
Mixed Infection of Blackcurrant with a Novel Cytorhabdovirus and Black Currant-Associated Nucleorhabdovirus
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Morphology and Localization
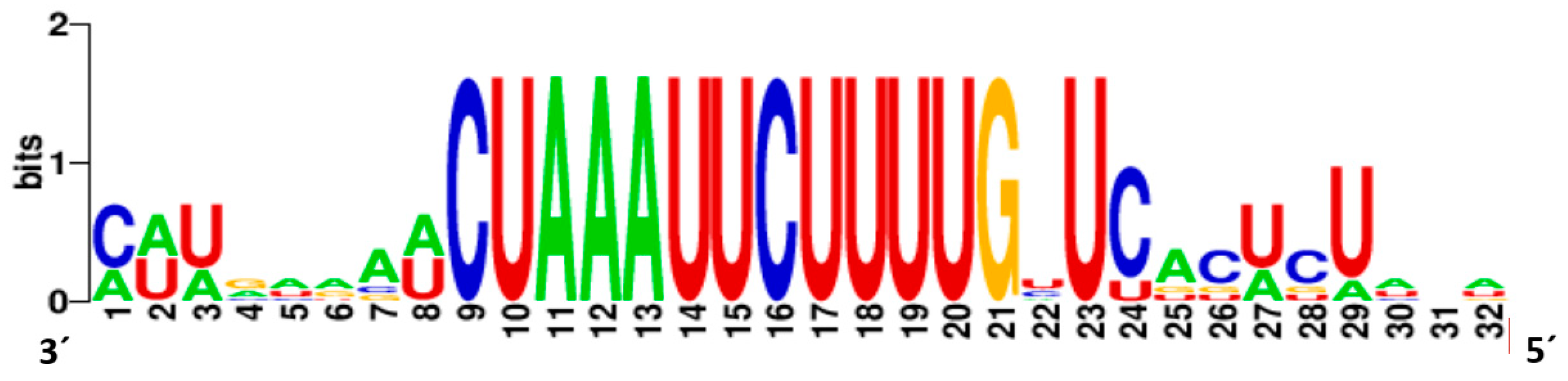
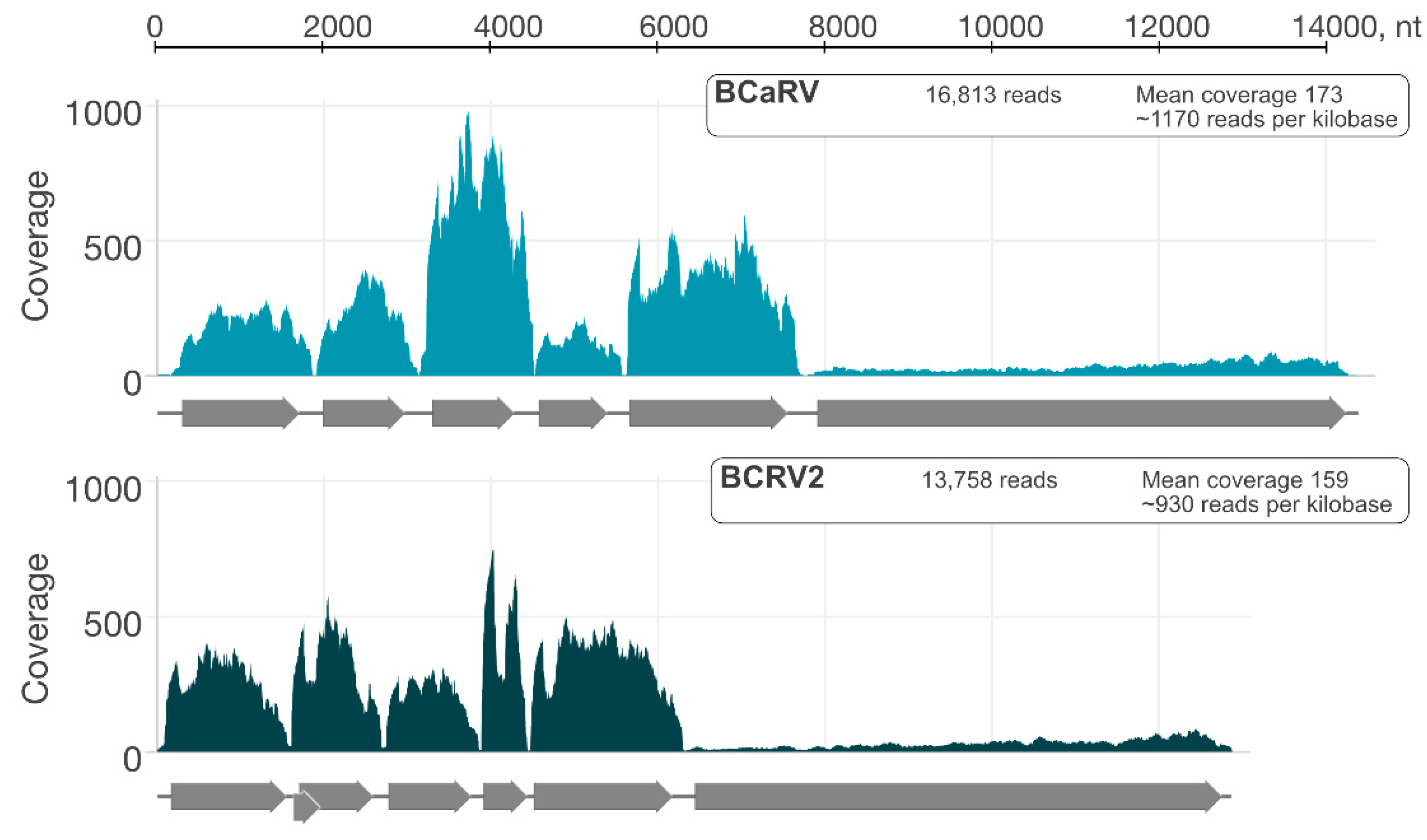
3.2. Genome Organisation
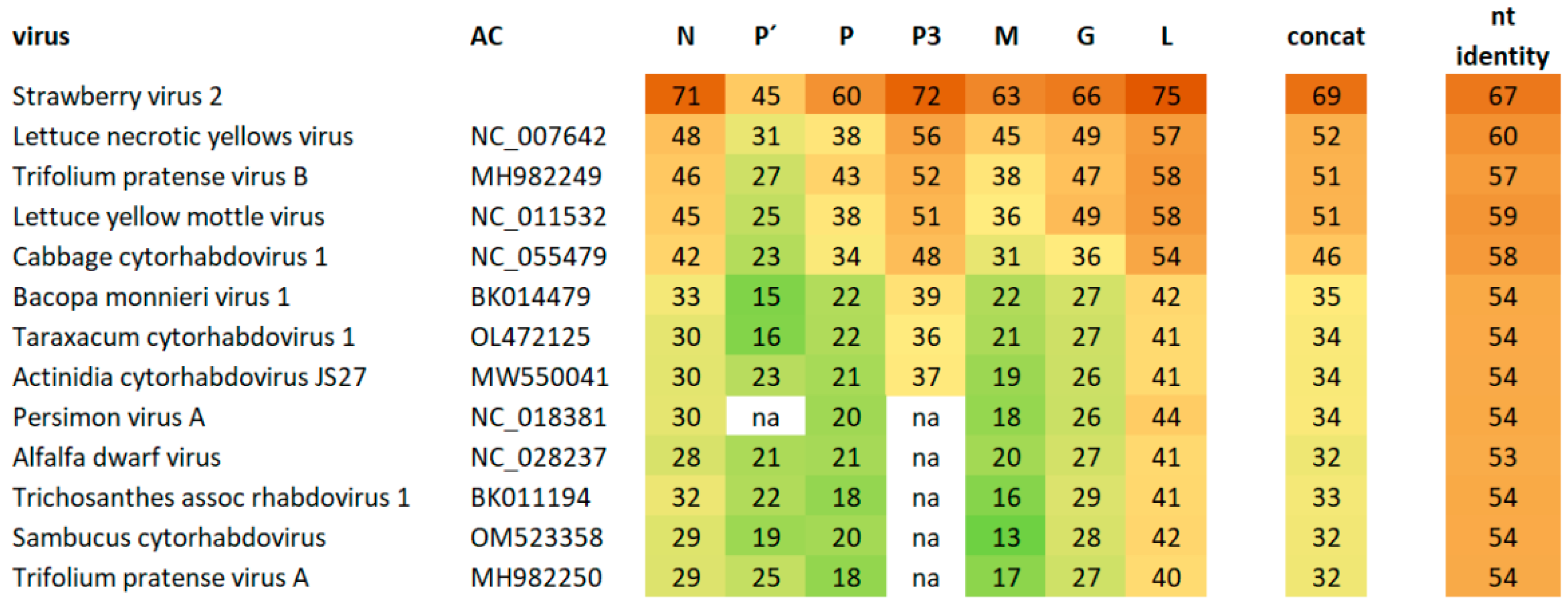
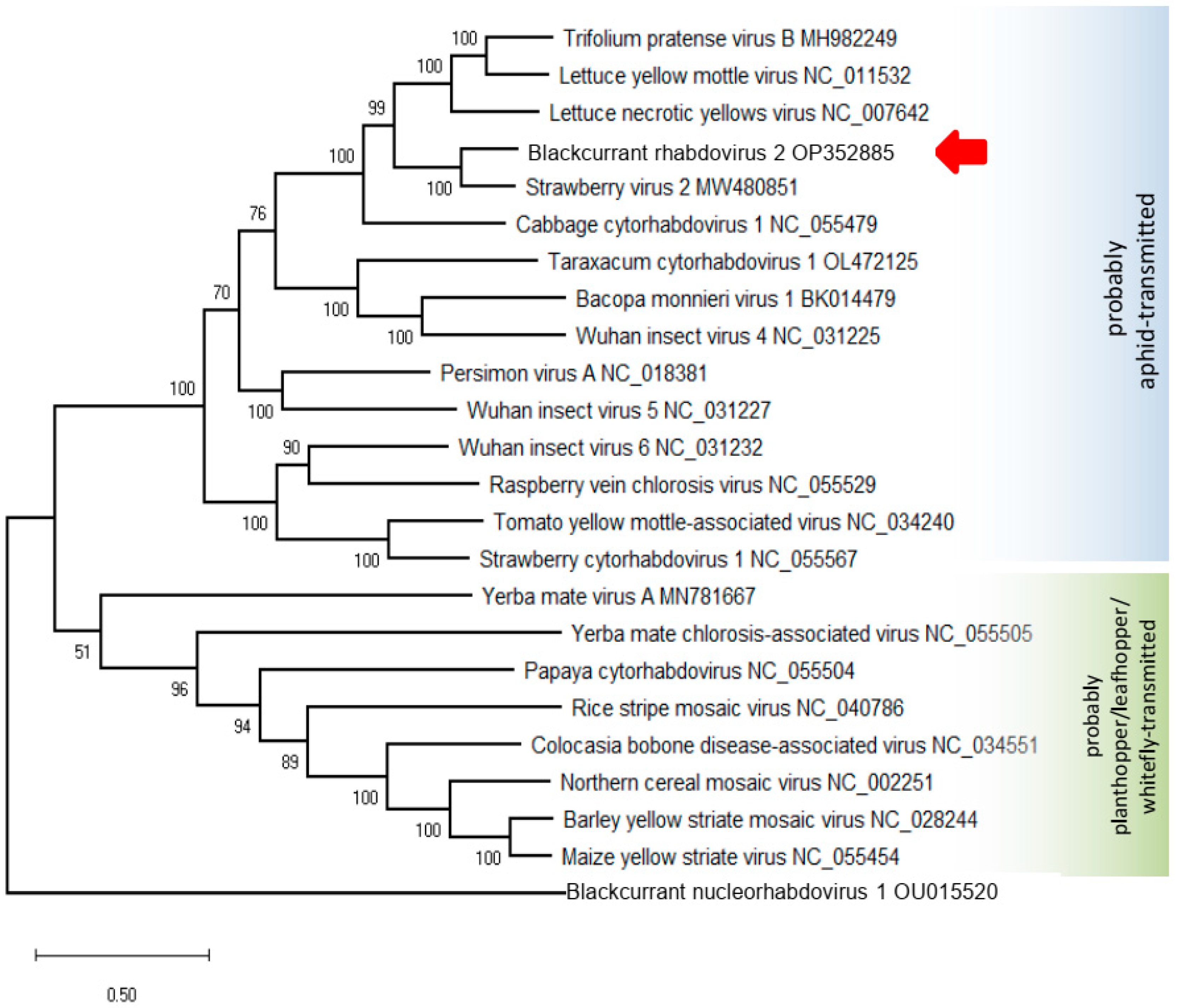
3.3. Phylogeny
3.4. Aphid Vectoring
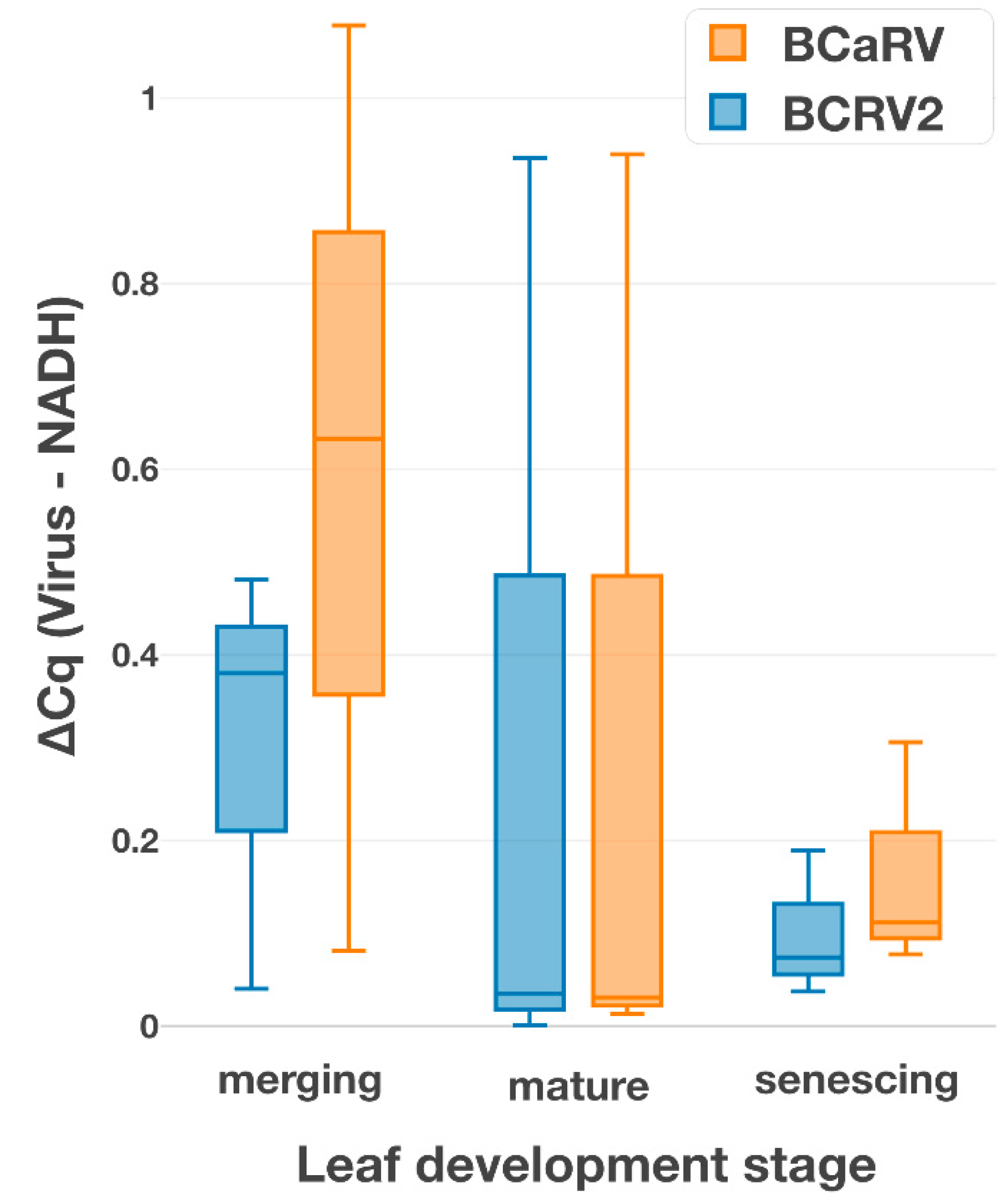
3.5. Relative Titers of the Cytorhabdovirus and the Nucleorhabdovirus in a Single Plant
3.6. Origin of BCaRV and BCRV2
4. Discussion
5. Conclusions
- To our knowledge, this is the first documented case of mixed cyto- and nucleorhabdoviral infection in a single host.
- Interestingly, Aphis schneideri seems to be unable to replicate and transmit BCRV2 and BCaRV. While part of the population of Cryptomyzus galeopsidis was positive for both viruses, it was unable to transmit the viruses to two blackcurrant cultivars.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bejerman, N.; Dietzgen, R.G.; Debat, H. Illuminating the plant rhabdovirus landscape through metatranscriptomics data. Viruses 2021, 13, 1304. [Google Scholar] [CrossRef] [PubMed]
- Bejerman, N.; Dietzgen, R.G.; Debat, H. Unlocking the hidden genetic diversity of varicosaviruses, the neglected plant rhabdoviruses. Pathogens 2022, 11, 1127. [Google Scholar] [CrossRef] [PubMed]
- Petrzik, K.; Vondrák, J.; Barták, M.; Peksa, O.; Kubešová, O. Lichens–new source or yet unknown host of higher plant viruses? Eur. J. Plant Pathol. 2014, 138, 549–559. [Google Scholar] [CrossRef]
- Petrzik, K. Bioinformatics’ analysis of the L polymerase gene lead to discrimination of new rhabdoviruses. J. Phytopathol. 2012, 160, 377–381. [Google Scholar] [CrossRef]
- Mifsud, J.C.O.; Gallagher, R.V.; Holmes, E.C.; Geoghegan, J.L. Transcriptome mining expands knowledge of RNA viruses across the plant kingdom. J. Virol. 2022. ahead of print. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Bejerman, N.E.; Goodin, M.M.; Higgins, C.M.; Huot, O.B.; Kondo, H.; Martin, K.M.; Whitfield, A.E. Diversity and epidemiology of plant rhabdoviruses. Virus Res. 2020, 281, 197942. [Google Scholar] [CrossRef]
- Whitfield, A.E.; Huot, O.B.; Martin, K.M.; Kondo, H.; Dietzgen, R.G. Plant rhabdoviruses-their origins and vector interactions. Curr. Opin. Virol. 2018, 33, 196–207. [Google Scholar] [CrossRef]
- Špak, J.; Koloniuk, I.; Tzanetakis, I.E. Graft-transmissible diseases of Ribes–pathogens, impact, and control. Plant Dis. 2021, 105, 242–250. [Google Scholar] [CrossRef]
- Roberts, I.M.; Jones, A.T. Rhabdovirus-like and closterovirus-like particles in ultrathin sections of Ribes species with symptoms of blackcurrant reversion and gooseberry veinbanding diseases. Ann. Appl. Biol. 1997, 130, 77–89. [Google Scholar] [CrossRef]
- Přibylová, J.; Špak, J.; Kubelková, D. Mixed infection of black currant (Ribes nigrum L.) plants with blackcurrant reversion associated virus and rhabdovirus-like particles with symptoms of black currant reversion disease. Acta Virol. 2002, 46, 253–256. [Google Scholar]
- Wu, L.P.; Yang, T.; Liu, H.W.; Postman, J.; Li, R. Molecular characterization of a novel rhabdovirus infecting blackcurrant identified by high-throughput sequencing. Arch. Virol. 2018, 163, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- Zrelovs, N.; Resevica, G.; Kalnciema, I.; Niedra, H.; Lācis, G.; Bartulsons, T.; Moročko-Bičevska, I.; Stalažs, A.; Drevinska, K.; Zeltins, A.; et al. First report of blackcurrant-associated rhabdovirus in blackcurrants in Latvia. Plant Dis. 2022, 106, 1078. [Google Scholar] [CrossRef] [PubMed]
- Fránová, J.; Sarkisova, T.; Jakešová, H.; Koloniuk, I. Molecular and biological properties of two putative new cytorhabdoviruses infecting Trifolium pratense. Plant Pathol. 2019, 68, 1276–1286. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Kondo, H.; Goodin, M.M.; Kurath, G.; Vasilakis, N. The family Rhabdoviridae: Mono- and bipartite negative-sense RNA viruses with diverse genome organization and common evolutionary origins. Virus Res. 2017, 227, 158–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masny, A.; Pluta, S.; Seliga, L. Breeding value of selected blackcurrant (Ribes nigrum L.) genotypes for early-age fruit yield and its quality. Euphytica 2018, 214, 89. [Google Scholar] [CrossRef]
- Johansen, E.; Edwards, M.C.; Hampton, R.O. Seed transmission of viruses: Current perspectives. Annu. Rev. Phytopathol. 1994, 32, 363–386. [Google Scholar] [CrossRef]
- Xu, Y.G.; Zhao, Y.Q.; Duan, H.M.; Sui, N.; Yuan, F.; Song, J. Transcriptomic profiling of genes in matured dimorphic seeds of euhalophyte Suaeda salsa. BMC Genom. 2017, 18, 727. [Google Scholar] [CrossRef]
- Lee, D.-S.; Kim, J.; Jun, M.; Shin, S.; Lee, S.-J.; Lim, S. Complete genome sequence of a putative novel cytorhabdovirus isolated from Rudbeckia sp. Arch. Virol. 2022, 167, 2381–2385. [Google Scholar] [CrossRef]
- Harry, M.; Solignac, M.; Lachaise, D. Molecular evidence for parallel evolution of adaptive syndromes in fig-breeding Lissocephala (Drosophilidae). Mol. Phylogenet. Evol. 1998, 9, 542–551. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thompson, J.R.; Wetzel, S.; Klerks, M.M.; Vašková, D.; Schoen, C.D.; Špak, J.; Jelkmann, W. Multiplex RT-PCR detection of four aphid-borne strawberry viruses in Fragaria spp. in combination with a plant mRNA specific internal control. J. Virol. Methods 2003, 111, 85–93. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environmnet for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Walker, P.J.; Freitas-Astúa, J.; Bejerman, N.; Blasdell, K.R.; Breyta, R.; Dietzgen, R.G.; Fooks, A.R.; Kondo, H.; Kurath, G.; Kuzmin, I.V.; et al. ICTV virus taxonomy profile: Rhabdoviridae 2022. J. Gen. Virol. 2022, 103, 001689. [Google Scholar] [CrossRef] [PubMed]
- Medberry, A.; Tzanetakis, I.E. Identification, characterization, and detection of a novel strawberry cytorhabdovirus. Plant Dis. 2022, 106, 2784–2787. [Google Scholar] [CrossRef]
- CABI. Crop Protection Compendium; CAB International: Wallingford, UK, 2022; Available online: www.cabi.org/cpc (accessed on 1 September 2022).
- Holman, J. Host Plant Catalog of Aphids: Palaearctic Region; Springer: Berlin/Heidelberg, Germany, 2009; pp. 917–920. ISBN 978-1-4020-8285-6. [Google Scholar]
- Syller, J. Facilitative and antagonistic interactions between plant viruses in mixed infections: Plant virus interactions in mixed infections. Mol. Plant Pathol. 2012, 13, 204–216. [Google Scholar] [CrossRef] [PubMed]
- Valentova, L.; Rejlova, M.; Franova, J.; Cmejla, R. Symptomless infection by strawberry virus 1 (StrV-1) leads to losses in strawberry yields. Plant Pathol. 2022, 71, 1220–1228. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrzik, K.; Přibylová, J.; Špak, J.; Sarkisova, T.; Fránová, J.; Holub, J.; Skalík, J.; Koloniuk, I. Mixed Infection of Blackcurrant with a Novel Cytorhabdovirus and Black Currant-Associated Nucleorhabdovirus. Viruses 2022, 14, 2456. https://doi.org/10.3390/v14112456
Petrzik K, Přibylová J, Špak J, Sarkisova T, Fránová J, Holub J, Skalík J, Koloniuk I. Mixed Infection of Blackcurrant with a Novel Cytorhabdovirus and Black Currant-Associated Nucleorhabdovirus. Viruses. 2022; 14(11):2456. https://doi.org/10.3390/v14112456
Chicago/Turabian StylePetrzik, Karel, Jaroslava Přibylová, Josef Špak, Tatiana Sarkisova, Jana Fránová, Jan Holub, Jan Skalík, and Igor Koloniuk. 2022. "Mixed Infection of Blackcurrant with a Novel Cytorhabdovirus and Black Currant-Associated Nucleorhabdovirus" Viruses 14, no. 11: 2456. https://doi.org/10.3390/v14112456