
Wind Damage and Temperature Effect on Tree Mortality Caused by Ips typographus L.: Phase Transition Model

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
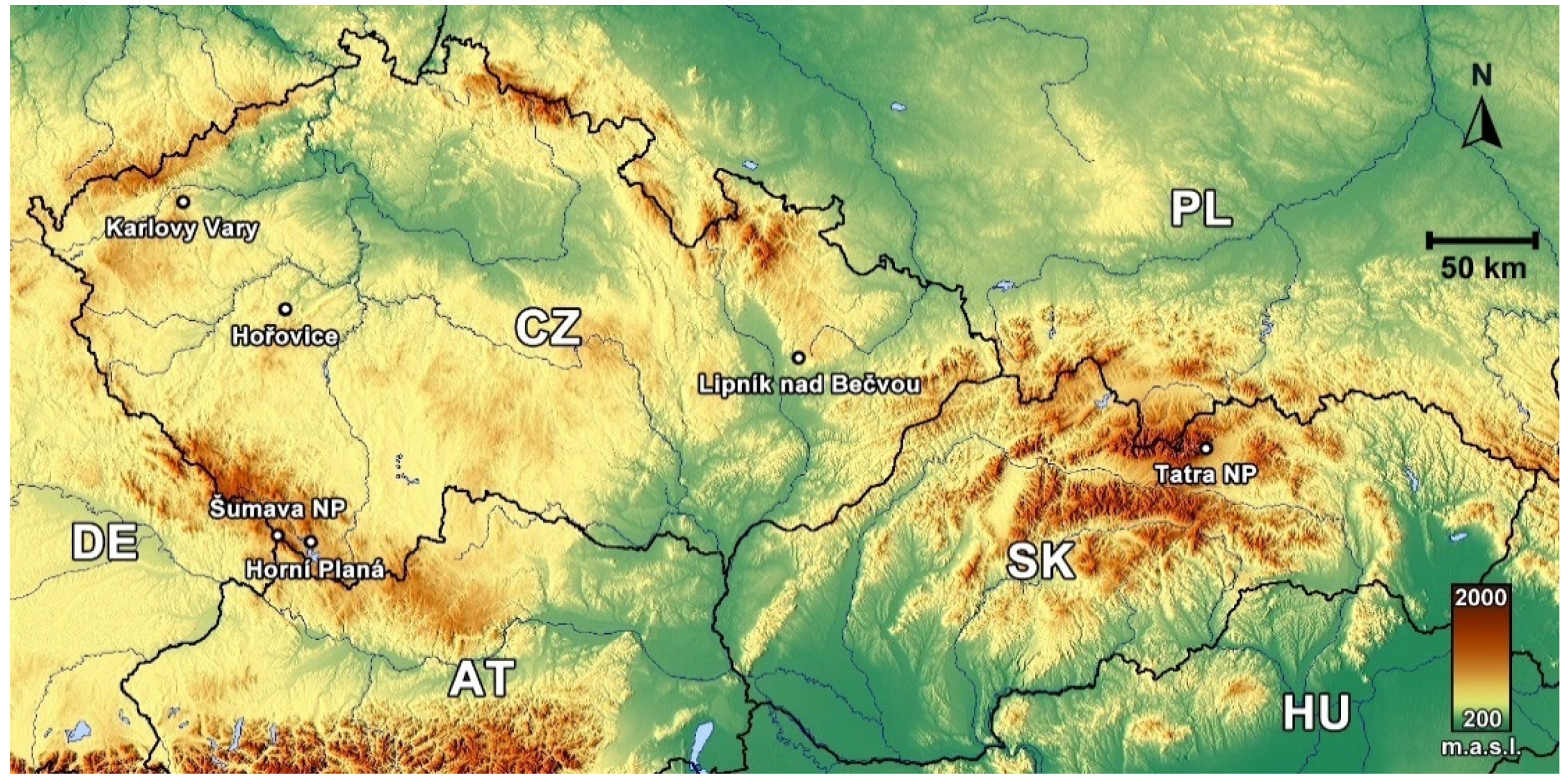
2.1. Study Area
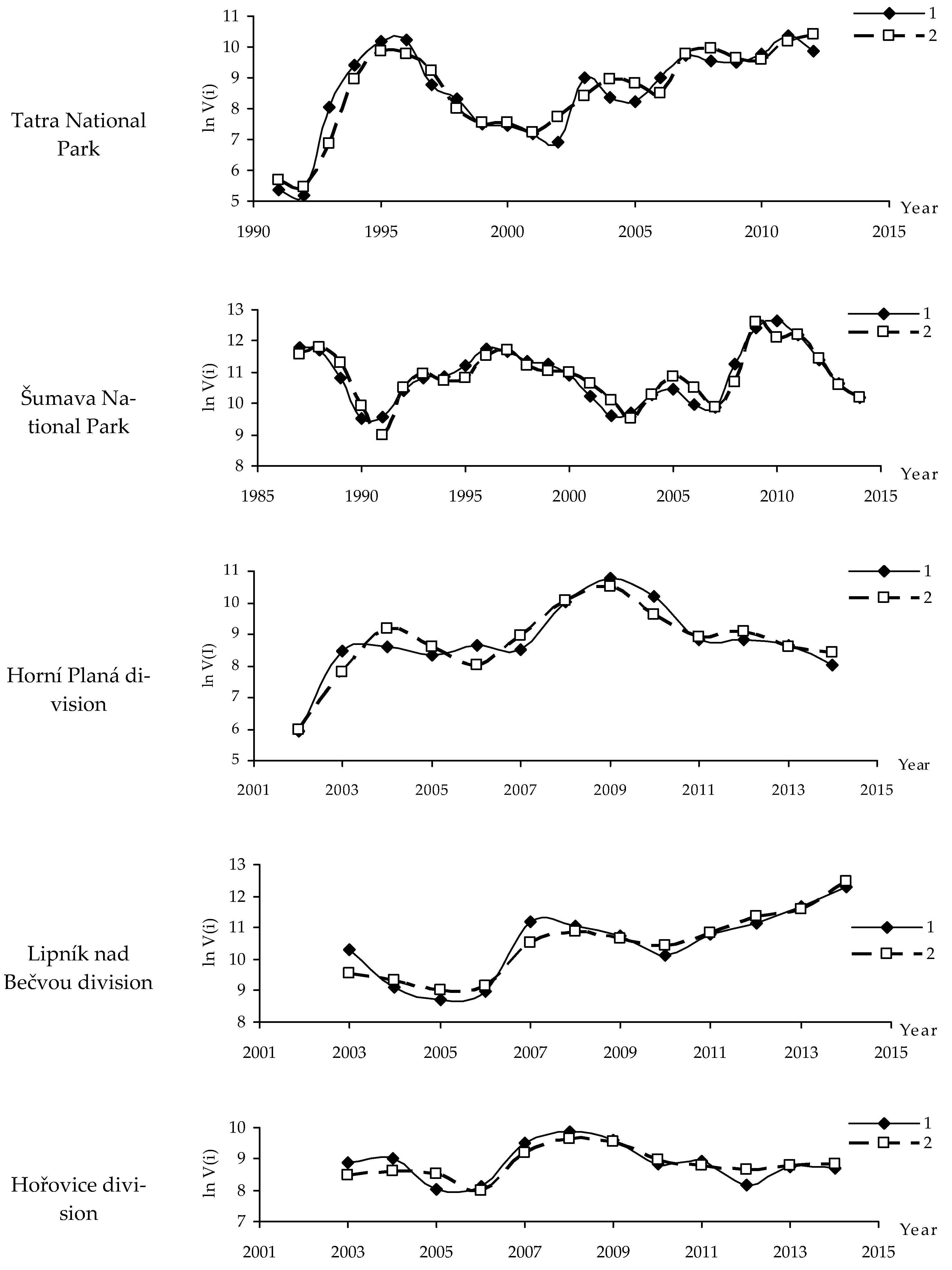
2.1.1. Tatra National Park
2.1.2. Šumava National Park
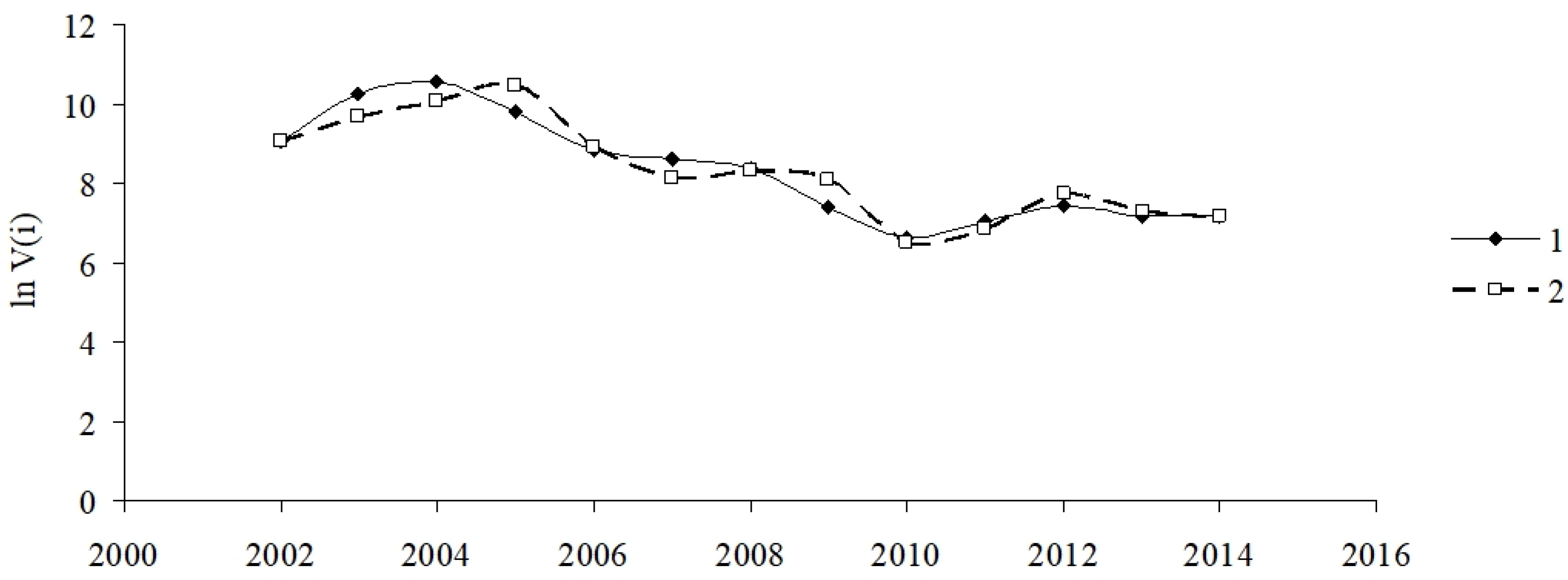
2.1.3. Karlovy Vary Division
2.1.4. Horní Planá Division
2.1.5. Lipník nad Bečvou Division
2.1.6. Hořovice Division
2.2. Tree Mortality Data
2.3. Meteorological Data
2.4. Modelling Design
- A determination coefficient R2, characterizing proportion of variance explained by model. The adjusted determination coefficient (adj.R2) is used to account for influence of variables number. In this case, variables number for different models is the same, so R2 is shifted in relation to adj.R2 by the same value.
- t-test for estimation difference of model coefficients from zero.
- F-test to test hypothesis that all coefficients of linear model are equal to zero.
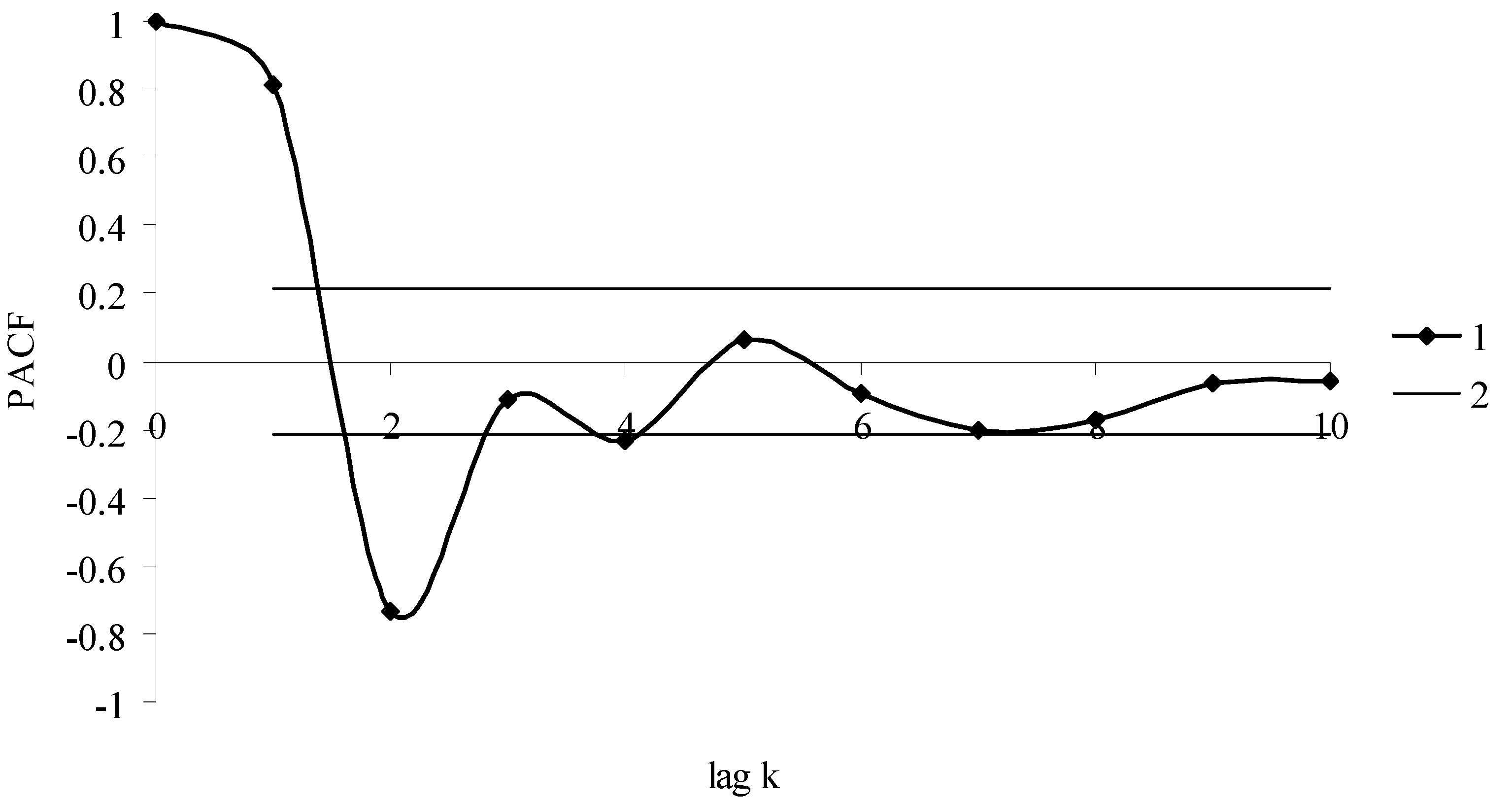
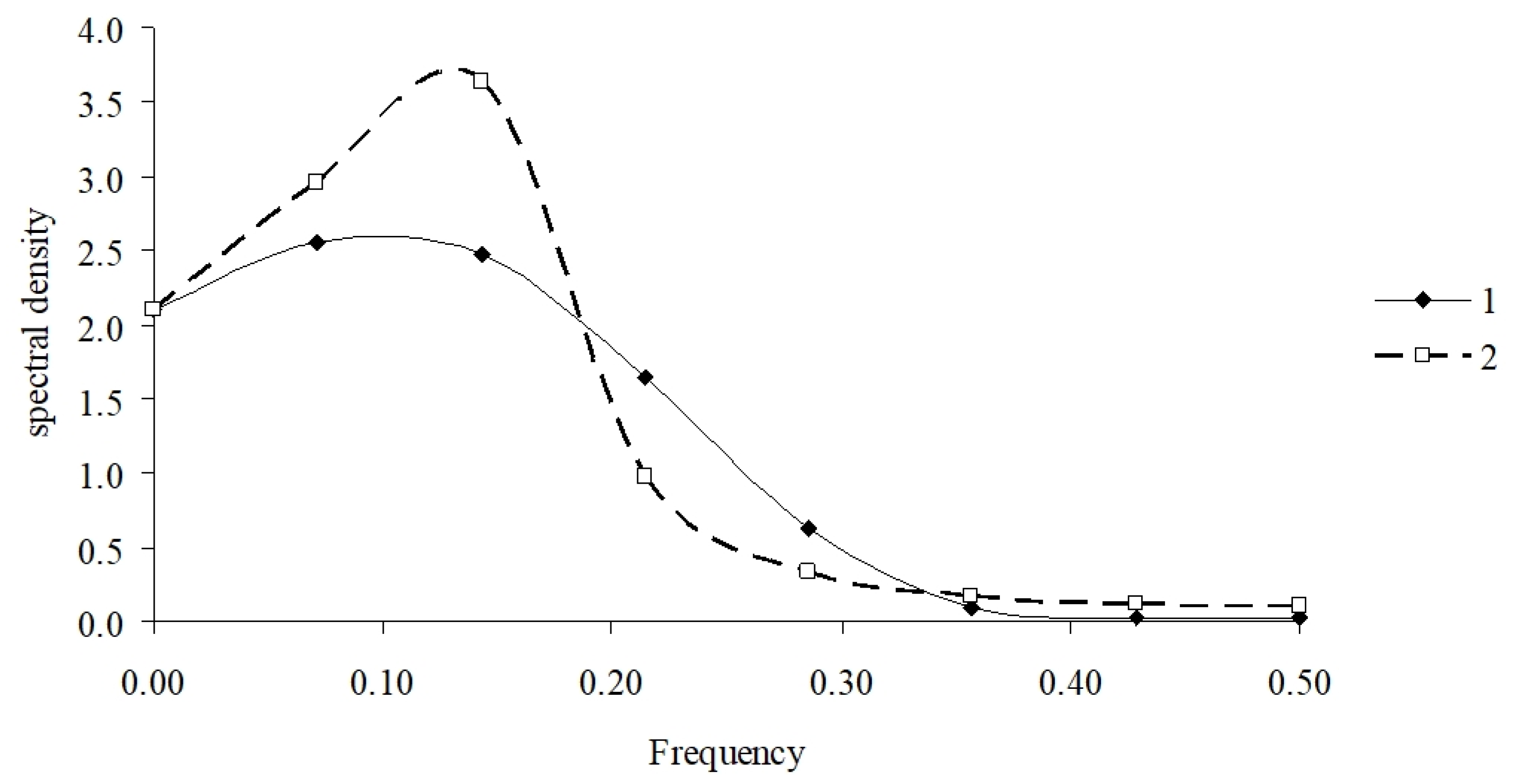
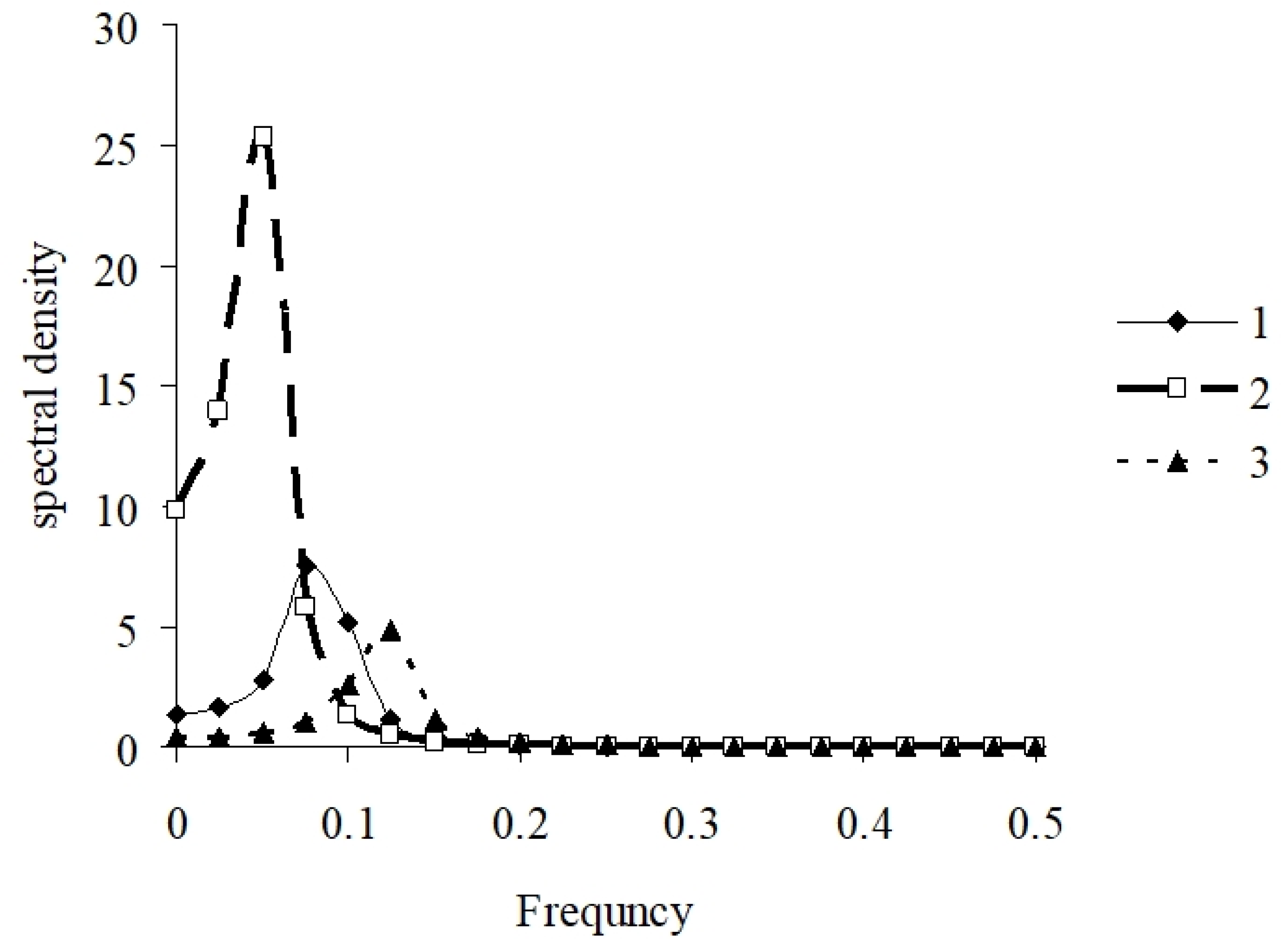
2.5. Data Processing
3. Results
3.1. Model and Model Coefficients
3.2. Model Performance
3.3. Estimation of Wind Damage and Temperature Effect on Tree Mortality Caused by Bark Beetle
4. Discussion
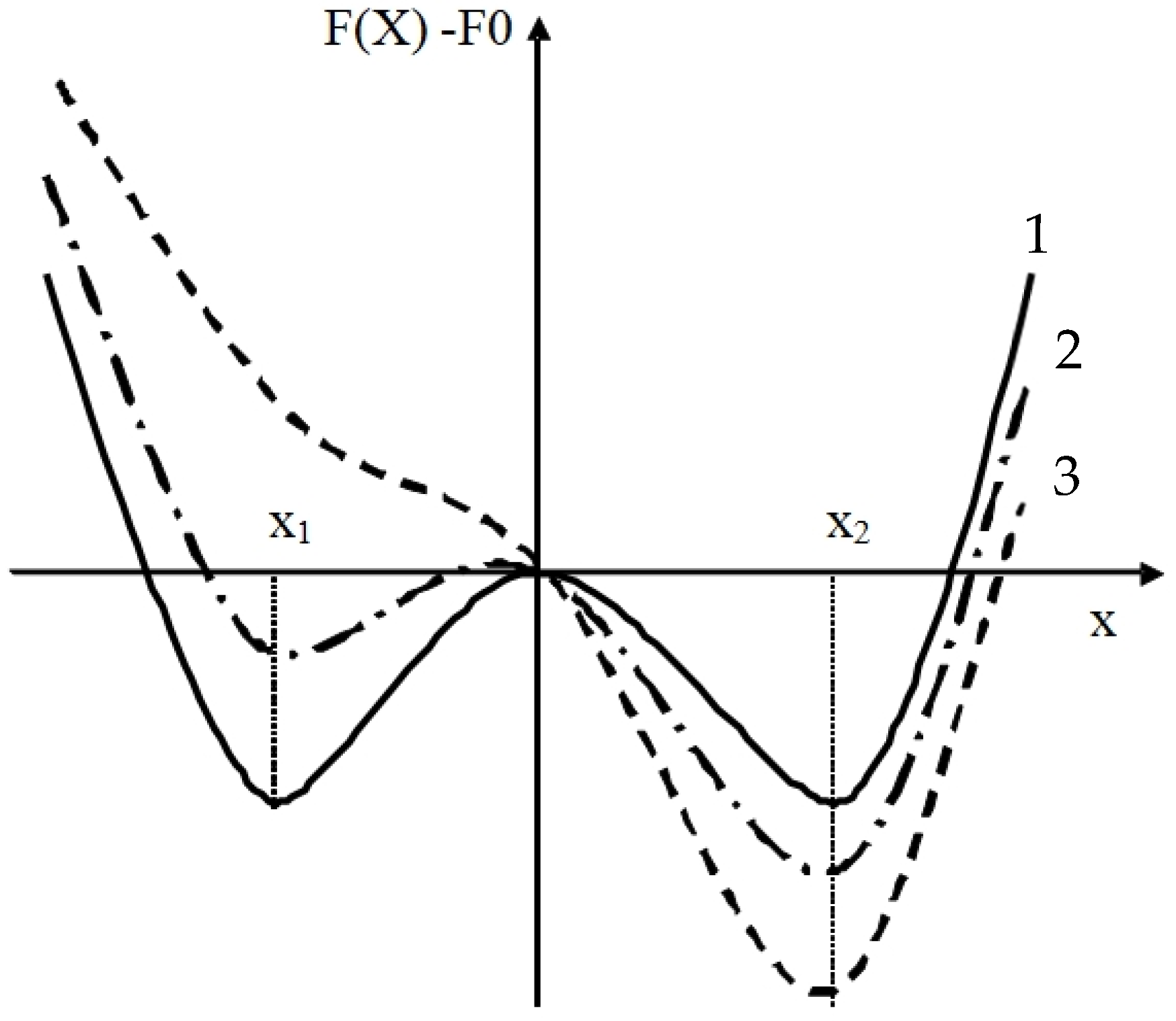
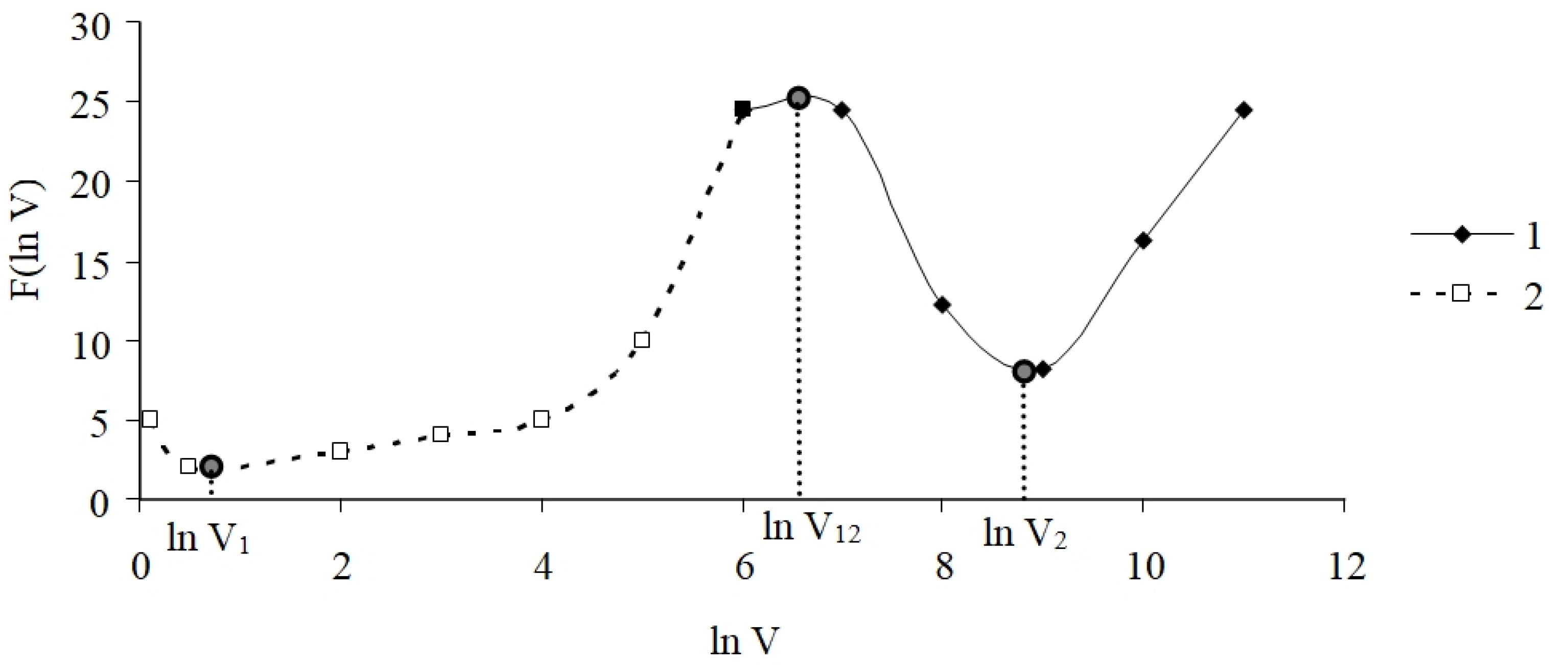
4.1. Model
4.2. Model Coefficients: Environmental and Internal Factors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isaev, A.S.; Girs, G.I. Tree-Xylophagous Insect Interactions; Nauka: Novosibirsk, Russia, 1975. [Google Scholar]
- Christiansen, E.; Bakke, A. The Spruce Bark Beetle of Eurasia. In Dynamics of Forest Insect Populations; Springer: New York, NY, USA, 1988; pp. 479–503. [Google Scholar]
- Økland, B.; Berryman, A. Resource dynamic plays a key role in regional fluctuations of the spruce bark beetles Ips typographus. Agric. For. Entomol. 2004, 6, 141–146. [Google Scholar] [CrossRef]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Jurc, M.; Perko, M.; Džeroski, S.; Demšar, D.; Hrašovec, B. Spruce bark beetles (Ips typographus, Pityogenes chalcographus, Col.: Scolytidae) in the Dinaric mountain forests of Slovenia: Monitoring and modeling. Ecol. Model. 2006, 194, 219–226. [Google Scholar] [CrossRef]
- Panayotov, M.; Kulakowski, D.; Laranjeiro Dos Santos, L.; Bebi, P. Wind disturbances shape old Norway spruce-dominated forest in Bulgaria. For. Ecol. Manag. 2011, 262, 470–481. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang. Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Marini, L.; Lindelöw, Å.; Jönsson, A.M.; Wulff, S.; Schroeder, L.M. Population dynamics of the spruce bark beetle: A long-term study. Oikos 2013, 122, 1768–1776. [Google Scholar] [CrossRef]
- Mezei, P.; Jakuš, R.; Pennerstorfer, J.; Havašová, M.; Škvarenina, J.; Ferenčík, J.; Slivinský, J.; Bičárová, S.; Bilčík, D.; Blaženec, M.; et al. Storms, temperature maxima and the Eurasian spruce bark beetle Ips typographus—An infernal trio in Norway spruce forests of the Central European High Tatra Mountains. Agric. For. Meteorol. 2017, 242, 85–95. [Google Scholar] [CrossRef]
- de Groot, M.; Diaci, J.; Ogris, N. Forest management history is an important factor in bark beetle outbreaks: Lessons for the future. For. Ecol. Manag. 2019, 433, 467–474. [Google Scholar] [CrossRef]
- Forster, B. Development of the bark beetle situation in the Swiss storm-damage areas. Schweiz. Z. Für Forstwes 1993, 144, 767–776. [Google Scholar]
- Doležal, P.; Sehnal, F. Effects of photoperiod and temperature on the development and diapause of the bark beetle Ips typographus. J. Appl. Entomol. 2007, 131, 165–173. [Google Scholar] [CrossRef]
- Wei, W.W.S. Time Series Analysis: Univariative and Multivariative Methods; Addison-Wesley: Boston, MA, USA, 2006. [Google Scholar]
- Shumway, R.H.; Stoffer, D.S. Time Series Analysis and Its Applications with R Examples; Springer Science Business Media: New York, NY, USA, 2006. [Google Scholar]
- Stock, J.H.; Watson, M.W. Introduction to Econometrics; Addison-Wesley: Boston, MA, USA, 2011. [Google Scholar]
- Havašová, M.; Ferenčík, J.; Jakuš, R. Interactions between windthrow, bark beetles and forest management in the Tatra national parks. For. Ecol. Manag. 2017, 391, 349–361. [Google Scholar] [CrossRef]
- Hlásny, T.; Zimová, S.; Merganičová, K.; Štěpánek, P.; Modlinger, R.; Turčáni, M. Devastating outbreak of bark beetles in the Czech Republic: Drivers, impacts, and management implications. For. Ecol. Manag. 2021, 490, 119075. [Google Scholar] [CrossRef]
- Faccoli, M. Effect of Weather on Ips typographus (Coleoptera Curculionidae) Phenology, Voltinism, and Associated Spruce Mortality in the Southeastern Alps. Environ. Entomol. 2009, 38, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Netherer, S.; Matthews, B.; Katzensteiner, K.; Blackwell, E.; Henschke, P.; Hietz, P.; Pennerstorfer, J.; Rosner, S.; Kikuta, S.; Schume, H.; et al. Do water-limiting conditions predispose Norway spruce to bark beetle attack? New Phytol. 2015, 205, 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Marini, L.; Økland, B.; Jönsson, A.M.; Bentz, B.; Carroll, A.; Forster, B.; Grégoire, J.C.; Hurling, R.; Nageleisen, L.M.; Netherer, S.; et al. Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 2017, 40, 1426–1435. [Google Scholar] [CrossRef]
- Berryman, A.A.; Stenseth, N.C.; Wollkind, D.J. Metastability of forest ecosystems infested by bark beetles. Res. Popul. Ecol. 1984, 26, 13–29. [Google Scholar] [CrossRef]
- Wichmann, L.; Ravn, H.P. The spread of Ips typographus (L.) (Coleoptera, Scolytidae) attacks following heavy windthrow in Denmark, analysed using GIS. For. Ecol. Manag. 2001, 148, 31–39. [Google Scholar] [CrossRef]
- Schroeder, L.M. Tree mortality by the bark beetle Ips typographus (L.) in storm-disturbed stands. Integr. Pest Manag. Rev. 2001, 6, 169–175. [Google Scholar] [CrossRef]
- Schroeder, L.M.; Lindelöw, Å. Attacks on living spruce trees by the bark beetle Ips typographus (Col. Scolytidae) following a storm-felling: A comparison between stands with and without removal of wind-felled trees. Agric. For. Entomol. 2002, 4, 47–56. [Google Scholar] [CrossRef]
- Komonen, A.; Schroeder, L.M.; Weslien, J. Ips typographus population development after a severe storm in a nature reserve in southern Sweden. J. Appl. Entomol. 2011, 135, 132–141. [Google Scholar] [CrossRef]
- Modlinger, R.; Novotný, P. Quantification of time delay between damages caused by windstorms and by Ips typographus. For. J. 2015, 61, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Harding, S.; Bärring, L.; Ravn, H.P. Impact of climate change on the population dynamics of Ips typographus in southern Sweden. Agric. For. Meteorol. 2007, 146, 70–81. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Appelberg, G.; Harding, S.; Bärring, L. Spatio-temporal impact of climate change on the activity and voltinism of the spruce bark beetle, Ips typographus. Glob. Chang. Biol. 2009, 15, 486–499. [Google Scholar] [CrossRef]
- Temperli, C.; Bugmann, H.; Elkin, C. Cross-scale interactions among bark beetles, climate change, and wind disturbances: A landscape modeling approach. Ecol. Monogr. 2013, 83, 383–402. [Google Scholar] [CrossRef]
- Křivan, V.; Lewis, M.; Bentz, B.J.; Bewick, S.; Lenhart, S.M.; Liebhold, A. A dynamical model for bark beetle outbreaks. J. Theor. Biol. 2016, 407, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Berryman, A.; Turchin, P. Identifying the density-dependent structure underlying ecological time series. Oikos 2001, 92, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Økland, B.; Bjørnstad, O.N. A resource-depletion model of forest insect outbreaks. Ecology 2006, 87, 283–290. [Google Scholar] [CrossRef]
- Faccoli, M.; Stergulc, F. A practical method for predicting the short-time trend of bivoltine populations of Ips typographus (L.) (Col., Scolytidae). J. Appl. Entomol. 2006, 130, 61–66. [Google Scholar] [CrossRef]
- Lange, H.; Økland, B.; Krokene, P. Thresholds in the life cycle of the spruce bark beetle under climate change. Interj. Complex Syst. 2006, 1648, 1–10. [Google Scholar]
- Baier, P.; Pennerstorfer, J.; Schopf, A. PHENIPS-A comprehensive phenology model of Ips typographus (L.) (Col., Scolytinae) as a tool for hazard rating of bark beetle infestation. For. Ecol. Manag. 2007, 249, 171–186. [Google Scholar] [CrossRef]
- Lewis, M.A.; Nelson, W.; Xu, C. A structured threshold model for mountain pine beetle outbreak. Bull. Math. Biol. 2010, 72, 565–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berec, L.; Doležal, P.; Hais, M. Population dynamics of Ips typographus in the Bohemian Forest (Czech Republic): Validation of the phenology model PHENIPS and impacts of climate change. For. Ecol. Manag. 2013, 292, 1–9. [Google Scholar] [CrossRef]
- Duračiová, R.; Muňko, M.; Barka, I.; Koreň, M.; Resnerová, K.; Holuša, J.; Blaženec, M.; Potterf, M.; Jakuš, R. A bark beetle infestation predictive model based on satellite data in the frame of decision support system tanabbo. IForest 2020, 13, 215–223. [Google Scholar] [CrossRef]
- Kärvemo, S.; Van Boeckel, T.P.; Gilbert, M.; Grégoire, J.C.; Schroeder, M. Large-scale risk mapping of an eruptive bark beetle—Importance of forest susceptibility and beetle pressure. For. Ecol. Manag. 2014, 318, 158–166. [Google Scholar] [CrossRef] [Green Version]
- Kausrud, K.L.; Grégoire, J.C.; Skarpaas, O.; Erbilgin, N.; Gilbert, M.; Økland, B.; Stenseth, N.C. Trees wanted-dead or alive! host selection and population dynamics in tree-killing bark beetles. PLoS ONE 2011, 6, e18274. [Google Scholar] [CrossRef] [Green Version]
- Toffin, E.; Gabriel, E.; Louis, M.; Deneubourg, J.L.; Grégoire, J.C. Colonization of weakened trees by mass-attacking bark beetles: No penalty for pioneers, scattered initial distributions and final regular patterns. R. Soc. Open Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Dobor, L.; Hlásny, T.; Rammer, W.; Zimová, S.; Barka, I.; Seidl, R. Is salvage logging effectively dampening bark beetle outbreaks and preserving forest carbon stocks? J. Appl. Ecol. 2020, 57, 67–76. [Google Scholar] [CrossRef]
- Isaev, A.; Khlebopros, R. The principle of stability in the forest insects population dynamics. Rep. USSR Acad. Sci. 1973, 225–228. [Google Scholar]
- Berryman, A.A. Towards a Unified Theory of Plant Defense. In Mechanisms of Woody Plant Defenses Against Insects; Springer: New York, NY, USA, 1988; pp. 39–55. [Google Scholar]
- Isaev, A.S.; Soukhovolsky, V.G.; Tarasova, O.V.; Palnikova, E.N.; Kovalev, A.V. Forest Insect Population Dynamics, Outbreaks, and Global Warming Effects; John Wiley & Sons, Inc.: New York, NY, USA, 2017. [Google Scholar]
- Netherer, S. Modelling of Bark Beetle Development and of Site- and Stand-Related Predisposition to Ips typographus (L.) (Coleoptera; Scolytidae)—A Contribution to Risk Assessment. Ph.D. Thesis, University of Natural Resources and Life Sciences, Vienna, Austria, 2003. [Google Scholar]
- Jakuš, R. A method for the protection of spruce stands againstIps typographus by the use of barriers of pheromone traps in north-eastern Slovakia. Anz. Schädlingskd. Pflanzenschutz Umweltschutz 1998, 71, 152–158. [Google Scholar] [CrossRef]
- Müller, J.; Bußler, H.; Goßner, M.; Rettelbach, T.; Duelli, P. The European spruce bark beetle Ips typographus in a national park: From pest to keystone species. Biodivers. Conserv. 2008, 17, 2979–3001. [Google Scholar] [CrossRef]
- Svoboda, M.; Janda, P.; Nagel, T.A.; Fraver, S.; Rejzek, J.; Bače, R. Disturbance history of an old-growth sub-alpine Picea abies stand in the Bohemian Forest, Czech Republic. J. Veg. Sci. 2012, 23, 86–97. [Google Scholar] [CrossRef]
- Křenová, Z.; Hruška, J. Proper zonation—An essential tool for the future conservation of the Šumava National Park. Eur. J. Environ. Sci. 2012, 2, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Křenová, Z.; Vrba, J. Just how many obstacles are there to creating a National Park? A case study from the Šumava National Park. Eur. J. Environ. Sci. 2014, 4, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Kindlmann, P.; Krenová, Z. Biodiversity: Protect Czech park from development. Nature 2016, 531, 448. [Google Scholar] [CrossRef] [Green Version]
- Tolasz, R.; Míková, T.; Valeriánová, A. Atlas Podnebí Česka; ČHMÚ, UPOL: Prague, Czech Republic, 2007. [Google Scholar]
- Desai, R.; Kapral, R. Dynamics of Self-Organized and Self-Assembled Structures; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Landau, L. On the theory of phase transitions. JETP 1937, 7, 19–32. [Google Scholar] [CrossRef]
- Van Kampen, N.G. Stochastic Processes in Physics and Chemistry, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Bartholomew, D.J. Time Series Analysis: Forecasting and Control. Oper. Res. Q. 1971, 22, 199–201. [Google Scholar] [CrossRef]
- Kendall, M.; Stuart, A. The Advanced Theory of Statistics; Griffin: London, UK, 1973. [Google Scholar]
- Anderson, T. The Statistical Analysis of Time Series; John Wiley & Sons, Inc.: New York, NY, USA, 1971. [Google Scholar]
- Box, G.E.P.; Jenkins, G.M.; Reinsel, G.C.; Ljung, G.M. Time Series Analysis: Forecasting and Control, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Brockwell, P.J.; Davis, R.A. Introduction to Time Series and Forecasting; Springer Texts in Statistics; Springer: Berlin, Germany, 2016; ISBN 978-3-319-29852-8. [Google Scholar]
- Hamming, R. Digital Filters; Prentice-Hall: New York, NY, USA, 1998. [Google Scholar]
- Jenkins, G.M.; Watts, D.G. Spectral Analysis and Its Applications.; Holden-Day: San Francisco, CA, USA, 1968. [Google Scholar]
- Marple, S.L. Digital Spectral Analysis: With Applications; Prentice-Hall: Hoboken, NJ, USA, 1987. [Google Scholar]
- Dorf, R.C.; Bishop, R.H. Modern Control Systems; Prentice-Hall: Hoboken, NJ, USA, 2001. [Google Scholar]
- Kim, D.P. Control Theory; Fizmathlit: Moscow, Russia, 2007. [Google Scholar]
- Gaiduk, A.P.; Belyaev, V.E.; Pyavchenko, T.A. Control Theory in Examples and Problems; Lan.: St. Petersburg, Russia, 2001. [Google Scholar]
- Stolina, M. The “indifference” of Ips typographus. Zborn. Ved. Pract. Lesn. Fak. VSLD Zvolen 1970, 12, 61–76. [Google Scholar]
- Zumr, V. Biologie a Ekologie Lýkožrouta Smrkového (Ips typographus) a Ochrana Proti Němu; Academia: San Francisco, CA, USA, 1985. [Google Scholar]
- Čermák, P.; Mikita, T.; Trnka, M.; Štšpáneke, P.; Jurečka, F.; Kusbach, A.; Šebesta, J.A.N. Changes in climate characteristics of forest altitudinal zones within the Czech Republic and their possible consequences for forest species composition. Balt. For. 2018, 24, 234–248. [Google Scholar]
- Matthews, B.; Netherer, S.; Katzensteiner, K.; Pennerstorfer, J.; Blackwell, E.; Henschke, P.; Hietz, P.; Rosner, S.; Jansson, P.E.; Schume, H.; et al. Transpiration deficits increase host susceptibility to bark beetle attack: Experimental observations and practical outcomes for Ips typographus hazard assessment. Agric. For. Meteorol. 2018, 263, 69–89. [Google Scholar] [CrossRef]
- Jakuš, R. Bark beetle (Col., Scolytidae) communities and host and site factors on tree level in Norway spruce primeval natural forest. J. Appl. Entomol. 1995, 119, 643–651. [Google Scholar] [CrossRef]
- Royama, T. Analytical Population Dynamics; Chapman and Hall: London, UK, 1992; 371p. [Google Scholar]
- Royama, T.; MacKinnon, W.E.; Kettela, E.G.; Carter, N.E.; Hartling, L.K. Analysis of spruce budworm outbreak cycles in New Brunswick, Canada, since 1952. Ecology 2005, 86, 1212–1224. [Google Scholar] [CrossRef] [Green Version]
- Turchin, P. Rarity of density dependence or population regulation with lags? Nature 1990, 344, 660–663. [Google Scholar] [CrossRef]
- Turchin, P.; Taylor, A.D. Complex dynamics in ecological time series. Ecology 1992, 73, 289–305. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SA Name | SA Coordinates | SA Elevation(m) | SA (ha) | Nearest MS (# id) | MS Coordinates | MS Elevation (m) | Distance between SA and MS (km) |
|---|---|---|---|---|---|---|---|
| Tatra National Park | 49.17° 20.24° | 980–1900 | 2980 | Tatranská Javorina | 49.26° 20.14° | 1013 | MS is located inside of SA |
| Šumava National Park | 48.77° 13.85° | 700–1378 | 68,064 | Churánov (11,457) | 49.07° 13.62° | 1118 | MS is located inside of SA |
| Karlovy Vary division | 50.26° 13.14° | 500–934 | 19,022 | Karlovy Vary airport | 50.20° 12.91° | 603 | 10 |
| Horní Planá division | 48.77° 14.03° | 700–1236 | 19,960 | Černána Podšumaví | 48.74° 14.11° | 740 | MS inside study area |
| Lipník nadBečvou division | 49.63° 17.54° | 500–706 | 27,118 | Červená u Libavé (11,766) | 49.78° 17.54° | 748 | 16 |
| Hořovice division | 49.83° 13.90° | 600–865 | 29,346 | Červená (11,766) | 49.80° 13.75° | 600 | 25 |
| Variables | Coefficients | Std.Err. | t-Test | p-Value |
|---|---|---|---|---|
| a0 | 2.212 | 2.713 | 0.815 | 0.438 |
| T(Mai, t) | 0.231 | 0.097 | 2.386 | 0.044 |
| ln W(t − 1) | −0.382 | 0.316 | −1.208 | 0.262 |
| Δ ln V(t − 2) | −0.475 | 0.266 | −1.786 | 0.112 |
| Δ ln V(t − 1) | 1.344 | 0.224 | 5.987 | 0.000 |
| R2 | 0.910 | |||
| adj.R2 | 0.87 | |||
| F-test | 19.350 | |||
| Variable | Coefficient | Std. Error | t-Test | p-Value |
|---|---|---|---|---|
| Tatra National Park | ||||
| a0 | −1.708 | 0.765 | −2.233 | 0.039 |
| T(Mai,t) | 0.120 | 0.056 | 2.144 | 0.047 |
| ln W(t) | 0.045 | 0.035 | 1.279 | 0.218 |
| Δ ln V(t − 2) | −0.801 | 0.128 | −6.262 | <0.001 |
| Δ ln V(t − 1) | 1.533 | 0.131 | 11.713 | 0.000 |
| R2 | 0.911 | |||
| adj.R2 | 0.88 | |||
| F-test | 43.66 | |||
| Šumava National Park | ||||
| a0 | 3.367 | 1.031 | 3.267 | 0.003 |
| T(Mai, t − 2) | 0.313 | 0.162 | 1.935 | 0.065 |
| ln W(t − 3) | 0.305 | 0.123 | 2.469 | 0.021 |
| Δ ln V(t − 2) | −1.242 | 0.219 | −5.660 | <0.001 |
| Δ ln V(t − 1) | 1.599 | 0.164 | 9.722 | <0.001 |
| R2 | 0.88 | |||
| adj.R2 | 0.84 | |||
| F-test | 40.87 | |||
| Horní Planá division | ||||
| a0 | −3.019 | 2.424 | −1.245 | 0.248 |
| T(Mai, t) | 0.192 | 0.116 | 1.650 | 0.138 |
| ln W(t) | 0.386 | 0.152 | 2.545 | 0.034 |
| ln V(t − 2) | −0.409 | 0.152 | −2.696 | 0.027 |
| ln V(t − 1) | 1.109 | 0.192 | 5.788 | 0.000 |
| R2 | 0.872 | |||
| adj.R2 | 0.78 | |||
| F-test | 13.580 | |||
| Lipník nad Bečvou | ||||
| a0 | −6.086 | 1.460 | −4.170 | 0.004 |
| T(April,t) | 0.047 | 0.037 | 1.277 | 0.242 |
| ln W(t) | 0.478 | 0.117 | 4.105 | 0.005 |
| Δ ln V(t − 2) | −0.717 | 0.124 | −5.757 | 0.001 |
| Δ ln V(t − 1) | 0.384 | 0.143 | 2.676 | 0.032 |
| R2 | 0.911 | |||
| adj.R2 | 0.87 | |||
| F-test | 17.84 | |||
| Hořovice division | ||||
| a0 | −3.047 | 0.933 | −3.266 | 0.014 |
| T(Mai, t) | 0.048 | 0.042 | 1.146 | 0.289 |
| ln W(t) | 0.230 | 0.082 | 2.806 | 0.026 |
| ln V(t − 2) | −0.373 | 0.189 | −1.968 | 0.090 |
| ln V(t − 1) | 0.979 | 0.171 | 5.730 | 0.0007 |
| R2 | 0.88 | |||
| adj.R2 | 0.79 | |||
| F-test | 12.25 | |||
| Parameters and Conditions | Study Area | |||||
|---|---|---|---|---|---|---|
| Karlovy Vary Division | Tatra NP | Šumava NP | Horní Planá Division | Lipník nadBečvou Division | Hořovice Division | |
| a2 | −0.48 | −0.80 | −1.24 | −0.41 | −0.72 | −0.375 |
| a1 | 1.34 | 1.53 | 1.60 | 1.11 | 0.38 | 0.98 |
| a1 + a2 < 1 | 0.26 < 1 | 0.73 < 1 | 0.36 < 1 | 0.70 < 1 | −0.33 < 1 | 0.61 < 1 |
| a1 − a2 > −1 | 1.82 > −1 | 2.33 > −1 | 2.84 > −1 | 1.52 > −1 | 1.10 > −1 | 1.35 > −1 |
| −1 < a2 < 1 | ||||||
| η | 0.13 | 0.10 | 0.17 | 0.29 | 0.28 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soukhovolsky, V.; Kovalev, A.; Tarasova, O.; Modlinger, R.; Křenová, Z.; Mezei, P.; Škvarenina, J.; Rožnovský, J.; Korolyova, N.; Majdák, A.; et al. Wind Damage and Temperature Effect on Tree Mortality Caused by Ips typographus L.: Phase Transition Model. Forests 2022, 13, 180. https://doi.org/10.3390/f13020180
Soukhovolsky V, Kovalev A, Tarasova O, Modlinger R, Křenová Z, Mezei P, Škvarenina J, Rožnovský J, Korolyova N, Majdák A, et al. Wind Damage and Temperature Effect on Tree Mortality Caused by Ips typographus L.: Phase Transition Model. Forests. 2022; 13(2):180. https://doi.org/10.3390/f13020180
Chicago/Turabian StyleSoukhovolsky, Vladislav, Anton Kovalev, Olga Tarasova, Roman Modlinger, Zdenka Křenová, Pavel Mezei, Jaroslav Škvarenina, Jaroslav Rožnovský, Nataliya Korolyova, Andrej Majdák, and et al. 2022. "Wind Damage and Temperature Effect on Tree Mortality Caused by Ips typographus L.: Phase Transition Model" Forests 13, no. 2: 180. https://doi.org/10.3390/f13020180