
Knock-Out of Retrovirus Receptor Gene Tva in the Chicken Confers Resistance to Avian Leukosis Virus Subgroups A and K and Affects Cobalamin (Vitamin B12)-Dependent Level of Methylmalonic Acid

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Chickens
2.3. PGC Cultivation and Tva Editing
2.4. Preparation of Sterile Recipient Roosters and PGC Transplantation
2.5. Breeding of Tva KO Chickens
2.6. Off-Target Site Selection and Analysis
2.7. In Vivo Infection of Chickens and Viremia Analysis
2.8. In Vitro Virus Spread in Infected Chicken Embryo Fibroblasts
2.9. In Vivo Induction of Tumors with Transforming Virus Pseudotype
2.10. Quantification of MMA Serum Levels
3. Results
3.1. Introduction of Tva Frame-Shift Deletions in Chicken PGCs
3.2. Generation of Tva KO, the Tva −/− Chicken Line
3.3. Absence of Mutations at Potential Off-Target Sites
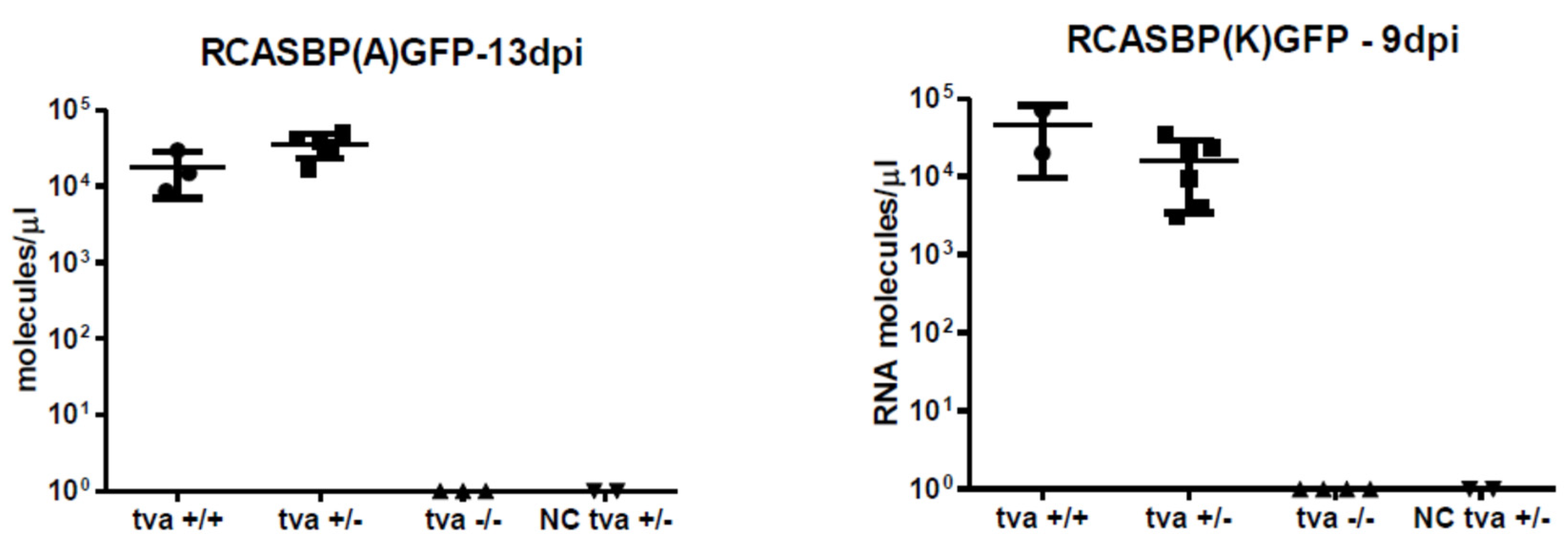
3.4. Tva −/− Chickens Are Resistant to Infection with ALV-A and ALV-K
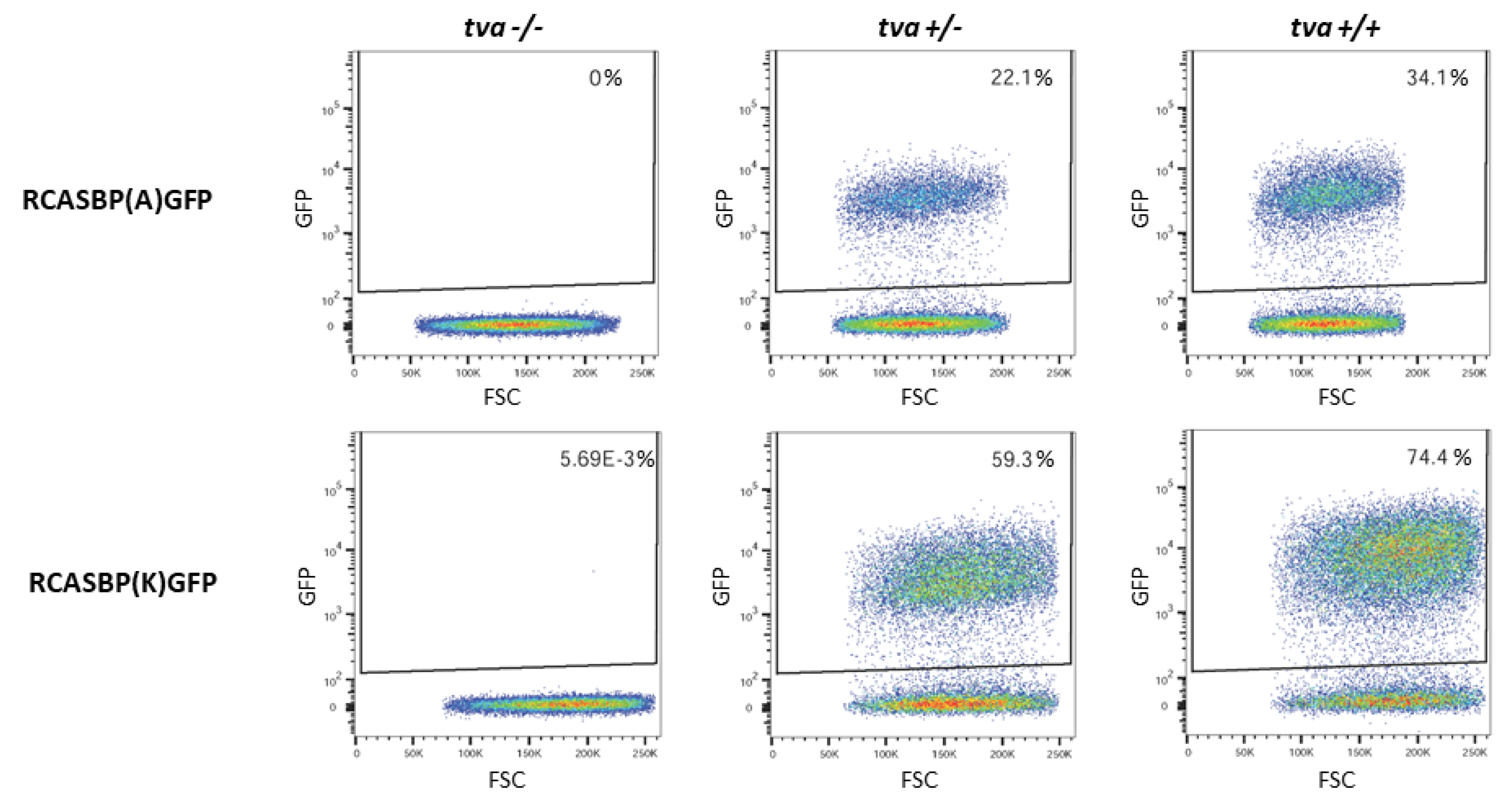
3.5. In Vitro Cultured Tva −/− Embryo Fibroblasts Do Not Support the Spread of ALV-A and ALV-K
3.6. Tva KO Chickens Are Resistant to an ASLV-A/K-Pseudotyped Transforming Virus
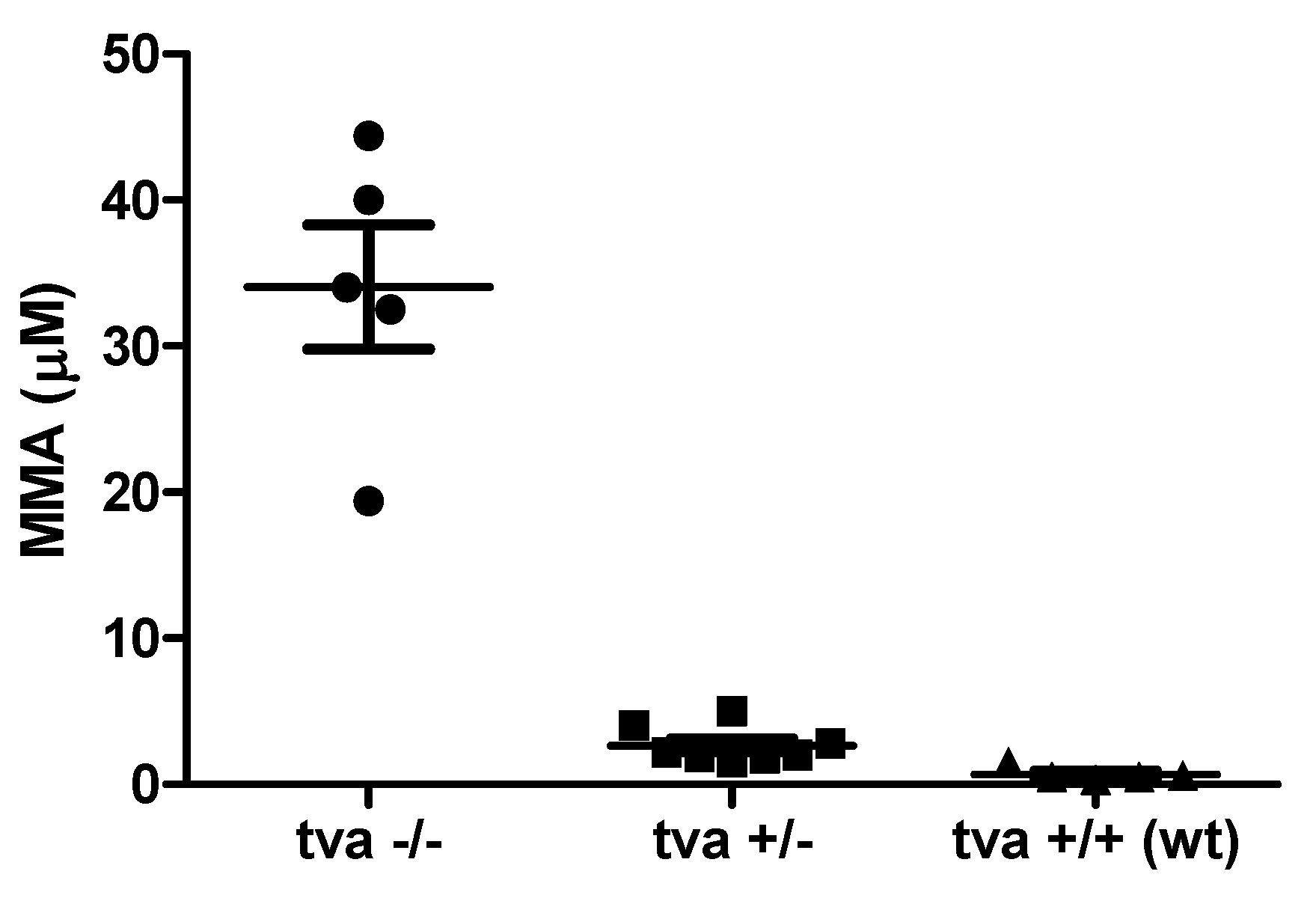
3.7. TvaKO Chickens Have Increased Serum Levels of Methylmalonic Acid
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Federspiel, M.J. Reverse engineering provides insight on the evolution of subgroups A to E avian sarcoma and leukosis virus receptor specificity. Viruses 2019, 11, 497. [Google Scholar] [CrossRef] [Green Version]
- Bates, P.; Young, J.A.; Varmus, H.E. A receptor for subgroup a Rous sarcoma virus is related to the low density lipoprotein receptor. Cell 1993, 74, 1043–1051. [Google Scholar] [CrossRef]
- Young, J.A.T.; Bates, P.; Varmus, H.E. Isolation of a chicken gene that confers susceptibility to infection by subgroup A avian leukosis and sarcoma viruses. J. Virol. 1993, 67, 1811–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brojatsch, J.; Naughton, J.; Rolls, M.M.; Zingler, K.; Young, J.A. CAR1, a TNFR-related protein, is a cellular receptor for cytopathic avian leukosis-sarcoma viruses and mediates apoptosis. Cell 1996, 87, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Adkins, H.B.; Brojatsch, J.; Naughton, J.; Rolls, M.M.; Pesola, J.M.; Young, J.A. Identification of a cellular receptor for subgroup E avian leukosis virus. Proc. Natl. Acad. Sci. USA 1997, 94, 11617–11622. [Google Scholar] [CrossRef] [Green Version]
- Adkins, H.B.; Brojatsch, J.; Young, J.A.T. Identification and characterization of a shared TNFR-related receptor for subgroup B, D, and E avian leukosis viruses reveal cysteine residues required specifically for subgroup E viral entry. J. Virol. 2000, 74, 3572–3578. [Google Scholar] [CrossRef] [Green Version]
- Elleder, D.; Stepanets, V.; Melder, D.C.; Šenigl, F.; Geryk, J.; Pajer, P.; Plachý, J.; Hejnar, J.; Svoboda, J.; Federspiel, M.J. The receptor for the subgroup C avian sarcoma and leukosis viruses, Tvc, is related to mammalian butyrophilins, members of the immunoglobulin superfamily. J. Virol. 2005, 79, 10408–10419. [Google Scholar] [CrossRef] [Green Version]
- Chai, N.; Bates, P. Na+/H+ exchanger type 1 is a receptor for pathogenic subgroup J avian leukosis virus. Proc. Natl. Acad. Sci. USA 2006, 103, 5531–5536. [Google Scholar] [CrossRef] [Green Version]
- Elleder, D.; Melder, D.C.; Trejbalova, K.; Svoboda, J.; Federspiel, M.J. Two different molecular defects in the Tva receptor gene explain the resistance of two tvar lines of chickens to infection by subgroup A avian sarcoma and leukosis viruses. J. Virol. 2004, 78, 13489–13500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klucking, S.; Adkins, H.B.; Young, J.A.T. Resistance to infection by subgroups B, D, and E avian sarcoma and leukosis viruses is explained by a premature stop codon within a resistance allele of the tvb receptor gene. J. Virol. 2002, 76, 7918–7921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinišová, M.; Šenigl, F.; Yin, X.; Plachý, J.; Geryk, J.; Elleder, D.; Svoboda, J.; Federspiel, M.J.; Hejnar, J. A single-amino-acid substitution in the TvbS1 receptor results in decreased susceptibility to infection by avian sarcoma and leukosis virus subgroups B and D and resistance to infection by subgroup E in vitro and in vivo. J. Virol. 2008, 82, 2097–2105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Přikryl, D.; Plachý, J.; Kučerová, D.; Koslová, A.; Reinišová, M.; Šenigl, F.; Hejnar, J. The Novel Avian Leukosis Virus Subgroup K Shares Its Cellular Receptor with Subgroup A. J. Virol. 2019, 93, e00580-19. [Google Scholar] [CrossRef] [Green Version]
- Rong, L.; Bates, P. Analysis of the subgroup a avian sarcoma and leukosis virus receptor: The 40-residue, cysteine-rich, low-density lipoprotein receptor repeat motif of tva is sufficient to mediate viral entry. J. Virol. 1995, 69, 4847–4853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zingler, K.; Belanger, C.; Peters, R.; Agard, D.; Young, J.A.T. Identification and characterization of the viral interaction determinant of the subgroup a avian leukosis virus receptor. J. Virol. 1995, 69, 4261–4266. [Google Scholar] [CrossRef] [Green Version]
- Rong, L.; Gendron, K.; Bates, P. Conversion of a human low-density lipoprotein receptor ligand-binding repeat to a virus receptor: Identification of residues important for ligand specificity. Proc. Natl. Acad. Sci. USA 1998, 95, 8467–8472. [Google Scholar] [CrossRef] [Green Version]
- Reinišová, M.; Plachý, J.; Trejbalová, K.; Šenigl, F.; Kučerová, D.; Geryk, J.; Svoboda, J.; Hejnar, J. Intronic deletions that disrupt mRNA splicing of the tva receptor gene result in decreased susceptibility to infection by avian sarcoma and leukosis virus subgroup A. J. Virol. 2012, 86, 2021–2030. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.G.; Liu, Y.; Li, H.X.; Chang, S.; Shu, D.M.; Zhang, H.M.; Chen, F.; Xie, Q. Intronic deletions of tva receptor gene decrease the susceptibility to infection by avian sarcoma and leukosis virus subgroup A. Sci. Rep. 2015, 5, 9900. [Google Scholar] [CrossRef] [Green Version]
- Koslová, A.; Kučerová, D.; Reinišová, M.; Geryk, J.; Trefil, P.; Hejnar, J. Genetic Resistance to Avian Leukosis Viruses Induced by CRISPR/Cas9 Editing of Specific Receptor Genes in Chicken Cells. Viruses 2018, 10, 605. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Park, K.J.; Lee, K.Y.; Yao, Y.X.; Nair, V.; Han, J.Y. Sequential disruption of ALV host receptor genes reveals no sharing of receptors between ALV subgroups A, B, and J. J. Anim. Sci. Biotechnol. 2019, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadros, E.V.; Nakayama, Y.; Sequeira, J.M. The Protein and the Gene Encoding the Receptor for the Cellular Uptake of Transcobalamin-Bound Cobalamin. Blood 2009, 113, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Krchlíková, V.; Mikešová, J.; Geryk, J.; Bařinka, C.; Nexo, E.; Fedosov, S.N.; Kosla, J.; Kučerová, D.; Reinišová, M.; Hejnar, J.; et al. The avian retroviral receptor Tva mediates the uptake of transcobalamin 1 bound vitamin B12 (cobalamin). J. Virol. 2021, 95, e02136-20. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Allen, L.H.; Bjørke-Monsen, A.-L.; Brito, A.; Guéant, J.-L.; Miller, J.W.; Molloy, A.M.; Nexo, E.; Stabler, S.; Toh, B.H.; et al. Vitamin B12 Deficiency. Nat. Rev. Dis. Primers 2017, 3, 17040. [Google Scholar] [CrossRef]
- Lai, S.C.; Nakayama, Y.; Sequeira, J.M.; Wlodarczyk, B.J.; Cabrera, R.M.; Finnell, R.H.; Bottiglieri, T.; Quadros, E.V. The Transcobalamin Receptor Knockout Mouse: A Model for Vitamin B12 Deficiency in the Central Nervous System. FASEB J. 2013, 27, 2468–2475. [Google Scholar] [CrossRef] [Green Version]
- Arora, K.; Sequeira, J.M.; Alarcon, J.M.; Wasek, B.; Arning, E.; Bottiglieri, T.; Quadros, E.V. Neuropathology of Vitamin B Deficiency in the Cd320 Mouse. FASEB J. 2019, 33, 2563–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trefil, P.; Mičáková, A.; Mucksová, J.; Hejnar, J.; Poplštein, M.; Bakst, M.R.; Kalina, J.; Brillard, J.P. Restoration of spermatogenesis and male fertility by transplantation of dispersed testicular cells in the chicken. Biol. Reprod. 2006, 75, 575–581. [Google Scholar] [CrossRef]
- Trefil, P.; Aumann, D.; Koslová, A.; Mucksová, J.; Benešová, B.; Kalina, J.; Wurmser, C.; Fries, R.; Elleder, D.; Schusser, B.; et al. Male fertility restored by transplanting primordial germ cells into testes: A new way towards efficient transgenesis in chicken. Sci. Rep. 2017, 7, 14246. [Google Scholar] [CrossRef]
- Plachý, J. The chicken–A laboratory animal of the class Aves. Folia Biol. 2000, 46, 17–23. [Google Scholar]
- Mucksová, J.; Reinišová, M.; Kalina, J.; Lejčková, B.; Hejnar, J.; Trefil, P. Conservation of chicken male germline by orthotopic transplantation of primordial germ cells from genetically distant donors. Biol. Reprod. 2019, 101, 200–207. [Google Scholar] [CrossRef]
- Whyte, J.; Glover, J.D.; Woodcock, M.; Brzeszczynska, J.; Taylor, L.; Sherman, A.; Kaiser, P.; McGrew, M.J. FGF, insulin, and SMAD signaling cooperate for avian primordial germ cell self-renewal. Stem Cell Rep. 2015, 5, 1171–1182. [Google Scholar] [CrossRef] [Green Version]
- Koslová, A.; Trefil, P.; Mucksová, J.; Reinišová, M.; Plachý, J.; Kalina, J.; Kučerová, D.; Geryk, J.; Krchlíková, V.; Lejčková, B.; et al. Precise CRISPR/Cas9 editing of the NHE1 gene renders chickens resistant to the J subgroup of avian leukosis virus. Proc. Natl. Acad. Sci. USA 2020, 117, 2108–2112. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Federspiel, M.J.; Hughes, S.H. Retroviral gene delivery. Methods Cell Biol. 1997, 52, 179–214. [Google Scholar]
- Moscovici, C.; Moscovici, M.G.; Jimenez, H.; Lai, M.M.; Hayman, M.J.; Vogt, P.K. Continuous tissue culture cell lines derived from chemically induced tumors of Japanese quail. Cell 1977, 11, 95–103. [Google Scholar] [CrossRef]
- Kheimar, A.; Klinger, R.; Bertzbach, L.D.; Sid, H.; Yu, Y.; Conradie, A.M.; Schade, B.; Böhm, B.; Preisinger, R.; Nair, V.; et al. A Genetically Engineered Commercial Chicken Line Is Resistant to Highly Pathogenic Avian Leukosis Virus Subgroup J. Microorganisms 2021, 9, 1066. [Google Scholar] [CrossRef]
- Whitworth, K.M.; Rowland, R.R.; Ewen, C.L.; Trible, B.R.; Kerrigan, M.A.; Cino-Ozuna, A.G.; Samuel, M.S.; Lightner, J.E.; McLaren, D.G.; Mileham, A.J.; et al. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus. Nat. Biotechnol. 2016, 34, 20–22. [Google Scholar] [CrossRef]
- Xu, K.; Zhou, Y.; Mu, Y.; Liu, Z.; Hou, S.; Xiong, Y.; Fang, L.; Ge, C.; Wei, Y.; Zhang, X.; et al. CD163 and pAPN double-knockout pigs are resistant to PRRSV and TGEV and exhibit decreased susceptibility to PDCoV while maintaining normal production performance. Elife 2020, 9, e57132. [Google Scholar] [CrossRef]
- van de Lavoir, M.C.; Diamond, J.H.; Leighton, P.A.; Mather-Love, C.; Heyer, B.S.; Bradshaw, R.; Kerchner, A.; Hooi, L.T.; Gessaro, T.M.; Swanberg, S.E.; et al. Germline transmission of genetically modified primordial germ cells. Nature 2006, 441, 766–769. [Google Scholar] [CrossRef]
- Schusser, B.; Collarini, E.J.; Yi, H.; Izquierdo, S.M.; Fesler, J.; Pedersen, D.; Klasing, K.C.; Kaspers, B.; Harriman, W.D.; van de Lavoir, M.C.; et al. Immunoglobulin knockout chickens via efficient homologous recombination in primordial germ cells. Proc. Natl. Acad. Sci. USA 2013, 110, 20170–20175. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.D.; Lee, J.H.; Song, S.; Kim, S.W.; Han, J.S.; Shin, S.P.; Park, B.C.; Park, T.S. Generation of myostatin-knockout chickens mediated by D10A-Cas9 nickase. FASEB J. 2020, 34, 5688–5696. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Ma, J.; Lee, K. Direct delivery of adenoviral CRISPR/Cas9 vector into the blastoderm for generation of targeted gene knockout in quail. Proc. Natl. Acad. Sci. USA 2019, 116, 13288–13292. [Google Scholar] [CrossRef] [Green Version]
- Rieblinger, B.; Sid, H.; Duda, D.; Bozoglu, T.; Klinger, R.; Schlickenrieder, A.; Lengyel, K.; Flisikowski, K.; Flisikowska, T.; Simm, N.; et al. Cas9-expressing chickens and pigs as resources for genome editing in livestock. Proc. Natl. Acad. Sci. USA 2021, 118, e2022562118. [Google Scholar] [CrossRef]
- Ballantyne, M.; Woodcock, M.; Doddamani, D.; Hu, T.; Taylor, L.; Hawken, R.J.; McGrew, M.J. Direct allele introgression into pure chicken breeds using Sire Dam Surrogate (SDS) mating. Nat. Commun. 2021, 12, 659. [Google Scholar] [CrossRef]
- Lopes-Marques, M.; Ruivo, R.; Delgado, I.; Wilson, J.M.; Aluru, N.; Castro, L.F.C. Basal gnathostomes provide unique insights into the evolution of vitamin B12 binders. Genome Biol. 2015, 7, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Greibe, E.; Fedosov, S.; Nexo, E. The cobalamin-binding protein in zebrafish is an intermediate between the three cobalamin-binding proteins in human. PLoS ONE 2012, 7, 35660. [Google Scholar] [CrossRef]
- Selhub, J. Folate, vitamin B12 and vitamin B6 and one carbon metabolism. J. Nutr. Health Aging 2002, 6, 39–42. [Google Scholar]
- Fernàndez-Roig, S.; Lai, S.-C.; Murphy, M.M.; Fernandez-Ballart, J.; Quadros, E.V. Vitamin B12 deficiency in the brain leads to DNA hypomethylation in the TCblR/CD320 knockout mouse. Nutr. Metab. 2012, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Li, J.; Casaccia-Bonnefil, P. Histone modifications affect timing of oligodendrocyte progenitor differentiation in the developing rat brain. J. Cell Biol. 2005, 169, 577–589. [Google Scholar] [CrossRef]
- Arora, K.; Sequeira, J.M.; Hernández, A.I.; Alarcon, J.M.; Quadros, E.V. Behavioral alterations are associated with vitamin B12 deficiency in the transcobalamin receptor/CD320 KO mouse. PLoS ONE 2017, 12, e0177156. [Google Scholar] [CrossRef]
- Quadros, E.V.; Lai, S.C.; Nakayama, Y.; Sequeira, J.M.; Hannibal, L.; Wang, S.; Jacobsen, D.W.; Fedosov, S.; Wright, E.; Gallagher, R.C.; et al. Positive newborn screen for methylmalonic aciduria identifies the first mutation in TCblR/CD320, the gene for cellular uptake of transcobalamin-bound vitamin B(12). Hum. Mutat. 2010, 31, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Hannah-Shmouni, F.; Cruz, V.; Schulze, A.; Mercimek-Andrews, S. Transcobalamin receptor defect: Identification of two new cases through positive newborn screening for propionic/methylmalonic aciduria and long-term outcome. Am. J. Med. Genet. A 2018, 176, 1411–1415. [Google Scholar] [CrossRef]
- Reinišová, M.; Plachý, J.; Kučerová, D.; Šenigl, F.; Vinkler, M.; Hejnar, J. Genetic diversity of NHE1, receptor for subgroup J avian leukosis virus, in domestic chicken and wild anseriform species. PLoS ONE 2016, 11, e0150589. [Google Scholar] [CrossRef]
- Kučerová, D.; Plachý, J.; Reinišová, M.; Šenigl, F.; Trejbalová, K.; Geryk, J.; Hejnar, J. Nonconserved tryptophan 38 of the cell surface receptor for subgroup J avian leukosis virus discriminates sensitive from resistant avian species. J. Virol. 2013, 87, 8399–8407. [Google Scholar] [CrossRef] [Green Version]
- Plachý, J.; Reinišová, M.; Kučerová, D.; Šenigl, F.; Stepanets, V.; Hron, T.; Trejbalová, K.; Elleder, D.; Hejnar, J. Identification of New World quails susceptible to infection with avian leukosis virus subgroup J. J. Virol. 2017, 91, e02002-16.s. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rooster No. (No. of PGC Clone) | Restoration of Spermatogenesis (Weeks after Transplantation) | Hens Inseminated a | Hatched Chickens (tva +/−) a |
|---|---|---|---|
| 1 (2) 16 (2) | 10 11 | 2/3 4/3 | 8 (5♂/3♀) 6 (4♂/2♀) |
| RCASBP(A)GFP | RCASBP(K)GFP | ||||
|---|---|---|---|---|---|
| Genotype | Chicken No. | Virus Titer a | Genotype | Chicken No. | Virus Titer a |
| tva −/− | 52 | 0/0 | tva −/− | 129 | 0/0 |
| tva −/− | 58 | 0/0 | tva −/− | 150 | 0/0 |
| tva −/− | 62 | 0/0 | tva −/− | 155 | 0/0 |
| tva +/− | 51 | 102/103 | tva +/− | 125 | 103/0 |
| tva +/− | 53 | 104/103 | tva +/− | 126 | 102/0 |
| tva +/− | 55 | 103/102 | tva +/− | 127 | 101/0 |
| tva +/− | 57 | 102/103 | tva +/− | 130 | 101/0 |
| tva +/− | 59 | 103/103 | tva +/− | 151 | 102/0 |
| tva +/+ | 50 | 102/103 | tva +/− | 152 | 102/101 |
| tva +/+ | 54 | 103/104 | tva +/− | 156 | 102/0 |
| tva +/+ | 61 | 0/102 | tva +/+ | 128 | 101/0 |
| tva +/+ | 131 | 102/0 | |||
| tva +/+ | 132 | 0/0 | |||
| tva +/+ | 133 | 101/0 | |||
| tva +/+ | 134 | 101/0 | |||
| tva +/+ | 148 | 102/0 | |||
| tva +/+ | 154 | 102/0 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koslová, A.; Trefil, P.; Mucksová, J.; Krchlíková, V.; Plachý, J.; Krijt, J.; Reinišová, M.; Kučerová, D.; Geryk, J.; Kalina, J.; et al. Knock-Out of Retrovirus Receptor Gene Tva in the Chicken Confers Resistance to Avian Leukosis Virus Subgroups A and K and Affects Cobalamin (Vitamin B12)-Dependent Level of Methylmalonic Acid. Viruses 2021, 13, 2504. https://doi.org/10.3390/v13122504
Koslová A, Trefil P, Mucksová J, Krchlíková V, Plachý J, Krijt J, Reinišová M, Kučerová D, Geryk J, Kalina J, et al. Knock-Out of Retrovirus Receptor Gene Tva in the Chicken Confers Resistance to Avian Leukosis Virus Subgroups A and K and Affects Cobalamin (Vitamin B12)-Dependent Level of Methylmalonic Acid. Viruses. 2021; 13(12):2504. https://doi.org/10.3390/v13122504
Chicago/Turabian StyleKoslová, Anna, Pavel Trefil, Jitka Mucksová, Veronika Krchlíková, Jiří Plachý, Jakub Krijt, Markéta Reinišová, Dana Kučerová, Josef Geryk, Jiří Kalina, and et al. 2021. "Knock-Out of Retrovirus Receptor Gene Tva in the Chicken Confers Resistance to Avian Leukosis Virus Subgroups A and K and Affects Cobalamin (Vitamin B12)-Dependent Level of Methylmalonic Acid" Viruses 13, no. 12: 2504. https://doi.org/10.3390/v13122504