
Diurnal Tree Stem CH4 and N2O Flux Dynamics from a Riparian Alder Forest
by
 , ,
, ,
Thomas Schindler
1,2,* ,
,
Katerina Machacova
1,2,
Ülo Mander
1,2 ,
,
Jordi Escuer-Gatius
3 and
Kaido Soosaar
1,2 1
Department of Geography, Institute of Ecology & Earth Sciences, University of Tartu, Vanemuise 46, 51014 Tartu, Estonia
2
Department of Ecosystem Trace Gas Exchange, Global Change Research Institute of the Czech Academy of Sciences, Belidla 4a, 60300 Brno, Czech Republic
3
Institute of Agricultural and Environmental Sciences, Estonian University of Life Sciences, Fr. R. Kreutzwaldi 1, 51006 Tartu, Estonia
*
Author to whom correspondence should be addressed.
Forests 2021, 12(7), 863; https://doi.org/10.3390/f12070863
Submission received: 21 April 2021
/
Revised: 14 June 2021
/
Accepted: 28 June 2021
/
Published: 30 June 2021
{kind=link}
{kind=link}
Abstract
:Tree stems play an important role in forest methane (CH4) and nitrous oxide (N2O) flux dynamics. Our paper aimed to determine the unknown diurnal variability of CH4 and N2O exchange in grey alder tree stems. The gas fluxes in tree stems and adjacent soil were measured using manual static and dynamic chamber systems with gas chromatographic and laser-spectroscopic analysis, respectively. The alder trees were predominant emitters of CH4 and N2O; however, N2O emission from stems was negligible. The soil mainly emitted N2O into the atmosphere and was both a source and sink of CH4, depending on environmental conditions. Neither the tree stems nor the riparian forest soil showed significant differences in their CH4 and N2O fluxes between the daytime and nighttime, independently of the exchange rates. In contrast to several previous studies revealing a diurnal variability of greenhouse gas fluxes from tree stems, our investigation did not show any clear daytime–nighttime differences. On the other hand, we found quite clear seasonal dynamics initiated by changing environmental conditions, such as temperature and soil water conditions and tree physiological activity. Our results imply a transport role of tree stems for soil-produced CH4 and N2O rather than the production of these gases in tree tissues, even though this cannot be excluded.
1. Introduction
Recent studies on methane (CH4) and nitrous oxide (N2O) balance have highlighted the importance of trees, which may act either as sources or sinks for both gases [1,2,3]. Grey alder (Alnus incana (L.) Moench) is a fast-growing, common pioneer tree species in the Northern hemisphere, often found in riparian zones [4,5,6] and with excellent potential for short-rotation forestry [7,8,9]. Wetland plants such as alder adapt to wet conditions by possessing air-filled aerenchyma to supply tissues with oxygen [10] and constitute a pathway for general gas exchange through plants [11,12]. Further, the decomposed litter of nutrient-rich alder forests improves soil properties, alters microbial activities in the soil, and consequentially affects the CH4 and N2O exchange [13,14]. Anaerobic conditions in water-saturated environments enhance the production of CH4 in methanogenesis, whereas N2O can be produced under both aerobic (nitrification) and anaerobic (denitrification) conditions [15,16]. Recent studies have identified possible mechanisms describing the contribution of trees to an ecosystem’s greenhouse gas (GHG) exchange: (i) gas uptake from the soil by tree roots, transport within the tree tissues, and release to the atmosphere [11]; (ii) gas uptake from the atmosphere [17]; (iii) alternation of gas turnover processes in adjacent soils [1,18]; (iv) in situ production of N2O and CH4 in plant tissues [19,20,21]. The factors related to decreasing CH4 and N2O production causing uptake by trees are less studied; however, several studies point out the role of cryptogams covering tree stems, which can be both sources [18,22] or sinks of these gases [3]. Likewise, methanotrophs on tree bark [23] or within the stem [24] can consume CH4, transforming it to CO2. The majority of studies on tree stem CH4 and N2O fluxes have focused on different tree species growing in various forest ecosystems, seasonal flux dynamics, and the effects of environmental conditions on the gas exchange [17,25,26]; however, investigations on diurnal flux dynamics, and specifically on measurements during the nighttime, are still scarce [27,28,29,30] but are necessary in order to understand the temporal patterns, better explain the origin of the exchanged gases, and especially to correctly estimate the daily fluxes of CH4 and N2O from tree stems and forest ecosystems, which are the basis for correct estimation of the monthly and annual GHG fluxes.
As such, our objective was to analyze the diurnal pattern of CH4 and N2O fluxes from grey alder stems. Based on several previous studies on temperate forests, we hypothesized that the diurnal pattern of soil fluxes is not reflected in the stem flux dynamics.
2. Materials and Methods
2.1. Description of the Study Site
Our measurement campaigns were performed based on expected climate characteristics with intensive solar radiation, anticipated seasonal plant activities, and consequential GHG fluxes during three periods (according to the unpublished data by Mander et al., 2021): 20.07–02.09.2017 (hereafter termed “summer 2017”, n = 6), 30.04–26.05.2018 (“spring 2018”, n = 3), and 12.07–06.09.2018 (“summer 2018”, n = 7). The riparian 40-year-old hemi-boreal Filipendula grey alder (Alnus incana (L.) Moench) forest stands on a former agricultural Gleysol in eastern Estonia. Our studied plots covered an area of 60 × 60 m with a slight slope of 1%. The tree stand density was approximately 1500 trees per hectare and the tree height was 19.2 ± 1.4 m. In addition, the tree diameter at 1.3 m height was approximately 0.17 ± 0.03 m [14]. The long-term average annual precipitation in the region is 650 mm, with an average temperature of 17.0 °C in July and −6.7 °C in January. Typically, the growing season lasts 175–180 days from mid-April to October [31].
2.2. Stem and Soil Gas Flux Measurements
For stem gas flux measurements, eleven representative grey alder trees were equipped with pairs of static tree stem chambers, set randomly across 180° and interconnected with tubes into one system, and installed approximately 10 cm above ground level. Four gas samples of 25 mL each were taken from each chamber system in an hourly sequence (0–1–2–3 h) over the periods of 12:00–16:00 (i.e., daytime) and 00:00–04:00 (i.e., nighttime) and analyzed using gas chromatography (GC-2014, Shimadzu, Japan) equipped with an electron capture detector for detection of N2O and a flame ionization detector for CH4 (for more details related to gas chromatographical settings, see [32].
For the investigation of soil gas exchange, opaque automatic dynamic soil chambers covering 0.16 m2 soil surface and containing a volume of 0.032 m3 were installed close to selected trees. A gas analyzer (G2508; Picarro Inc., Santa Clara, CA, USA) monitored CH4 and N2O concentrations in the chambers. Only fluxes measured in the daytime and nighttime periods characterized above were used (for further details of the experimental setup, see [14]).
2.3. Measurements of Environmental Parameters
Close to soil flux chambers and measurement trees, soil moisture (ML3 ThetaProbe; Delta-T Devices Ltd., Cambridge, UK) and soil temperature sensors (CS 107; Campbell Scientific Inc., Logan, UT, USA) at a soil depth of 5 cm and piezometers for the soil water table (Hobo U20L-04; Onset Computer Corporation, Bourne, MA, USA) were installed for the continuous measurements. Air temperature (Rotronic HC2A-S3; Rotronic AG, Bassersdorf, Switzerland) and photosynthetically active radiation (LI-190SL; LI-COR Biosciences, Lincoln, NE, USA) were measured continuously in a micrometeorological tower at heights of 3 m and 21 m from the ground, respectively.
2.4. Data Quality Check and Statistical Analysis
Stem and soil gas fluxes were calculated on a linear approach according to CH4 and N2O concentration changes in the chamber headspace over the measurement period, using the equation proposed by [33]. The R2 of the linear fit of the CO2 measurement was used to indicate the flux stability and quality of the measurement. CH4 and N2O values were accepted, independently of their respective R2 values, if R2 (CO2) > 0.95.
Sigmaplot (version 12, Systat Software, San Jose, CA, USA) software was used for all statistical analyses. The flux data were checked for normal distribution (Shapiro–Wilk test) and equality of variances in the different subpopulations. The nonparametric Mann–Whitney rank sum test, the Spearman rank correlation test for pair comparison, and the Kruskal–Wallis one-way ANOVA on ranks for multiple comparisons were applied to compare the daytime and nighttime fluxes. The significance level was set to 0.05.
The environmental data were normally distributed and we used means ± standard deviation to present the results. Since the CH4 and N2O fluxes from both stems and soils were non-normally distributed, for the graphical presentation, the median values, 25th and 75th percentiles, and 10th and 90th percentiles were used.
3. Results
3.1. Environmental Parameters
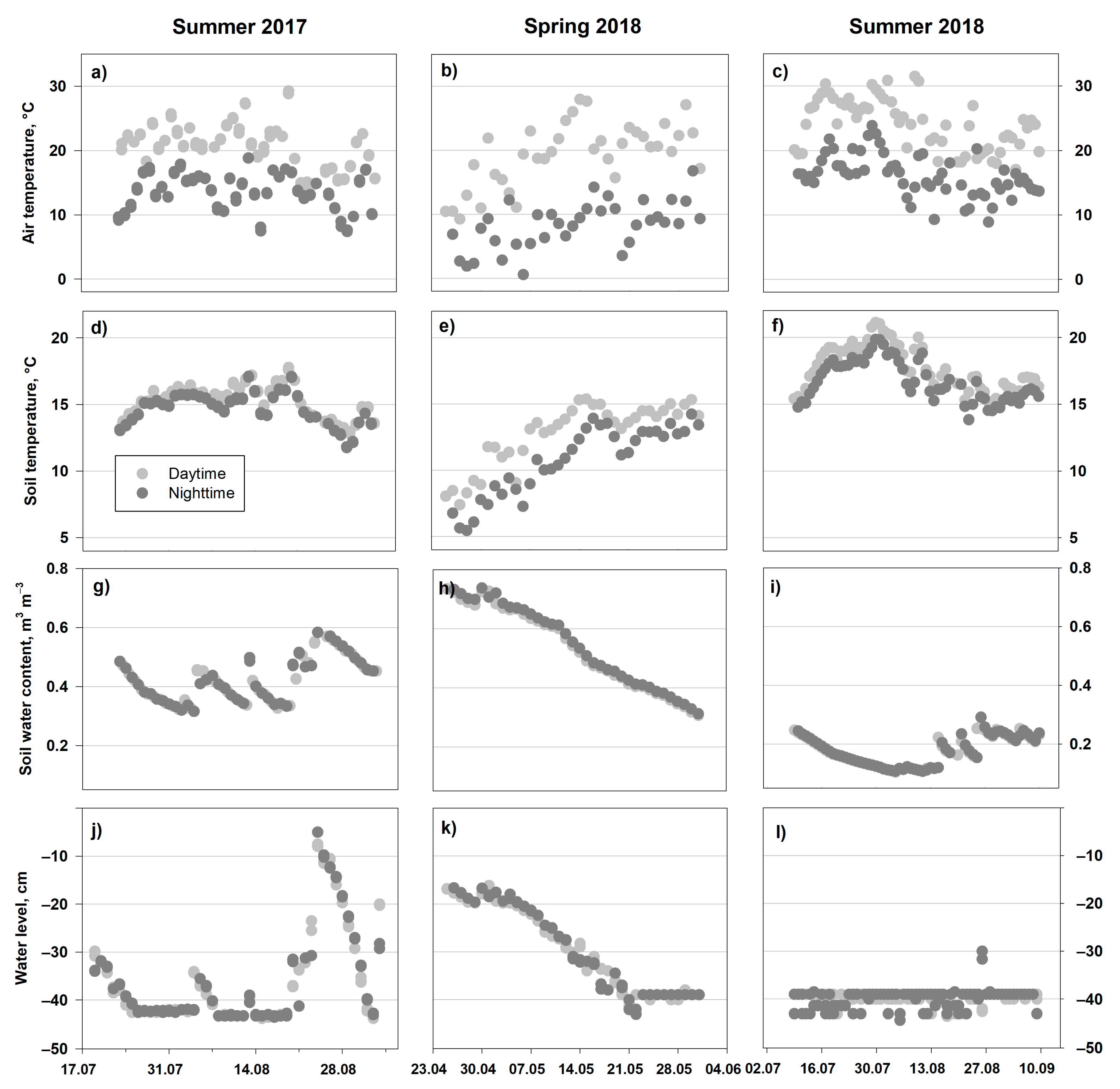
The daytime (noon) mean air temperatures were 20.5 ± 3.4 (mean ± standard deviation) in summer 2017, 20.2 ± 4.1 in spring 2018, and 24.0 ± 4.2 °C in summer 2018, whereas nighttime (midnight) values were 13.7 ± 2.9, 8.1 ± 3.3, and 16.1 ± 3.3 °C, respectively, showing related diurnal trends (Figure 1a–c).
The soil temperatures (Figure 1d–f) were 15.3 ± 1.2, 12.7 ± 2.4, and 17.7 ± 1.7 °C at noon during the three investigated periods. While showing a similar trend, the values at nighttime were 14.7 ± 1.2, 10.7 ± 2.6, and 16.7 ± 1.5 °C.
The photosynthetically active radiation (PAR) during the nighttime was close to zero (from 0.0234 to 0.218 µmol s−1), while daytime values reached 828 ± 385, 1003 ± 308, and 793 ± 420 µmol s−1 during the three study periods, respectively.
There were no significant differences in volumetric soil water content (SWC) between daytime and nighttime. In summer 2017, SWC declined from 0.63 to 0.32 m3 m−3, showing intermediate increases. In spring and summer 2018, SWC declined from 0.74 to 0.38 and from 0.29 to 0.1 m3 m−3, respectively (Figure 1g–i). Further, the soil water table was in good accordance with SWC, underlining the impacts of intense precipitation events on the water table in August 2017 and the declining trend starting in May 2018 (Figure 1j–l).
3.2. Soil and Stem CH4 Fluxes
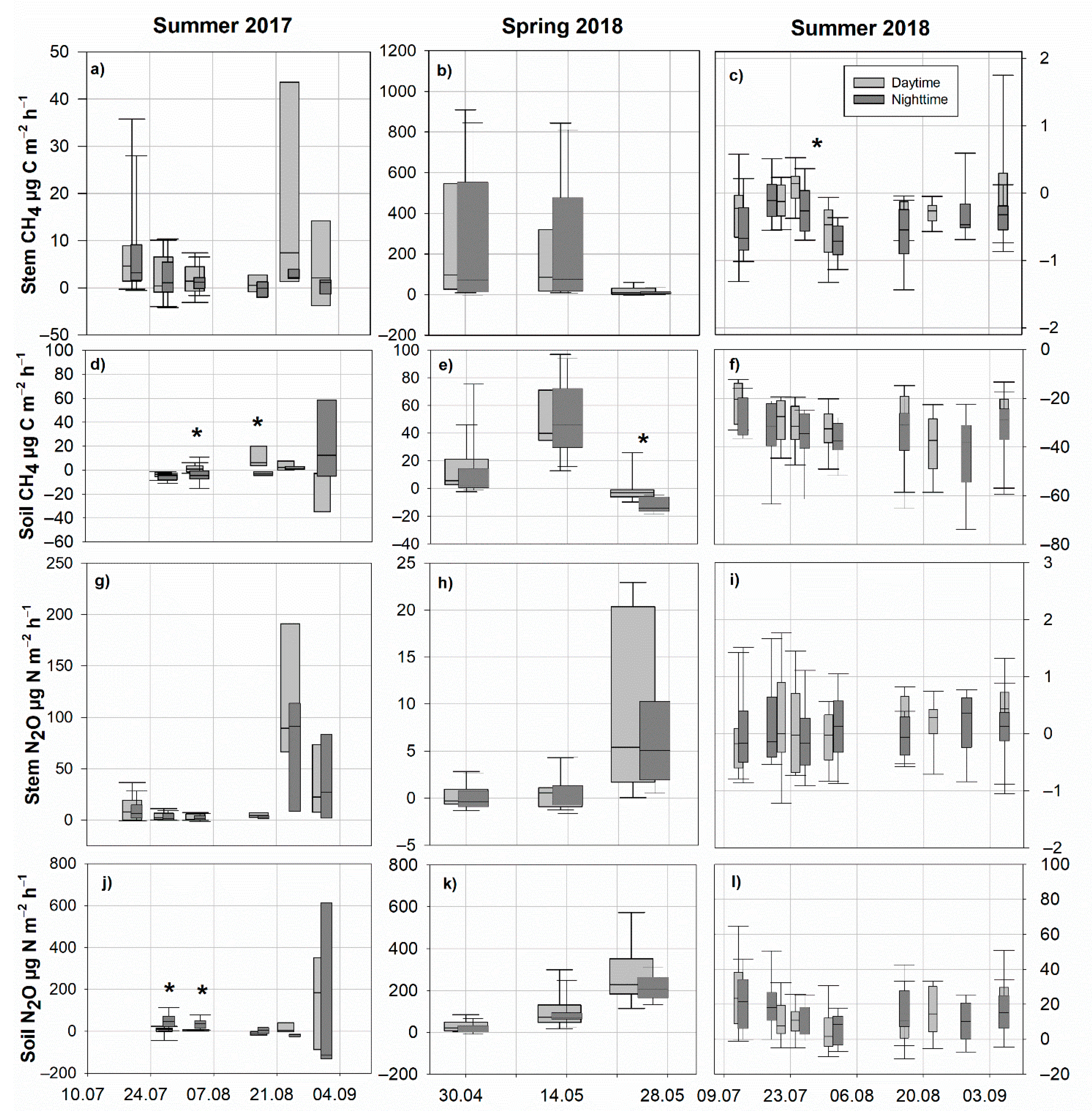
Measured tree stems were emitters of CH4 with low flux rates (mean ± standard error) of 4.63 ± 1.42 µg C m−2 h−1 during daytime and 2.96 ± 0.88 µg C m−2 h−1 during nighttime in summer 2017, which were substantially higher in spring 2018 (164.1 ± 33.92 µg C m−2 h−1, daytime) versus May 2018 (168.47 ± 33.59 µg C m−2 h−1, nighttime), when the SWC was the highest. In contrast, stems had fluctuating CH4 uptake rates of −0.21 ± 0.06 (daytime) and −0.40 ± 0.05 (nighttime) µg C m−2 h−1 during the very dry summer of 2018 (Figure 2a–c).
The soil CH4 exchange varied seasonally. Predominant CH4 uptake was observed in summer 2017 and 2018, both during the day- and nighttime; however, the predominant CH4 emissions were detected in spring 2018 (Figure 2d–f), strongly connected to high SWC and the related water table. Despite single campaigns, no statistically significant differences between day and nighttime soil fluxes were observed.
3.3. Soil and Stem N2O Fluxes
Tree stems were net emitters of N2O during all measurement periods (Figure 2g–i): summer 2017, 14.93 ± 4.90 µg N m−2 h−1 at daytime and 11.17 ± 3.56 µg N m−2 h−1 at nighttime; in spring 2018, 3.30 ± 0.83 µg N m−2 h−1 at daytime and 2.39 ± 0.56 µg N m−2 h−1 at nighttime; summer 2018, 0.11 ± 0.08 µg N m−2 h−1 at daytime and −0.04 ± 0.22 µg N m−2 h−1 at nighttime.
The soil was also dominantly a net emitter of N2O, with the highest fluxes detected in spring 2018 (Figure 2j–l) when the SWC and water table were rapidly decreasing from 0.75 to 0.55 m3 m−3 and from −5 to −30 cm, respectively (unpublished data by Mander et al., 2021). Differences between day- and nighttime were not statistically significant for either stem or soil N2O fluxes.
4. Discussion
Regardless of the tree stem flux direction (gas emission or uptake) and the gas exchange rates, which considerably differed for each of our studied periods, no statistically significant differences between day- and nighttime stem CH4 and N2O fluxes were revealed. Similarly, the soil fluxes of CH4 and N2O mostly did not significantly vary between the day- and nighttime. In contrast to Barba et al. [28] and Pitz and Megonigal [30], who found significant diurnal temporal correlations between these gases with temperature or sap flux intensity, in our study, such relationships could not be found. Similarly, fluctuations of the hydrological regime in forests that initiated diurnal fluctuations of stem methane fluxes [29] could not be observed in our grey alder forest.
Various studies have shown seasonal patterns of stem GHG fluxes [2,17,26,34], initiated by changing environmental conditions. For instance, seasonal changes in air and soil temperature in temperate forests [2], as well as soil water content and related oxygen availability, can initiate regular differences in both CH4 and N2O fluxes from stems [2,34]. Machacova et al. [17] found that the physiological activity of trees is the main factor causing seasonal fluctuations of N2O stem fluxes. To our knowledge, few existing investigations have detected a high diurnal flux variability [2,29,30] while being reported as not consistent throughout the growing season [28]. Individual trees, as investigated before, may reflect a dynamic gas exchange during the daytime, although such diurnal variability could not be found in our 12 h time-shifted experiment with 11 trees.
Methane in soils is produced under anoxic conditions, and known controlling factors are temperature and SWC [35,36]. Unlike previous diurnal studies on CH4 emissions in different soils [21,35,37], we could not observe a significant difference within our studied periods. However, moderate diurnal changes in soil temperature and SWC were detected. Even the observed impact of heavy precipitation on the soil water table showed no significant effect on the diurnal CH4 flux ratio, indicating that the anaerobicity within the soils was less affected in short terms; however, the dependency of fluxes on the availability of water and temperatures detected in our study confirmed the seasonal trends in CH4 fluxes, as also described in the literature for both soils [38] and stems [2].
The initial positive correlation of PAR, photosynthesis rates, and CH4 emissions [39] in vascular plants was updated by Joabsson et al. [40], associating the interactions of belowground ventilation and soil microbial processes. The hypothesized independence of tree stem CH4 fluxes on tree physiological activity, such as photosynthesis rates expressed by PAR measurements, was confirmed, since our study did not show any fast response of CH4 emissions to increased solar radiation within the 12 h time slots; however, wetland tree species such as Alnus trees are assumed to preferably passively transport soil-produced CH4 via gas diffusion through the aerenchyma system and exchange it with the atmosphere later on [11,41].
Nitrous oxide is produced under aerobic (nitrification) or anaerobic (denitrification) conditions [15]; hence, soil water content influences both of these processes in soil [42,43]. Investigations on the changes of soil water conditions and their impacts on N2O fluxes have revealed the importance of a specific range of SWC rather than a temporal dependence [44]. In compliance with these findings, our study confirms that diurnal differences of SWC and soil temperature are minor and that they do not affect N2O fluxes from either tree stems or soils; however, similarly to the recently reported study by Machacova et al. [17], we found that seasonally changing soil water regimes and soil temperatures influence N2O fluxes from soil and stems. Further, we observed inter- and intraseasonal influences of low SWC levels on N2O emissions. A similar impact was mentioned by Barrat et al. [45]. Further analysis of the anaerobicity, substrate conditions, and microbial communities are needed to explain the temporal dynamics of N2O fluxes from stems.
Deciduous trees such as ash (Fraxinus excelsior L.) and beech (Fagus sylvatica L.) may emit less N2O under laboratory-simulated light conditions [46]. Further, Machacova et al. [17] found a positive, interseasonal correlation of stem N2O fluxes with air temperature and PAR. Likewise, our results indicate possible interseasonal dynamics, particularly regarding the summer 2017 and spring 2018 results.
Regarding the sources of CH4 and N2O stem emissions and their diurnal pattern, Megonigal et al. [47], using radon as a natural tracer of gas transport through trees, found that tree stems emit soil gases during the night when transpiration rates are negligible. This study also suggests that axial diffusion is an important plant transport of soil gases controlled by plant hydraulics, whether by diffusion or mass flow via transpiration [47]. Future tracer investigations may, therefore, underline the transport process theory. An additional factor potentially regulating N2O fluxes from tree stems is their coverage with cryptogamic stem covers (i.e., photoautotrophic organisms living on bark such as lichens, algae, and mosses), which might also exchange N2O and CH4 with the atmosphere [18]. In accordance with the findings of Lenhart et al. [22], we did not observe PAR-dependent differences in fluxes in our diurnal study; however, the previous study in our ecosystem revealed a substantial effect of SWC and nitrogen availability on fluxes from stems and soil [14]. As such, a relevant impact of cryptogams on N2O fluxes from stems seems unlikely, although it cannot be excluded and may be part of future investigations.
Our results suggest a transport role of tree stems for soil-produced CH4 and N2O rather than a production of CH4 and N2O in tree tissues, even if this cannot be excluded. Based on our results detecting no differences in tree stem fluxes during daytime or nighttime, the tree stem CH4 and N2O fluxes commonly measured in the daytime can be used alone to estimate the daily, monthly, and annual fluxes in riparian alder forests; however, further detailed research and modeling approaches could provide deeper insights into this topic and contribute to the solution.
5. Conclusions
Our hypothesis on diurnal changes in CH4 and N2O soil and stem fluxes was partly supported; we did not find significant diurnal dynamics of CH4 and N2O fluxes from alder stems. However, the soil fluxes unexpectedly did not follow any regular diurnal pattern. On the other hand, our study revealed a seasonal pattern of CH4 and N2O fluxes from tree stems. Although we could not find significant diurnal dynamics for either stems or soil fluxes, the findings still indicated that the tree stems of grey alder most likely act as passive transport systems of gases from the soil, even though production of these gases in tree tissues cannot be excluded. The soil was a significant emitter of N2O into the atmosphere, whereas CH4 was either emitted or consumed in soil, mainly depending on the soil water content. Our survey underlines that it is important to analyze GHG fluxes in this complex dynamic system to better understand carbon and nitrogen cycles in forest ecosystems.
Author Contributions
Conceptualization, T.S.; methodology, T.S., K.M. and K.S.; software, T.S. and J.E.-G.; resources, Ü.M., K.M. and K.S.; writing—original draft preparation, T.S.; writing—review and editing, Ü.M., K.M., J.E.-G. and K.S.; visualization, T.S.; supervision, Ü.M., K.M. and K.S.; funding acquisition, Ü.M., K.M. and K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Estonian Research Council (IUT2-16, PRG-352, and MOBERC20); the Czech Science Foundation (17-18112Y); the Ministry of Education, Youth, and Sports of Czech Republic within the National Sustainability Program I (NPU I, grant number LO1415); project SustES—Adaptation Strategies for Sustainable Ecosystem Services and Food Security Under Adverse Environmental Conditions (CZ.02.1.01/0.0/0.0/16_019/0000797); the European Commission through the European Regional Development Fund (the Center of Excellence EcolChange), LIFE OrgBalt project “Demonstration of Climate Change Mitigation Potential of Nutrient-Rich Organic Soils in Baltic States and Finland” (LIFE18 CCM/LV/001158); and the European Social Fund (Doctoral School of Earth Sciences and Ecology).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Marek Jakubik for his technical support by stem chamber installation.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Maier, M.; Machacova, K.; Lang, F.; Svobodova, K.; Urban, O. Combining soil and tree-stem flux measurements and soil gas profiles to understand CH4 pathways in Fagus sylvatica forests. J. Plant Nutr. Soil Sci. 2018, 181, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Barba, J.; Bradford, M.A.; Brewer, P.E.; Bruhn, D.; Covey, K.; Van Haren, J.; Megonigal, J.P.; Mikkelsen, T.N.; Pangala, S.R.; Pihlatie, M.; et al. Methane emissions from tree stems: A new frontier in the global carbon cycle. New Phytol. 2019, 222, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Machacova, K.; Borak, L.; Agyei, T.; Schindler, T.; Soosaar, K.; Mander, Ü.; Ah-Peng, C. Trees as net sinks for methane (CH4) and nitrous oxide (N2O) in the lowland tropical rain forest on volcanic Réunion Island. New Phytol. 2021, 229, 1983–1994. [Google Scholar] [CrossRef]
- Uri, V.; Lõhmus, K.; Kiviste, A.; Aosaar, J. The dynamics of biomass production in relation to foliar and root traits in a grey alder (Alnus incana (L.) Moench) plantation on abandoned agricultural land. Forestry 2008, 82, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Rytter, L.; Rytter, R.-M. Growth and carbon capture of grey alder (Alnus incana (L.) Moench.) under north European conditions—Estimates based on reported research. For. Ecol. Manag. 2016, 373, 56–65. [Google Scholar] [CrossRef]
- Uri, V.; Tullus, H.; Lõhmus, K. Biomass production and nutrient accumulation in short-rotation grey alder (Alnus incana (L.) Moench) plantation on abandoned agricultural land. For. Ecol. Manag. 2002, 161, 169–179. [Google Scholar] [CrossRef]
- Uri, V.; Kukumägi, M.; Aosaar, J.; Varik, M.; Becker, H.; Soosaar, K.; Morozov, G.; Ligi, K.; Padari, A.; Ostonen, I.; et al. Carbon budgets in fertile grey alder (Alnus incana (L.) Moench.) stands of different ages. For. Ecol. Manag. 2017, 396, 55–67. [Google Scholar] [CrossRef]
- Uri, V.; Lõhmus, K.; Mander, Ü.; Ostonen, I.; Aosaar, J.; Maddison, M.; Helmisaari, H.-S.; Augustin, J. Long-term effects on the nitrogen budget of a short-rotation grey alder (Alnus incana (L.) Moench) forest on abandoned agricultural land. Ecol. Eng. 2011, 37, 920–930. [Google Scholar] [CrossRef]
- Aosaar, J.; Varik, M.; Lõhmus, K.; Ostonen, I.; Becker, H.; Uri, V. Long-term study of above- and below-ground biomass production in relation to nitrogen and carbon accumulation dynamics in a grey alder (Alnus incana (L.) Moench) plantation on former agricultural land. Eur. J. For. Res. 2013, 132, 737–749. [Google Scholar] [CrossRef]
- Sorz, J.; Hietz, P. Gas diffusion through wood: Implications for oxygen supply. Trees Struct. Funct. 2006, 20, 34–41. [Google Scholar] [CrossRef]
- Rusch, H.; Rennenberg, H. Black alder (Alnus Glutinosa (L.) Gaertn.) trees mediate methane and nitrous oxide emission from the soil to the atmosphere. Plant Soil 1998, 201, 1–7. [Google Scholar] [CrossRef]
- Schröder, P. Aeration of the root system in Alnus glutinosa L. Gaertn. Ann. Sci. For. 1989, 46, 310s–314s. [Google Scholar] [CrossRef]
- Huth, V.; Hoffmann, M.; Bereswill, S.; Popova, Y.; Zak, D.; Augustin, J. The climate warming effect of a fen peat meadow with fluctuating water table is reduced by young alder trees. Mires Peat 2018, 21, 1–18. [Google Scholar] [CrossRef]
- Schindler, T.; Mander, Ü.; Machacova, K.; Espenberg, M.; Krasnov, D.; Escuer-Gatius, J.; Veber, G.; Pärn, J.; Soosaar, K. Short-term flooding increases CH4 and N2O emissions from trees in a riparian forest soil-stem continuum. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Butterbach-Bahl, K.; Baggs, L.; Dannenmann, M.; Kiese, R.; Zechmeister-Boltenstern, S. Nitrous oxide emissions from soils: How well do we understand the processes and their controls? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130122. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.A.; Ball, T.; Conen, F.; Dobbie, K.E.; Massheder, J.; Rey, A. Exchange of greenhouse gases between soil and atmosphere: Interactions of soil physical factors and biological processes. Eur. J. Soil Sci. 2018, 69, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Machacova, K.; Vainio, E.; Urban, O.; Pihlatie, M. Seasonal dynamics of stem N2O exchange follow the physiological activity of boreal trees. Nat. Commun. 2019, 10, 4989. [Google Scholar] [CrossRef]
- Machacova, K.; Maier, M.; Svobodova, K.; Lang, F.; Urban, O. Cryptogamic stem covers may contribute to nitrous oxide consumption by mature beech trees. Sci. Rep. 2017, 7, 13243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, D.R.; Bloom, A.J. Wheat leaves emit nitrous oxide during nitrate assimilation. Proc. Natl. Acad. Sci. USA 2001, 98, 7875–7878. [Google Scholar] [CrossRef] [Green Version]
- Keppler, F.; Hamilton, J.T.G.; Brass, M.; Röckmann, T. Methane emissions from terrestrial plants under aerobic conditions. Nature 2006, 439, 187–191. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Nikkel, D.J.; Scherloski, L.M.; Tkach, R.E.; Smits, K.M.; Selinger, L.B.; Rood, S.B. Multiple processes contribute to methane emission in a riparian cottonwood forest ecosystem. New Phytol. 2021, 229, 1970–1982. [Google Scholar] [CrossRef]
- Lenhart, K.; Weber, B.; Elbert, W.; Steinkamp, J.; Clough, T.; Crutzen, P.; Pöschl, U.; Keppler, F. Nitrous oxide and methane emissions from cryptogamic covers. Glob. Chang. Biol. 2015, 21, 3889–3900. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, L.C.; Maher, D.T.; Chiri, E.; Leung, P.M.; Nauer, P.A.; Arndt, S.K.; Tait, D.R.; Greening, C.; Johnston, S.G. Bark-dwelling methanotrophic bacteria decrease methane emissions from trees. Nat. Commun. 2021, 12, 1–8. [Google Scholar] [CrossRef]
- Putkinen, A.; Siljanen, H.M.P.; Laihonen, A.; Paasisalo, I.; Porkka, K.; Tiirola, M.; Haikarainen, I.; Tenhovirta, S.; Pihlatie, M. New insight to the role of microbes in the methane exchange in trees: Evidence from metagenomic sequencing. New Phytol. 2021, 231, 524–536. [Google Scholar] [CrossRef]
- Terazawa, K.; Yamada, K.; Ohno, Y.; Sakata, T.; Ishizuka, S. Spatial and temporal variability in methane emissions from tree stems of Fraxinus mandshurica in a cool-temperate floodplain forest. Biogeochemistry 2015, 123, 349–362. [Google Scholar] [CrossRef]
- Pitz, S.L.; Megonigal, J.P.; Chang, C.-H.; Szlavecz, K. Methane fluxes from tree stems and soils along a habitat gradient. Biogeochemistry 2018, 137, 307–320. [Google Scholar] [CrossRef]
- Pangala, S.R.; Gowing, D.J.; Hornibrook, E.; Gauci, V. Controls on methane emissions from Alnus glutinosa saplings. New Phytol. 2014, 201, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barba, J.; Poyatos, R.; Vargas, R. Automated measurements of greenhouse gases fluxes from tree stems and soils: Magnitudes, patterns and drivers. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Jeffrey, L.C.; Maher, D.T.; Tait, D.R.; Euler, S.; Johnston, S.G. Tree stem methane emissions from subtropical lowland forest (Melaleuca quinquenervia) regulated by local and seasonal hydrology. Biogeochemistry 2020, 151, 273–290. [Google Scholar] [CrossRef]
- Pitz, S.; Megonigal, J.P. Temperate forest methane sink diminished by tree emissions. New Phytol. 2017, 214, 1432–1439. [Google Scholar] [CrossRef] [Green Version]
- Kupper, P.; Sõber, J.; Sellin, A.; Lõhmus, K.; Tullus, A.; Räim, O.; Lubenets, K.; Tulva, I.; Uri, V.; Zobel, M.; et al. An experimental facility for free air humidity manipulation (FAHM) can alter water flux through deciduous tree canopy. Environ. Exp. Bot. 2011, 72, 432–438. [Google Scholar] [CrossRef]
- Soosaar, K.; Mander, Ü.; Maddison, M.; Kanal, A.; Kull, A.; Lõhmus, K.; Truu, J.; Augustin, J. Dynamics of gaseous nitrogen and carbon fluxes in riparian alder forests. Ecol. Eng. 2011, 37, 40–53. [Google Scholar] [CrossRef]
- Livingston, G.P.; Hutchinson, G.L. Enclosure-based measurement of trace gas exchange: Applications and sources of error. In Biogenic Trace Gases: Measuring Emissions from Soil and Water; Matson, P.A., Harris, R.C., Eds.; Blackwell Science Ltd.: Oxford, UK, 1995; pp. 14–51. [Google Scholar]
- Pangala, S.R.; Hornibrook, E.; Gowing, D.J.G.; Gauci, V. The contribution of trees to ecosystem methane emissions in a temperate forested wetland. Glob. Chang. Biol. 2015, 21, 2642–2654. [Google Scholar] [CrossRef] [PubMed]
- Mikkelä, C.; Sundh, I.; Svensson, B.H.; Nilsson, M. Diurnal variation in methane emission in relation to the water table, soil temperature, climate and vegetation cover in a Swedish acid mire. Biogeochemistry 1995, 28, 93–114. [Google Scholar] [CrossRef]
- Le Mer, J.; Roger, P. of methane by soils: A review. Eur. J. Soil Biol. 2001, 37, 25–50. [Google Scholar] [CrossRef]
- Koch, O.; Tscherko, D.; Kandeler, E. Seasonal and Diurnal Net Methane Emissions from Organic Soils of the Eastern Alps, Austria: Effects of Soil Temperature, Water Balance, and Plant Biomass. Arctic Antarct. Alp. Res. 2007, 39, 438–448. [Google Scholar] [CrossRef] [Green Version]
- Oertel, C.; Matschullat, J.; Zurba, K.; Zimmermann, F.; Erasmi, S. Greenhouse gas emissions from soils—A review. Chem. Erde 2016, 76, 327–352. [Google Scholar] [CrossRef] [Green Version]
- Whiting, G.J.; Chanton, J.P. Plant-dependant CH4 emission in a subartic Canadia fen. Glob. Biogeochem. Cycles 1992, 6, 225–231. [Google Scholar] [CrossRef]
- Joabsson, A.; Christensen, T.R.; Wallén, B. Vascular plant controls on methane emissions from northern peatforming wetlands. Trends Ecol. Evol. 1999, 14, 385–388. [Google Scholar] [CrossRef]
- Machacova, K.; Papen, H.; Kreuzwieser, J.; Rennenberg, H. Inundation strongly stimulates nitrous oxide emissions from stems of the upland tree Fagus sylvatica and the riparian tree Alnus glutinosa. Plant Soil 2013, 364, 287–301. [Google Scholar] [CrossRef]
- Klemedtsson, L.; Svensson, B.; Rosswall, T. Relationships between soil moisture content and nitrous oxide production during nitrification and denitrification. Biol. Fertil. Soils 1988, 6, 106–111. [Google Scholar] [CrossRef]
- Bateman, E.J.; Baggs, E.M. Contributions of nitrification and denitrification to N2O emissions from soils at different water-filled pore space. Biol. Fertil. Soils 2005, 41, 379–388. [Google Scholar] [CrossRef]
- Christiansen, J.R.; Vesterdal, L.; Gundersen, P. Nitrous oxide and methane exchange in two small temperate forest catchments—effects of hydrological gradients and implications for global warming potentials of forest soils. Biogeochemistry 2010, 107, 437–454. [Google Scholar] [CrossRef]
- Barrat, H.A.; Evans, J.; Chadwick, D.R.; Clark, I.M.; Le Cocq, K.; Cardenas, L.M. The impact of drought and rewetting on N2O emissions from soil in temperate and Mediterranean climates. Eur. J. Soil Sci. 2020, 1–13. [Google Scholar] [CrossRef]
- Schützenmeister, K.; Meurer, K.H.E.; Gronwald, M.; Hartmann, A.B.D.; Gansert, D.; Jungkunst, H.F. N2O emissions from plants are reduced under photosynthetic activity. Plant Environ. Interact. 2020, 1, 48–56. [Google Scholar] [CrossRef]
- Megonigal, J.P.; Brewer, P.E.; Knee, K.L. Radon as a natural tracer of gas transport through trees. New Phytol. 2020, 225, 1470–1475. [Google Scholar] [CrossRef]
Figure 1.
Environmental parameters for the three study periods: (a–c) air temperature; (d–f) soil temperature at 5 cm depth; (g–i) soil water content at 5 cm depth; (j–l) soil water level. Averaged values from 12 measuring points (air temperature from one central point) at 12:00 and 00:00 per day are presented.
Figure 1.
Environmental parameters for the three study periods: (a–c) air temperature; (d–f) soil temperature at 5 cm depth; (g–i) soil water content at 5 cm depth; (j–l) soil water level. Averaged values from 12 measuring points (air temperature from one central point) at 12:00 and 00:00 per day are presented.

Figure 2.
Stem CH4-C (a–c) and N2O-N (g–i) fluxes and soil fluxes of CH4-C (d–f) and N2O-N (j–l) during the three studied periods, each at daytime (12:00–16:00) and nighttime (0:00–04:00). Fluxes are expressed per m2 of stem or soil surface area. Please note different flux scales. Asterisks mark the statistically significant differences (p < 0.05) in fluxes between day- and nighttime Positive fluxes indicate gas emission, while the negative fluxes indicate gas uptake.
Figure 2.
Stem CH4-C (a–c) and N2O-N (g–i) fluxes and soil fluxes of CH4-C (d–f) and N2O-N (j–l) during the three studied periods, each at daytime (12:00–16:00) and nighttime (0:00–04:00). Fluxes are expressed per m2 of stem or soil surface area. Please note different flux scales. Asterisks mark the statistically significant differences (p < 0.05) in fluxes between day- and nighttime Positive fluxes indicate gas emission, while the negative fluxes indicate gas uptake.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schindler, T.; Machacova, K.; Mander, Ü.; Escuer-Gatius, J.; Soosaar, K. Diurnal Tree Stem CH4 and N2O Flux Dynamics from a Riparian Alder Forest. Forests 2021, 12, 863. https://doi.org/10.3390/f12070863
AMA Style
Schindler T, Machacova K, Mander Ü, Escuer-Gatius J, Soosaar K. Diurnal Tree Stem CH4 and N2O Flux Dynamics from a Riparian Alder Forest. Forests. 2021; 12(7):863. https://doi.org/10.3390/f12070863
Chicago/Turabian StyleSchindler, Thomas, Katerina Machacova, Ülo Mander, Jordi Escuer-Gatius, and Kaido Soosaar. 2021. "Diurnal Tree Stem CH4 and N2O Flux Dynamics from a Riparian Alder Forest" Forests 12, no. 7: 863. https://doi.org/10.3390/f12070863
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.