Cyanobacterial Harmful Bloom Lipopolysaccharides Induce Pro-Inflammatory Effects in Immune and Intestinal Epithelial Cells In Vitro

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. LPS Samples Characterization and Their Endotoxin Activity
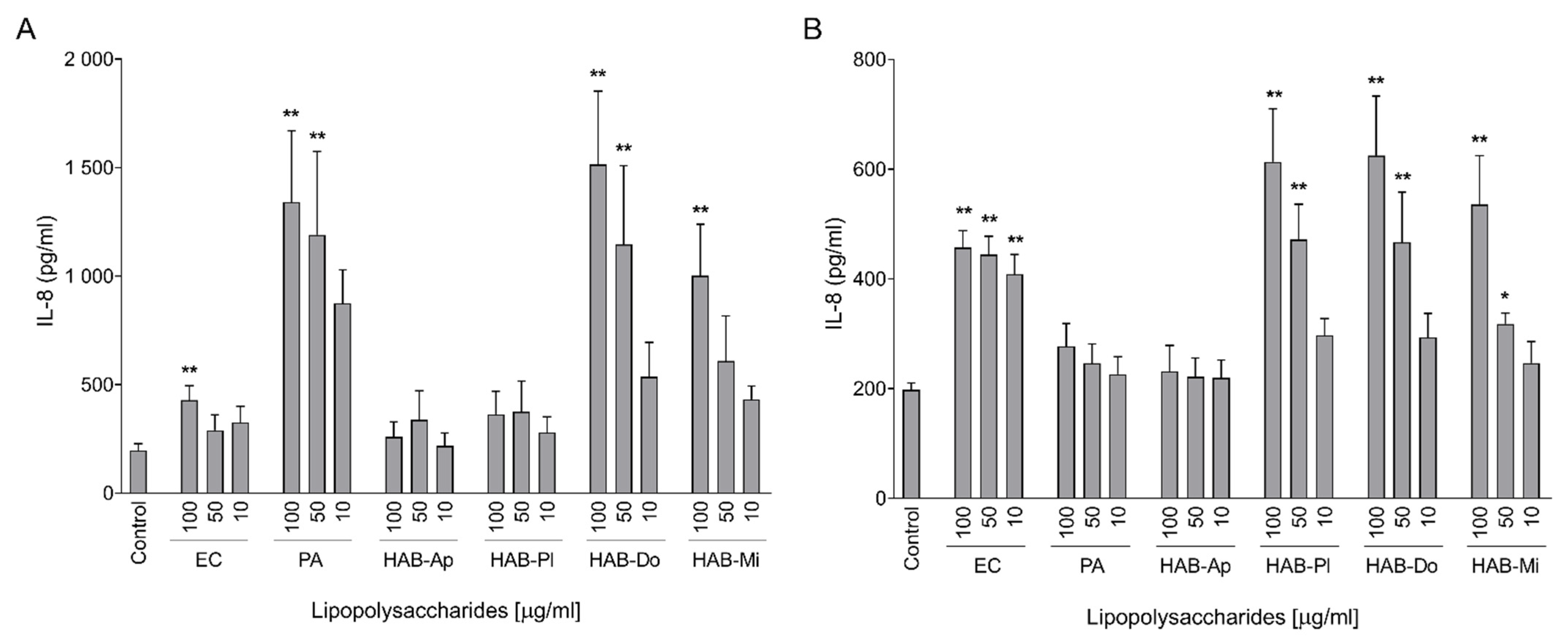
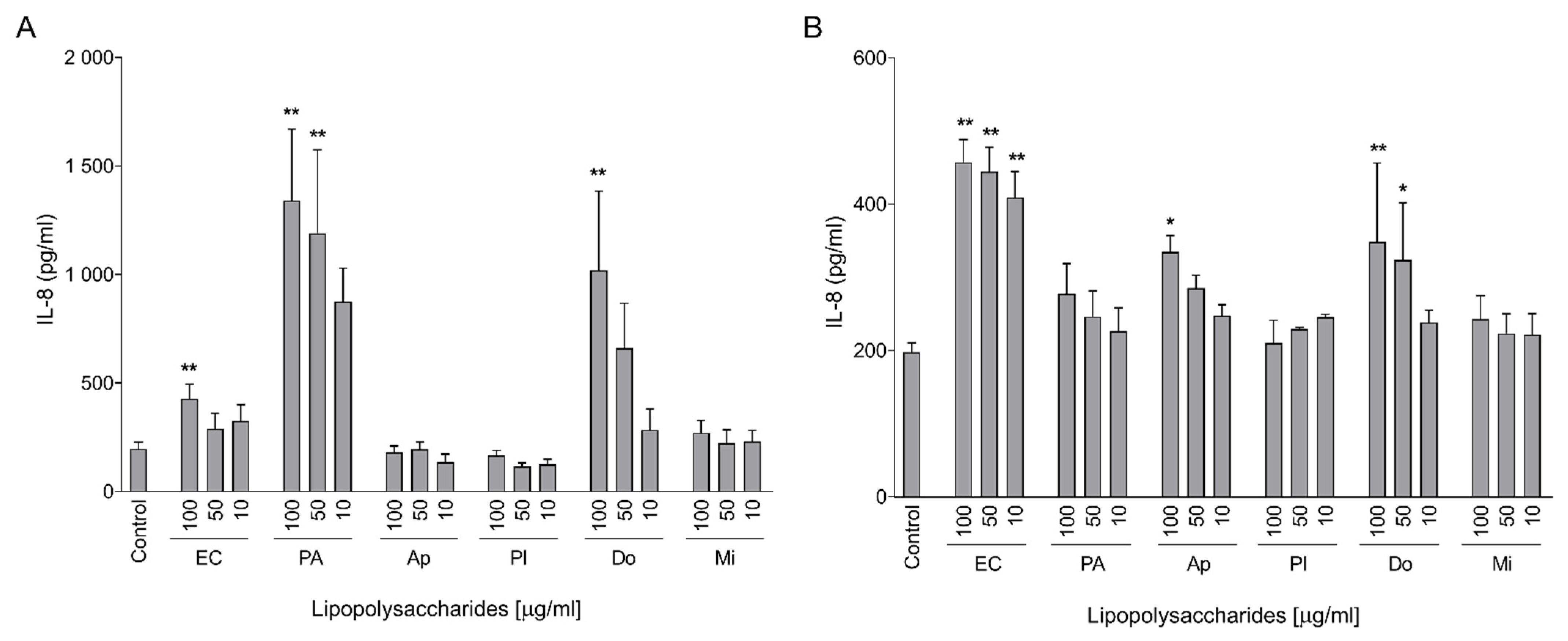
2.2. Pro-Inflammatory Activation of Intestinal Cells
2.3. Pro-Inflammatory Effects on Macrophages
2.4. Biomass DNA Composition
2.5. SDS-PAGE Migration Patterns of Studied LPS
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cyanobacterial Biomass Preparation
5.2. LPS Preparation
5.3. Characterization of LPSs by SDS-PAGE
5.4. RT-qPCR Detection of Cyanobacteria, Heterotrophic Bacteria, and Gram-Negative Heterotrophic Bacteria
5.5. PyroGeneTM rFC Assay
5.6. Cell Cultures
5.7. Lactate Dehydrogenase Assay
5.8. Specific Cytokine Detection
5.9. Cytokine Array
5.10. NO Production
5.11. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blahova, L.; Adamovsky, O.; Kubala, L.; Svihalkova Sindlerova, L.; Zounkova, R.; Blaha, L. The isolation and characterization of lipopolysaccharides from Microcystis aeruginosa, a prominent toxic water bloom forming cyanobacteria. Toxicon Off. J. Int. Soc. Toxinology 2013, 76, 187–196. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31, 31. [Google Scholar] [CrossRef] [Green Version]
- Novakova, K.; Babica, P.; Adamovsky, O.; Blaha, L. Modulation of gap-junctional intercellular communication by a series of cyanobacterial samples from nature and laboratory cultures. Toxicon Off. J. Int. Soc. Toxinology 2011, 58, 76–84. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Vasicek, O.; Hajek, J.; Blahova, L.; Hrouzek, P.; Babica, P.; Kubala, L.; Sindlerova, L. Cyanobacterial lipopeptides puwainaphycins and minutissamides induce disruptive and pro-inflammatory processes in Caco-2 human intestinal barrier model. Harmful Algae 2020, 96, 101849. [Google Scholar] [CrossRef]
- Durai, P.; Batool, M.; Choi, S. Structure and Effects of Cyanobacterial Lipopolysaccharides. Mar. Drugs 2015, 13, 4217–4230. [Google Scholar] [CrossRef] [Green Version]
- Sulc, R.; Szekely, G.; Shinde, S.; Wierzbicka, C.; Vilela, F.; Bauer, D.; Sellergren, B. Phospholipid imprinted polymers as selective endotoxin scavengers. Sci. Rep. 2017, 7, 44299. [Google Scholar] [CrossRef] [Green Version]
- Mankiewicz-Boczek, J.; Font-Najera, A. Temporal and functional interrelationships between bacterioplankton communities and the development of a toxigenic Microcystis bloom in a lowland European reservoir. Sci. Rep. 2022, 12, 19332. [Google Scholar] [CrossRef]
- Perez-Carrascal, O.M.; Tromas, N.; Terrat, Y.; Moreno, E.; Giani, A.; Correa Braga Marques, L.; Fortin, N.; Shapiro, B.J. Single-colony sequencing reveals microbe-by-microbiome phylosymbiosis between the cyanobacterium Microcystis and its associated bacteria. Microbiome 2021, 9, 194. [Google Scholar] [CrossRef]
- Tu, J.; Chen, L.; Gao, S.; Zhang, J.; Bi, C.; Tao, Y.; Lu, N.; Lu, Z. Obtaining Genome Sequences of Mutualistic Bacteria in Single Microcystis Colonies. Int. J. Mol. Sci. 2019, 20, 5047. [Google Scholar] [CrossRef] [Green Version]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caroff, M.; Novikov, A. Lipopolysaccharides: Structure, function and bacterial identifications. OCL 2020, 27, 31. [Google Scholar] [CrossRef]
- Gemma, S.; Molteni, M.; Rosseti, C. Lipopolysaccharides in Cyanobacteria: A Brief Overview. Adv. Microbiol. 2016, 6, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Welker, M. Cyanobacterial Lipopolysaccharides (LPS). In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: London, UK, 2021; pp. 137–148. [Google Scholar] [CrossRef]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, N.; Fernandez, R.C. Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front. Cell. Infect. Microbiol. 2013, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Allaire, J.M.; Crowley, S.M.; Law, H.T.; Chang, S.Y.; Ko, H.J.; Vallance, B.A. The Intestinal Epithelium: Central Coordinator of Mucosal Immunity. Trends Immunol. 2018, 39, 677–696. [Google Scholar] [CrossRef]
- Vasicek, O.; Lojek, A.; Ciz, M. Serotonin and its metabolites reduce oxidative stress in murine RAW264.7 macrophages and prevent inflammation. J. Physiol. Biochem. 2020, 76, 49–60. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Billings, W.H. Water-Associated Human Illness in Northeast Pennsylvania and its Suspected Association with Blue-Green Algae Blooms; Springer: Boston, MA, USA, 1981. [Google Scholar]
- Dillenberg, H.O.; Dehnel, M.K. Toxic waterbloom in Saskatchewan, 1959. Can. Med. Assoc. J. 1960, 83, 1151–1154. [Google Scholar]
- Rapala, J.; Robertson, A.; Negri, A.P.; Berg, K.A.; Tuomi, P.; Lyra, C.; Erkomaa, K.; Lahti, K.; Hoppu, K.; Lepisto, L. First report of saxitoxin in Finnish lakes and possible associated effects on human health. Environ. Toxicol. 2005, 20, 331–340. [Google Scholar] [CrossRef]
- Turner, P.C.; Gammie, A.J.; Hollinrake, K.; Codd, G.A. Pneumonia associated with contact with cyanobacteria. BMJ 1990, 300, 1440–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moosova, Z.; Sindlerova, L.; Ambruzova, B.; Ambrozova, G.; Vasicek, O.; Velki, M.; Babica, P.; Kubala, L. Lipopolysaccharides from Microcystis Cyanobacteria-Dominated Water Bloom and from Laboratory Cultures Trigger Human Immune Innate Response. Toxins 2019, 11, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson-Mungerson, M.; Incrocci, R.; Subramaniam, V.; Williams, P.; Hall, M.L.; Mayer, A.M.S. Effects of cyanobacteria Oscillatoria sp. lipopolysaccharide on B cell activation and Toll-like receptor 4 signaling. Toxicol. Lett. 2017, 275, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Swanson-Mungerson, M.; Williams, P.; Gurr, J.R.; Incrocci, R.; Subramaniam, V.; Radowska, K.; Hall, M.L.; Mayer, A.M.S. Biochemical and Functional Analysis of Cyanobacterium Geitlerinema sp. LPS on Human Monocytes. Toxicol. Sci. 2019, 171, 421–430. [Google Scholar] [CrossRef]
- Labohá, P.; Sychrová, E.; Brózman, O.; Sovadinová, I.; Bláhová, L.; Prokeš, R.; Ondráček, J.; Babica, P. Cyanobacteria, Cyanotoxins and Lipopolysaccharides in Aerosols From Inland Freshwaters and Their Effects on Human Bronchial Cells. Environ. Toxicol. Pharmacol. 2023, 98, 104373. [Google Scholar] [CrossRef]
- Swartzendruber, J.A.; Del Toro, R.M.; Incrocci, R.; Seangmany, N.; Gurr, J.R.; Mayer, A.M.S.; Williams, P.G.; Swanson-Mungerson, M. Lipopolysaccharide from the Cyanobacterium Geitlerinema sp. Induces Neutrophil Infiltration and Lung Inflammation. Toxins 2022, 14, 267. [Google Scholar] [CrossRef]
- Pipal, M.; Priebojova, J.; Koci, T.; Blahova, L.; Smutna, M.; Hilscherova, K. Field cyanobacterial blooms producing retinoid compounds cause teratogenicity in zebrafish embryos. Chemosphere 2020, 241, 125061. [Google Scholar] [CrossRef]
- Javůrek, J. Biodetection Systems in the Assessment of Endocrine Disrupting Potential of Compounds in Surface Waters. Ph.D. Thesis, Masaryk University, Brno, Czech Republic, 2019. [Google Scholar]
- Szmucová, V. Instrumental and Biological In Vitro Methods for Analyses of Neurotoxic Cyanobacterial Metabolites. Master’s Thesis, Masaryk University, Brno, Czech Republic, 2019. [Google Scholar]
- Javurek, J.; Sychrova, E.; Smutna, M.; Bittner, M.; Kohoutek, J.; Adamovsky, O.; Novakova, K.; Smetanova, S.; Hilscherova, K. Retinoid compounds associated with water blooms dominated by Microcystis species. Harmful Algae 2015, 47, 116–125. [Google Scholar] [CrossRef]
- Sarkar, S.; Ulett, G.C.; Totsika, M.; Phan, M.D.; Schembri, M.A. Role of capsule and O antigen in the virulence of uropathogenic Escherichia coli. PLoS ONE 2014, 9, e94786. [Google Scholar] [CrossRef] [Green Version]
- Werts, C.; Tapping, R.I.; Mathison, J.C.; Chuang, T.H.; Kravchenko, V.; Saint Girons, I.; Haake, D.A.; Godowski, P.J.; Hayashi, F.; Ozinsky, A.; et al. Leptospiral lipopolysaccharide activates cells through a TLR2-dependent mechanism. Nat. Immunol. 2001, 2, 346–352. [Google Scholar] [CrossRef]
- Gupta, S.K.; Masinick, S.; Garrett, M.; Hazlett, L.D. Pseudomonas Aeruginosa Lipopolysaccharide Binds Galectin-3 and Other Human Corneal Epithelial Proteins. Infect. Immun. 1997, 65, 2747–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.; Murphy, K.; Lo, R.Y.; Khursigara, C.M.; Lam, J.S. Single-Nucleotide Polymorphisms Found in the MigA and WbpX Glycosyltransferase Genes Account for the Intrinsic Lipopolysaccharide Defects Exhibited by Pseudomonas Aeruginosa PA14. J. Bacteriol. 2015, 197, 2780–2791. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, C.; Midoux, C.; Vergnaud, G.; Latino, L. The Basis for Natural Multiresistance to Phage in Pseudomonas Aeruginosa. Antibiotics 2020, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jia, J.; Chen, S.; Huang, T.; Wang, Y.; Zhao, Z.; Feng, J.; Hao, H.; Li, S.; Ma, X. Dynamics of Bacterial and Fungal Communities during the Outbreak and Decline of an Algal Bloom in a Drinking Water Reservoir. Int. J. Environ. Res. Public Health 2018, 15, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Wenzel, C.Q.; Lam, J.S. Nonradiolabeling assay for WaaP, an essential sugar kinase involved in biosynthesis of core lipopolysaccharide of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2002, 46, 2035–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef] [Green Version]
- Bernardova, K.; Babica, P.; Marsalek, B.; Blaha, L. Isolation and endotoxin activities of lipopolysaccharides from cyanobacterial cultures and complex water blooms and comparison with the effects of heterotrophic bacteria and green alga. J. Appl. Toxicol. JAT 2008, 28, 72–77. [Google Scholar] [CrossRef]
- Smith, D.J.; Tan, J.Y.; Powers, M.A.; Lin, X.N.; Davis, T.W.; Dick, G.J. Individual Microcystis colonies harbour distinct bacterial communities that differ by Microcystis oligotype and with time. Environ. Microbiol. 2021, 23, 3020–3036. [Google Scholar] [CrossRef]
- Ohkouchi, Y.; Tajima, S.; Nomura, M.; Itoh, S. Inflammatory responses and potencies of various lipopolysaccharides from bacteria and cyanobacteria in aquatic environments and water supply systems. Toxicon Off. J. Int. Soc. Toxinology 2015, 97, 23–31. [Google Scholar] [CrossRef]
- Snyder, D.S.; Brahamsha, B.; Azadi, P.; Palenik, B. Structure of compositionally simple lipopolysaccharide from marine synechococcus. J. Bacteriol. 2009, 191, 5499–5509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, M.; Sato, Y.; Ito, H.; Masago, Y.; Omura, T. Monosaccharide composition of the outer membrane lipopolysaccharide and O-chain from the freshwater cyanobacterium Microcystis aeruginosa NIES-87. J. Appl. Microbiol. 2012, 113, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Haeffner-Cavaillon, N.; Carreno, M.P.; Aussel, L.; Caroff, M. Molecular aspects of endotoxins relevant to their biological functions. Nephrol. Dial. Transpl. 1999, 14, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucieklak, K.; Koj, S.; Niedziela, T. Bordetella holmesii Lipopolysaccharide Hide and Seek Game with Pertussis: Structural Analysis of the O-Specific Polysaccharide and the Core Oligosaccharide of the Type Strain ATCC 51541. Int. J. Mol. Sci. 2020, 21, 6433. [Google Scholar] [CrossRef]
- Poole, S.; Dawson, P.; Gaines Das, R.E. Second international standard for endotoxin: Calibration in an international collaborative study. J. Endotoxin Res. 1997, 4, 221–231. [Google Scholar] [CrossRef]
- Jurga, A.M.; Rojewska, E.; Makuch, W.; Mika, J. Lipopolysaccharide from Rhodobacter sphaeroides (TLR4 antagonist) attenuates hypersensitivity and modulates nociceptive factors. Pharm. Biol. 2018, 56, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Jiang, C.; Xu, S.; Zheng, X.; Lv, P.; Wang, C.; Wang, D.; Zhuang, X. Spatiotemporal changes of bacterial communities during a cyanobacterial bloom in a subtropical water source reservoir ecosystem in China. Sci. Rep. 2022, 12, 14573. [Google Scholar] [CrossRef]
- Camba-Gomez, M.; Arosa, L.; Gualillo, O.; Conde-Aranda, J. Chemokines and chemokine receptors in inflammatory bowel disease: Recent findings and future perspectives. Drug Discov. Today 2022, 27, 1167–1175. [Google Scholar] [CrossRef]
- Jia, S.N.; Han, Y.B.; Yang, R.; Yang, Z.C. Chemokines in colon cancer progression. Semin. Cancer Biol. 2022, 86, 400–407. [Google Scholar] [CrossRef]
- Mello, J.D.C.; Gomes, L.E.M.; Silva, J.F.; Siqueira, N.S.N.; Pascoal, L.B.; Martinez, C.A.R.; Ayrizono, M.L.S.; Leal, R.F. The role of chemokines and adipokines as biomarkers of Crohn’s disease activity: A systematic review of the literature. Am. J. Transl. Res. 2021, 13, 8561–8574. [Google Scholar]
- Zhu, Y.; Yang, S.; Zhao, N.; Liu, C.; Zhang, F.; Guo, Y.; Liu, H. CXCL8 chemokine in ulcerative colitis. Biomed. Pharmacother. 2021, 138, 111427. [Google Scholar] [CrossRef]
- Curciarello, R.; Canziani, K.E.; Docena, G.H.; Muglia, C.I. Contribution of Non-immune Cells to Activation and Modulation of the Intestinal Inflammation. Front. Immunol. 2019, 10, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kany, S.; Vollrath, J.T.; Relja, B. Cytokines in Inflammatory Disease. Int. J. Mol. Sci. 2019, 20, 6008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luissint, A.C.; Parkos, C.A.; Nusrat, A. Inflammation and the Intestinal Barrier: Leukocyte-Epithelial Cell Interactions, Cell Junction Remodeling, and Mucosal Repair. Gastroenterology 2016, 151, 616–632. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillebrand, H.; Durselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Skacelova, O.; Leps, J. The relationship of diversity and biomass in phytoplankton communities weakens when accounting for species proportions. Hydrobiologia 2014, 724, 67–77. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during Assembly of Head of Bacteriophage-T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Sehnal, L.; Smutna, M.; Blahova, L.; Babica, P.; Splichalova, P.; Hilscherova, K. The Origin of Teratogenic Retinoids in Cyanobacteria. Toxins 2022, 14, 636. [Google Scholar] [CrossRef]
- Lang-Yona, N.; Lehahn, Y.; Herut, B.; Burshtein, N.; Rudich, Y. Marine aerosol as a possible source for endotoxins in coastal areas. Sci. Total. Environ. 2014, 499, 311–318. [Google Scholar] [CrossRef]
- Bino, L.; Kucera, J.; Stefkova, K.; Svihalkova Sindlerova, L.; Lanova, M.; Kudova, J.; Kubala, L.; Pachernik, J. The stabilization of hypoxia inducible factor modulates differentiation status and inhibits the proliferation of mouse embryonic stem cells. Chem. Biol. Interact. 2016, 244, 204–214. [Google Scholar] [CrossRef]
- Moosova, Z.; Pekarova, M.; Sindlerova, L.S.; Vasicek, O.; Kubala, L.; Blaha, L.; Adamovsky, O. Immunomodulatory effects of cyanobacterial toxin cylindrospermopsin on innate immune cells. Chemosphere 2019, 226, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Pekarova, M.; Koudelka, A.; Kolarova, H.; Ambrozova, G.; Klinke, A.; Cerna, A.; Kadlec, J.; Trundova, M.; Sindlerova Svihalkova, L.; Kuchta, R.; et al. Asymmetric dimethyl arginine induces pulmonary vascular dysfunction via activation of signal transducer and activator of transcription 3 and stabilization of hypoxia-inducible factor 1-alpha. Vasc. Pharmacol. 2015, 73, 138–148. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Locality | Dominating Species (% Biovolume) | mg LPS/g d.w. (±S.E.) | PyroGeneTM (×103 EU/mg LPS) | PyroGeneTM (EU/mg d.w.) |
|---|---|---|---|---|---|
| Water blooms | |||||
| HAB-Ap | Ruda 25 July 2013 | Aphanizomenon klebahnii (92.4%) [29] | 15.88 (±4.61) | 0.43 | 6.9 |
| HAB-Pl | Kamenický Šutrák 28 August 2014 | Planktothrix agardhii (92%) [30] | 7.85 (±3.09) | 0.17 | 1.3 |
| HAB-Do | Staňkov 11 September 2013 | Dolichospermum curvum (73.5%) [31] | 12.35 (n.d.) | 0.58 | 7.1 |
| HAB-Mi | Nové Mlýny, 14 August 2012 | Microcystis aeruginosa (99.8%) [32] | 7.13 (±1.16) | 37,202. 67 | 265,092.9 |
| Cyanobacterial cultures | |||||
| Ap | Aphanizomenon flos-aquae PCC 7905 | 45.98 (n.d.) | 1237.34 | 21,158.44 | |
| Pl | Planktothrix aghardii NIVA/CYA 126/8 | 7.60 (n.d.) | 2.01 | 6.96 | |
| Do | Dolichospermum flos-aquae CCALA 007 | 23.43 (±1.54) | 0.67 | 14.63 | |
| Mi | Microcystis aeruginosa PCC 7806 | 13.24 (±2.89) | 1078.50 | 9870.34 | |
| Gram-negative bacteria (positive controls) | |||||
| EC | Escherichia coli O111:B4 | 3689.95 | |||
| PA | Pseudomonas aeruginosa | 387.66 | |||
| Target | Primer Sets and Probe Sequences (5′->3′) |
|---|---|
| Cyanobacteria | Forward: ACGGGTGAGTAACRCGTRA |
| Reverse: CCATGGCGGAAAATTCCCC | |
| Probe: CTCAGTCCCAGTGTGGCTGNTC | |
| Total heterotrophic bacteria | Forward: TCCTACGGGAGGCAGCAGT |
| Reverse: GGACTACCAGGGTATCTAATCCTGTT | |
| Probe: CGTATTACCGCGGCTGCTGGCAC-3′ | |
| Gram-negative bacteria | Forward: AACTGGAGGAAGGTGGGGAT |
| Reverse: AGGAGGTGATCCAACCGCA | |
| Probe: GACGTAAGGGCCATGAGGACTTGACGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skočková, V.; Vašíček, O.; Sychrová, E.; Sovadinová, I.; Babica, P.; Šindlerová, L. Cyanobacterial Harmful Bloom Lipopolysaccharides Induce Pro-Inflammatory Effects in Immune and Intestinal Epithelial Cells In Vitro. Toxins 2023, 15, 169. https://doi.org/10.3390/toxins15030169
Skočková V, Vašíček O, Sychrová E, Sovadinová I, Babica P, Šindlerová L. Cyanobacterial Harmful Bloom Lipopolysaccharides Induce Pro-Inflammatory Effects in Immune and Intestinal Epithelial Cells In Vitro. Toxins. 2023; 15(3):169. https://doi.org/10.3390/toxins15030169
Chicago/Turabian StyleSkočková, Veronika, Ondřej Vašíček, Eliška Sychrová, Iva Sovadinová, Pavel Babica, and Lenka Šindlerová. 2023. "Cyanobacterial Harmful Bloom Lipopolysaccharides Induce Pro-Inflammatory Effects in Immune and Intestinal Epithelial Cells In Vitro" Toxins 15, no. 3: 169. https://doi.org/10.3390/toxins15030169