF 11®-based prototype ligament scaffold. J. Biomed. Mater. Res. Part A 2005, 73, 275–283. [Google Scholar] [CrossRef]
- Pasquinelli, G.; Orrico, C.; Foroni, L.; Bonafè, F.; Carboni, M.; Guarnieri, C.; Raimondo, S.; Penna, C.; Geuna, S.; Pagliaro, P.; et al. Mesenchymal stem cell interaction with a non-woven hyaluronan-based scaffold suitable for tissue repair. J. Anat. 2008, 213, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Dennis, J.E.; Solchaga, L.A.; Awadallah, A.S.; Goldberg, V.M.; Caplan, A.I. Tissue-Engineered Fabrication of an Osteochondral Composite Graft Using Rat Bone Marrow-Derived Mesenchymal Stem Cells. Tissue Eng. 2001, 7, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, C.; Desando, G.; Columbaro, M.; Ferrari, A.; Zini, N.; Facchini, A.; Grigolo, B. Chondrogenic differentiation of bone marrow concentrate grown onto a hylauronan scaffold: Rationale for its use in the treatment of cartilage lesions. J. Biomed. Mater. Res. Part A 2013, 101, 1559–1570. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, C.; Desando, G.; Ferrari, A.; Zini, N.; Mariani, E.; Grigolo, B. Hyaluronan scaffold supports osteogenic differentiation of bone marrow concentrate cells. J. Biol. Regul. Homeost. Agents 2016, 30, 409–420. [Google Scholar]
- Cao, C.; Tarlé, S.; Kaigler, D. Characterization of the immunomodulatory properties of alveolar bone-derived mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Ortiz, A.D.C.; Fideles, S.O.M.; Pomini, K.T.; Bellini, M.Z.; Pereira, E.D.S.B.M.; Reis, C.H.B.; Pilon, J.P.G.; de Marchi, M.; Trazzi, B.F.d.M.; da Silva, W.S.; et al. Potential of Fibrin Glue and Mesenchymal Stem Cells (MSCs) to Regenerate Nerve Injuries: A Systematic Review. Cells 2022, 11, 221. [Google Scholar] [CrossRef]
- Ortiz, A.D.C.; Fideles, S.O.M.; Pomini, K.T.; Reis, C.H.B.; Bueno, C.R.D.S.; Pereira, E.D.S.B.M.; Rossi, J.D.O.; Novais, P.C.; Pilon, J.P.G.; Junior, G.M.R.; et al. Effects of Therapy with Fibrin Glue combined with Mesenchymal Stem Cells (MSCs) on Bone Regeneration: A Systematic Review. Cells 2021, 10, 2323. [Google Scholar] [CrossRef]
- Rogulska, O.; Tykhvynska, O.; Revenko, O.; Grischuk, V.; Mazur, S.; Volkova, N.; Vasyliev, R.; Petrenko, A.; Petrenko, Y.; Yuriy, P. Novel Cryopreservation Approach Providing Off-the-Shelf Availability of Human Multipotent Mesenchymal Stromal Cells for Clinical Applications. Stem Cells Int. 2019, 2019, 11. [Google Scholar] [CrossRef]
- Fuller, B.J.; Petrenko, A.; Rodriguez, J.V.; Somov, A.Y.; Balaban, C.L.; Guibert, E.E. Biopreservation of hepatocytes: Current concepts on hypothermic preservation, cryopreservation, and vitrification. Cryo Lett. 2013, 34, 432–452. [Google Scholar]
- Guibert, E.E.; Petrenko, A.Y.; Balaban, C.L.; Somov, A.Y.; Rodriguez, J.V.; Fuller, B.J. Organ Preservation: Current Concepts and New Strategies for the Next Decade. Transfus. Med. Hemotherapy 2011, 38, 125–142. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, Y.; Chudickova, M.; Vackova, I.; Groh, T.; Kosnarova, E.; Cejkova, J.; Turnovcova, K.; Petrenko, A.; Sykova, E.; Kubinova, S. Clinically Relevant Solution for the Hypothermic Storage and Transportation of Human Multipotent Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019, 5909524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarusin, D.N.; Petrenko, Y.A.; Semenchenko, O.A.; Mutsenko, V.V.; Zaikov, V.S.; Petrenko, A.Y. Efficiency of the sucrose-based solution and UW solution for hypothermic storage of human mesenchymal stromal cells in suspension or within alginate microspheres. Probl. Cryobiol. Cryomedicine 2015, 25, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Tam, E.; McGrath, M.; Sladkova, M.; AlManaie, A.; Alostaad, A.; De Peppo, G.M. Hypothermic and cryogenic preservation of tissue-engineered human bone. Ann. N. Y. Acad. Sci. 2020, 1460, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, G.; Zhou, G.; Cen, L.; Cui, L.; Cao, Y. Comparative investigation of viability, metabolism and osteogenic capability of tissue-engineered bone preserved in sealed osteogenic media at 37 °C and 4 °C. Biomed. Mater. 2010, 5, 35010. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, G. Hypothermic Storage of Human Umbilical Cord Mesenchymal Stem Cells and Their Hydrogel Constructs. Cryo Lett. 2020, 41, 100–105. [Google Scholar]
- Kubrova, E.; Qu, W.; Galvan, M.L.; Paradise, C.R.; Yang, J.; Dietz, A.B.; Dudakovic, A.; Smith, J.; van Wijnen, A.J. Hypothermia and nutrient deprivation alter viability of human adipose-derived mesenchymal stem cells. Gene 2019, 722, 144058. [Google Scholar] [CrossRef]
- Johnson, M.R.; Valentine, C.; Basilico, C.; Mansukhani, A. FGF signaling activates STAT1 and p21 and inhibits the estrogen response and proliferation of MCF-7 cells. Oncogene 1998, 16, 2647–2656. [Google Scholar] [CrossRef] [Green Version]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair 2016, 42, 63–71. [Google Scholar] [CrossRef]
- Aizman, I.; Vinodkumar, D.; McGrogan, M.; Bates, D. Cell Injury-Induced Release of Fibroblast Growth Factor 2: Relevance to Intracerebral Mesenchymal Stromal Cell Transplantations. Stem Cells Dev. 2015, 24, 1623–1634. [Google Scholar] [CrossRef] [Green Version]
- Bock, F.J.; Sedov, E.; Koren, E.; Koessinger, A.L.; Cloix, C.; Zerbst, D.; Athineos, D.; Anand, J.; Campbell, K.J.; Blyth, K.; et al. Apoptotic stress-induced FGF signalling promotes non-cell autonomous resistance to cell death. Nat. Commun. 2021, 12, 6572. [Google Scholar] [CrossRef]
- Pang, S.H.M.; D’Rozario, J.; Mendonca, S.; Bhuvan, T.; Payne, N.L.; Zheng, D.; Hisana, A.; Wallis, G.; Barugahare, A.; Powell, D.; et al. Mesenchymal stromal cell apoptosis is required for their therapeutic function. Nat. Commun. 2021, 12, 6495. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; English, K.; Krasnodembskaya, A.; Isaza-Correa, J.M.; Hawthorne, I.J.; Mahon, B.P. The Necrobiology of Mesenchymal Stromal Cells Affects Therapeutic Efficacy. Front. Immunol. 2019, 10, 1228. [Google Scholar] [CrossRef] [PubMed]
- Zimna, A.; Kurpisz, M. Hypoxia-Inducible Factor-1 in Physiological and Pathophysiological Angiogenesis: Applications and Therapies. BioMed Res. Int. 2015, 2015, 549412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.-C.; Wang, L.-L.; Zhang, X.-D.; Xu, J.-L.; Li, P.-F.; Liang, H.; Zhang, X.-B.; Xie, L.; Zhou, Z.-H.; Yang, J.; et al. The relationship between expression of PD-L1 and HIF-1α in glioma cells under hypoxia. J. Hematol. Oncol. 2021, 14, 92. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Full Gene Name |
|---|---|
| Chemokines: | |
| CXCL12/SDF1a | C-X-C Motif Chemokine Ligand 12/SDF1AStromal Cell-Derived Factor 1 |
| CCL2/MCP1 | C-C Motif Chemokine Ligand 2/MCP1Monocyte Chemoattractant Protein 1 |
| Adhesion molecules: | |
| ICAM1 | Intercellular Adhesion Molecule 1 |
| VCAM1 | Vascular cell adhesion protein |
| Transcription factors: | |
| TWIST | Twist Family BHLH Transcription Factor |
| IL6 | Interleukin 6 |
| Immunoregulatory/inflammatory/anti-inflammatory genes: | |
| PDL1 | Programmed death-ligand 1 |
| COX2 | Cyclooxygenase 2 |
| TGFB1 | Transforming Growth Factor Beta 1 |
| IL6 | Interleukin 6 |
| Angiogenic/neurotrophic factors: | |
| VEGFA | Vascular Endothelial Growth Factor A |
| HGF | Hepatocyte Growth Factor |
| FGF2 | Fibroblast Growth Factor 2 |
| Genes associated with cell cycle, metabolism, and apoptotic processes: | |
| P21 | Cyclin-Dependent Kinase Inhibitor 1 |
| HIF1A | Hypoxia Inducible Factor 1 Subunit Alpha |
| BAX | BCL2 Associated X Protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vackova, I.; Vavrinova, E.; Musilkova, J.; Havlas, V.; Petrenko, Y. Hypothermic Storage of 3D Cultured Multipotent Mesenchymal Stromal Cells for Regenerative Medicine Applications. Polymers 2022, 14, 2553. https://doi.org/10.3390/polym14132553
Vackova I, Vavrinova E, Musilkova J, Havlas V, Petrenko Y. Hypothermic Storage of 3D Cultured Multipotent Mesenchymal Stromal Cells for Regenerative Medicine Applications. Polymers. 2022; 14(13):2553. https://doi.org/10.3390/polym14132553
Chicago/Turabian StyleVackova, Irena, Eliska Vavrinova, Jana Musilkova, Vojtech Havlas, and Yuriy Petrenko. 2022. "Hypothermic Storage of 3D Cultured Multipotent Mesenchymal Stromal Cells for Regenerative Medicine Applications" Polymers 14, no. 13: 2553. https://doi.org/10.3390/polym14132553