Interactive Effect of Elevated CO2 and Reduced Summer Precipitation on Photosynthesis is Species-Specific: The Case Study with Soil-Planted Norway Spruce and Sessile Oak in a Mountainous Forest Plot

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
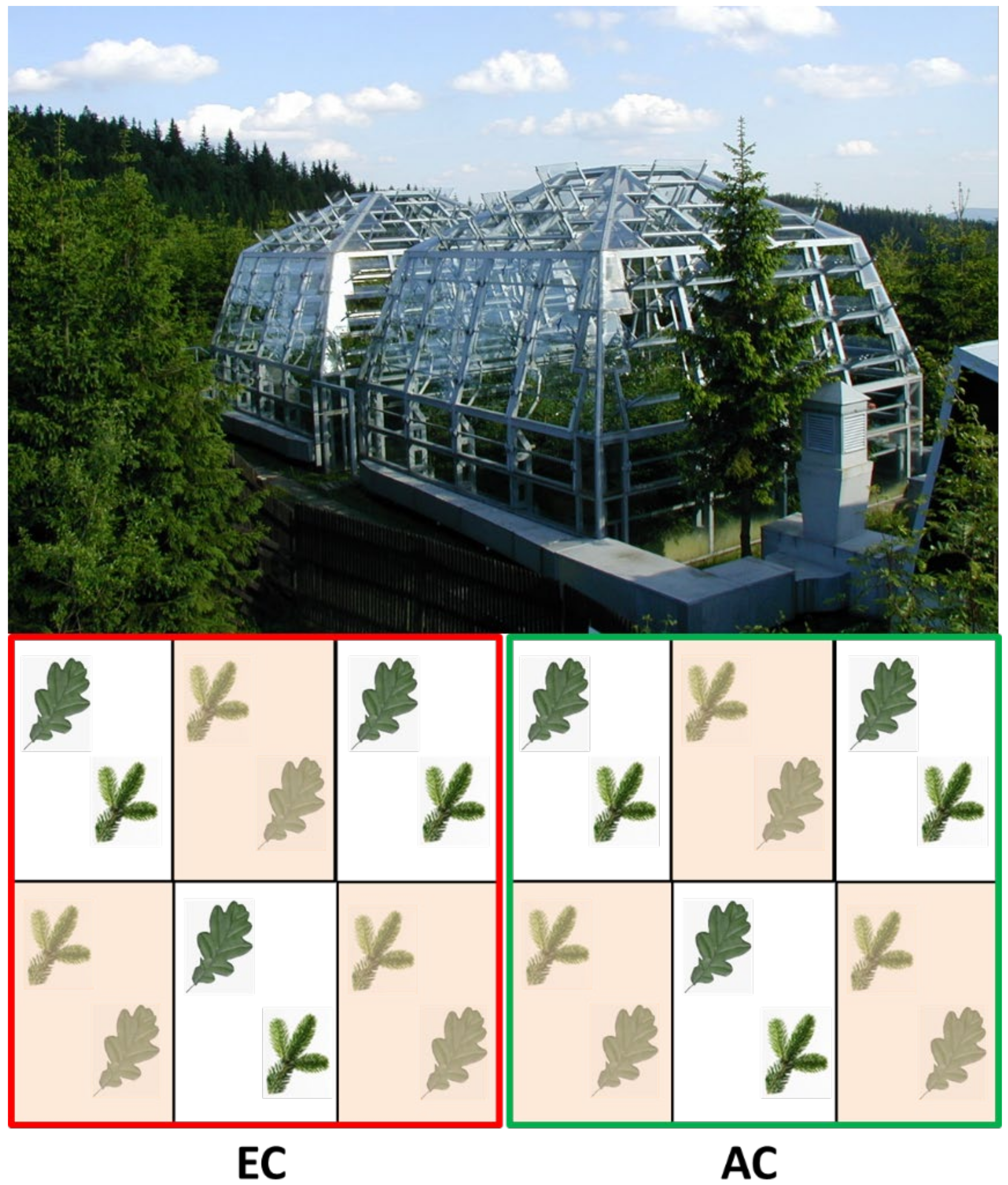
2.1. Plants and Experimental Design
2.2. Physiological Measurements
2.2.1. Gas-Exchange Measurements
2.2.2. Chlorophyll Fluorescence
2.3. Statistical Data Processing
3. Results
3.1. Growth Conditions
3.2. Gas-Exchange Parameters
3.2.1. Assimilation Capacity (Asat) and Light-Saturated CO2 Assimilation Rate (Amax)
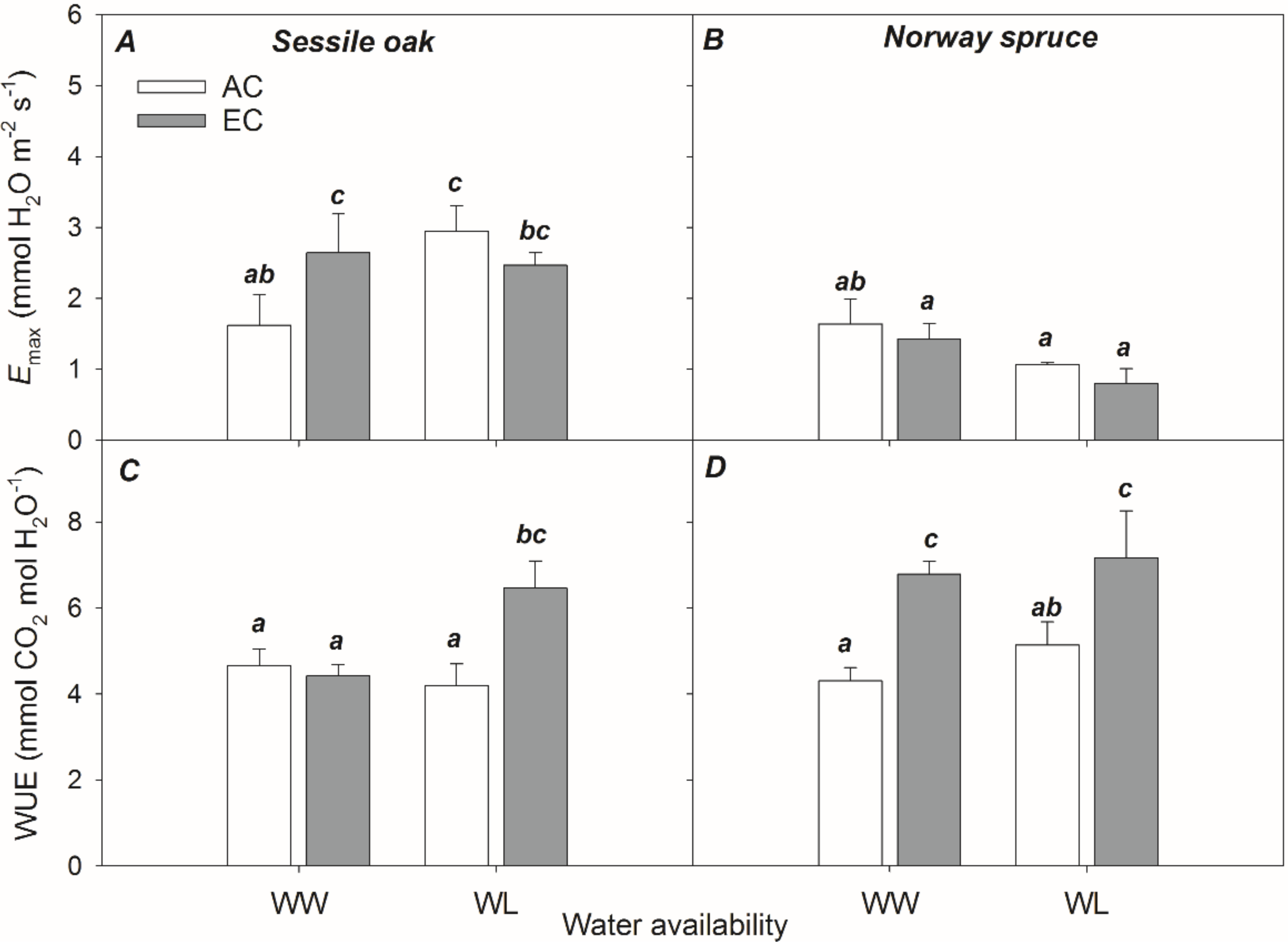
3.2.2. Light-Saturated Stomatal Conductance (GSmax) and Water Use Efficiency (WUE)
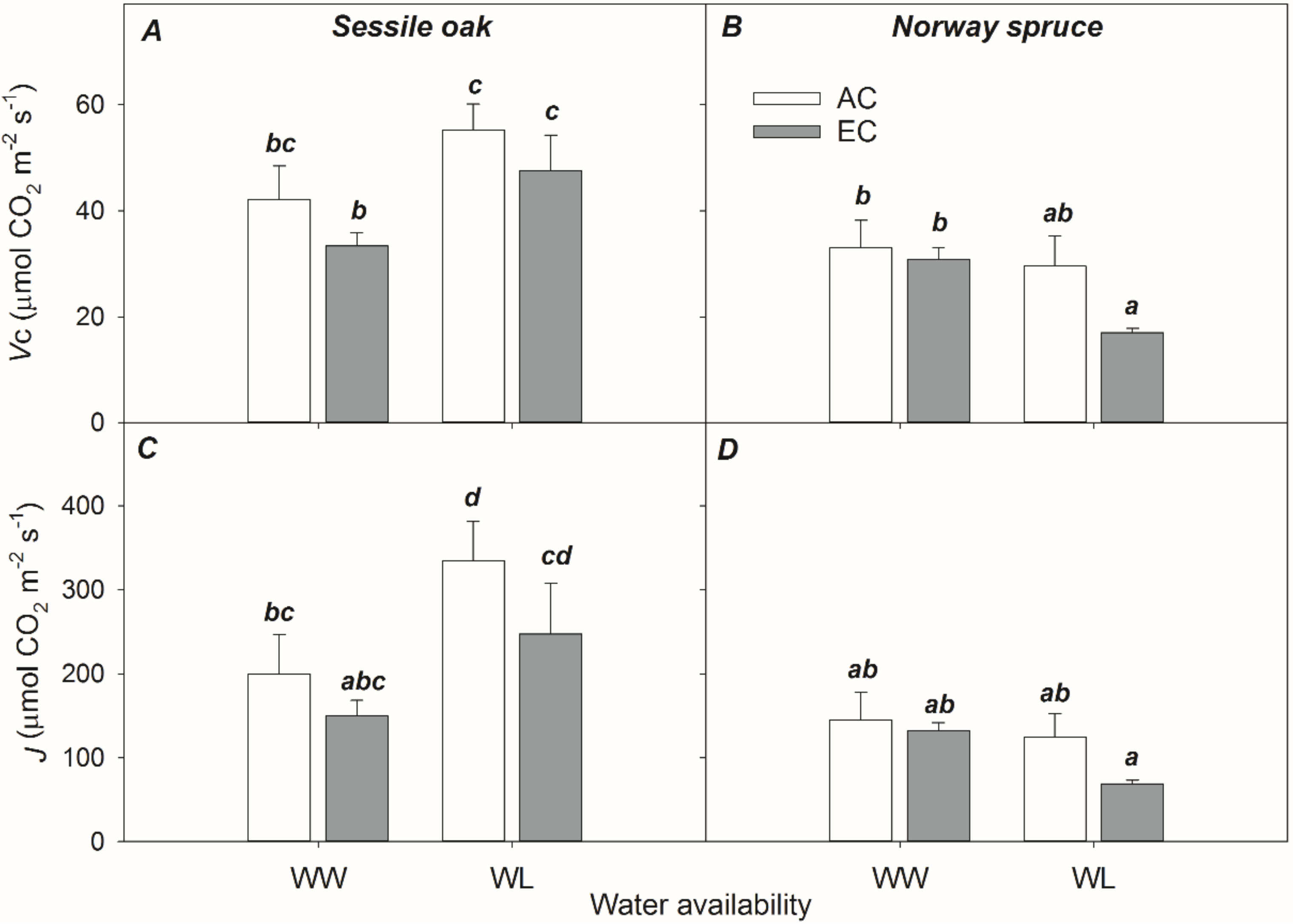
3.2.3. Biochemical Parameters Derived from A–Ci Response Curves
3.3. Chl-F Parameters
4. Discussion
4.1. Evaluation of Experimental Conditions
4.2. Species Specificity in Response to Elevated CO2 and Water Availability
4.2.1. Fluorescence Parameters
4.2.2. Gas-Exchange Parameters
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; DeFries, R.; Galloway, J.; Heimann, M.; et al. Carbon and other biogeochemical cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 465–570. [Google Scholar]
- Intergovernmental Panel on climate Chage (IPCC). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; pp. 1–151. [Google Scholar]
- Körner, C.; Asshoff, R.; Bignucolo, O.; Hättenschwiler, S.; Keel, S.G.; Peláez-Riedl, S.; Pepin, S.; Siegwolf, R.T.; Zotz, G. Carbon flux and growth in mature deciduous forest trees exposed to elevated CO2. Science 2005, 309, 1360–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuzinger, S.; Körner, C. Water savings in mature deciduous forest trees under elevated CO2. Glob. Chang. Biol. 2007, 13, 2498–2508. [Google Scholar] [CrossRef]
- Urban, O.; Klem, K.; Holišová, P.; Šigut, L.; Šprtová, M.; Teslová-Navrátilová, P.; Zitová, M.; Špunda, V.; Marek, M.V.; Grace, J. Impact of elevated CO2 concentration on dynamics of leaf photosynthesis in Fagus sylvatica is modulated by sky conditions. Environ. Pollut. 2014, 185, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medlyn, B.E.; Badeck, F.W.; De Pury, D.G.G.; Barton, C.V.M.; Broadmeadow, M.; Ceulemans, R.; De Angelis, P.; Forstreuter, M.; Jach, M.E.; Kellomäki, S.; et al. Effects of elevated [CO2] on photosynthesis in European forest species: A meta-analysis of model parameters. Plant Cell Environ. 1999, 22, 1475–1495. [Google Scholar] [CrossRef]
- Crafts-Brandner, S.J.; Salvucci, M.E. Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2. Proc. Natl. Acad. Sci. USA 2000, 97, 13430–13435. [Google Scholar] [CrossRef] [Green Version]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C3, C4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef]
- Badger, M.R.; von Caemmerer, S.; Ruuska, S.; Nakano, H. Electron flow to oxygen in higher plants and algae: Rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 1433–1446. [Google Scholar] [CrossRef] [Green Version]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Ruan, C.J.; Shao, H.B.; Teixeira da Silva, J.A. A critical review on the improvement of photosynthetic carbon assimilation in C3 plants using genetic engineering. Crit. Rev. Biotechnol. 2012, 32, 1–21. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, Y.; Jia, B.; Zhou, G. Elevated-CO2 response of stomata and its dependence on environmental factors. Front. Plant Sci. 2016, 7, 657. [Google Scholar] [CrossRef] [Green Version]
- Leakey, A.D.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Asensio, J.S.; Randall, L.; Rachmilevitch, S.; Cousins, A.B.; Carlisle, E.A. CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants. Ecology 2012, 93, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Townend, J. Effects of elevated carbon dioxide and drought on the growth and physiology of clonal Sitka spruce plants (Picea sitchensis (Bong.) Carr.). Tree Physiol. 1993, 13, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Dixon, M.; Thiec, D.L.; Garrec, J.P. The growth and gas exchange response of soil-planted Norway spruce [Picea abies (L.) Karst.] and red oak (Quercus rubra L.) exposed to elevated CO2 and to naturally occurring drought. New Phytol. 1995, 129, 265–273. [Google Scholar] [CrossRef]
- Picon, C.; Guehl, J.M.; Ferhi, A. Leaf gas exchange and carbon isotope composition responses to drought in a drought-avoiding (Pinus pinaster) and a drought-tolerant (Quercus petraea) species under present and elevated atmospheric CO2 concentrations. Plant Cell Environ. 1996, 19, 182–190. [Google Scholar] [CrossRef]
- Picon, C.; Guehl, J.M.; Aussenac, G. Growth dynamics, transpiration and water-use efficiency in Quercus robur plants submitted to elevated CO2 and drought. Ann. Sci. For. 1996, 53, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Tingey, D.T.; Phillips, D.L.; Johnson, M.G. Elevated CO2 and conifer roots: Effects on growth, life span and turnover. New Phytol. 2000, 147, 87–103. [Google Scholar] [CrossRef]
- Mildner, M.; Bader, M.K.F.; Baumann, C.; Körner, C. Respiratory fluxes and fine root responses in mature Picea abies trees exposed to elevated atmospheric CO2 concentrations. Biogeochemistry 2015, 124, 95–111. [Google Scholar] [CrossRef]
- Arora, V.K.; Boer, G.J.; Christian, J.R.; Curry, C.L.; Denman, K.L.; Zahariev, K.; Flato, G.M.; Scinocca, J.F.; Merryfield, W.J.; Lee, W.G. The effect of terrestrial photosynthesis down regulation on the twentieth-century carbon budget simulated with the CCCma Earth System Model. J. Clim. 2009, 22, 6066–6088. [Google Scholar] [CrossRef]
- Ruiz-Vera, U.M.; De Souza, A.P.; Long, S.P.; Ort, D.R. The role of sink strength and nitrogen availability in the down-regulation of photosynthetic capacity in field-grown Nicotiana tabacum L. at elevated CO2 concentration. Front. Plant Sci. 2017, 8, 998. [Google Scholar] [CrossRef]
- Urban, O.; Janouš, D.; Pokorný, R.; Marková, I.; Pavelka, M.; Fojtík, Z.; Šprtová, M.; Kalina, J.; Marek, M.V. Glass domes with adjustable windows: A novel technique for exposing juvenile forest stands to elevated CO2 concentration. Photosynthetica 2001, 39, 395–401. [Google Scholar] [CrossRef]
- Holub, P.; Klem, K.; Linder, S.; Urban, O. Distinct seasonal dynamics of responses to elevated CO2 in two understorey grass species differing in shade-tolerance. Ecol. Evol. 2019, 9, 13663–13677. [Google Scholar] [CrossRef] [PubMed]
- Urban, O.; Hrstka, M.; Holub, P.; Veselá, B.; Večeřová, K.; Novotná, K.; Grace, J.; Klem, K. Interactive effects of ultraviolet radiation and elevated CO2 concentration on photosynthetic characteristics of European beech saplings during the vegetation season. Plant Physiol. Biochem. 2019, 134, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Demmig-Adams, B.; Adams, W.W.; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264. [Google Scholar] [CrossRef]
- Trnka, M.; Balek, J.; Štěpánek, P.; Zahradníček, P.; Možný, M.; Eitzinger, J.; Žalud, Z.; Formayer, H.; Turna, M.; Nejedlík, P.; et al. Drought trends over part of Central Europe between 1961 and 2014. Clim. Res. 2016, 70, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Medlyn, B.E.; Barton, C.V.M.; Broadmeadow, M.S.J.; Ceulemans, R.; De Angelis, P.; Forstreuter, M.; Freeman, M.; Jackson, S.B.; Kellomaki, S.; Laitat, E.; et al. Stomatal conductance of forest species after long-term exposure to elevated CO2 concentration: A synthesis. New Phytol. 2001, 149, 247–264. [Google Scholar] [CrossRef]
- Urban, O. Physiological impacts of elevated CO2 concentration ranging from molecular to whole plant responses. Photosynthetica 2003, 41, 9–20. [Google Scholar] [CrossRef]
- Dieleman, W.I.; Luyssaert, S.; Rey, A.; de Angelis, P.; Barton, C.V.; Broadmeadow, M.S.; Broadmeadow, S.B.; Chigwerewe, K.S.; Crookshanks, M.; Dufrêne, E.; et al. Soil [N] modulates soil C cycling in CO2-fumigated tree stands: A meta-analysis. Plant Cell Environ. 2010, 33, 2001–2011. [Google Scholar] [CrossRef]
- Sardans, J.; Gargallo-Garriga, A.; Urban, O.; Klem, K.; Walker, T.W.N.; Holub, P.; Janssens, I.A.; Peñuelas, J. Ecometabolomics for a better understanding of plant responses and acclimation to abiotic factors linked to global change. Metabolites 2020, 10, 239. [Google Scholar] [CrossRef]
- Moran-Lopez, T.; Poyatos, R.; Llorens, P.; Sabate, S. Effects of past growth trends and current water use strategies on Scots pine and pubescent oak drought sensitivity. Eur. J. For. Res. 2014, 133, 369–382. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef] [PubMed]
- Raftoyannis, Y.; Radoglou, K. Physiological responses of beech and sessile oak in a natural mixed stand during a dry summer. Ann. Bot. 2002, 89, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K.; Ač, A.; Marek, M.V.; Kalina, J.; Urban, O. Differences in pigment composition, photosynthetic rates and chlorophyll fluorescence images of sun and shade leaves of four tree species. Plant Physiol. Biochem. 2007, 45, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Košvancová, M.; Urban, O.; Šprtová, M.; Hrstka, M.; Kalina, J.; Tomášková, I.; Špunda, V.; Marek, M.V. Photosynthetic induction in broadleaved Fagus sylvatica and coniferous Picea abies cultivated under ambient and elevated CO2 concentrations. Plant Sci. 2009, 177, 123–130. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Baba, W.; Horaczek, T.; Mojski, J.; Kociel, H.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Turcsán, A.; Maes, J.; Duchêne, N.; Meeus, S.; Steppe, K.; Steenackers, M. Repeated summer drought and re-watering during the first growing year of oak (Quercus petraea) delay autumn senescence and bud burst in the following spring. Front. Plant Sci. 2016, 7, 419. [Google Scholar] [CrossRef]
- Klein, T.; Ramon, U. Stomatal sensitivity to CO2 diverges between angiosperm and gymnosperm tree species. Funct. Ecol. 2019, 33, 1411–1424. [Google Scholar] [CrossRef]
- Mishra, K.B.; Mishra, A.; Novotná, K.; Rapantová, B.; Hodaňová, P.; Urban, O.; Klem, K. Chlorophyll a fluorescence, under half of the adaptive growth-irradiance, for high-throughput sensing of leaf-water deficit in Arabidopsis thaliana accessions. Plant Methods 2016, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- Kitao, M.; Koike, T.; Tobita, H.; Maruyama, Y. Elevated CO2 and limited nitrogen nutrition can restrict excitation energy dissipation in photosystem II of Japanese white birch (Betula platyphylla var. japonica) leaves. Physiol. Plant 2005, 125, 64–73. [Google Scholar] [CrossRef]
- Killi, D.; Bussotti, F.; Raschi, A.; Haworth, M. Adaptation to high temperature mitigates the impact of water deficit during combined heat and drought stress in C3 sunflower and C4 maize varieties with contrasting drought tolerance. Physiol. Plant 2017, 159, 130–147. [Google Scholar] [CrossRef]
- Wang, M.; Xie, B.; Fu, Y.; Dong, C.; Hui, L.; Guanghui, L.; Liu, H. Effects of different elevated CO2 concentrations on chlorophyll contents, gas exchange, water use efficiency, and PSII activity on C3 and C4 cereal crops in a closed artificial ecosystem. Photosynth. Res. 2015, 126, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Urban, O.; Hrstka, M.; Zitová, M.; Holišová, P.; Šprtová, M.; Klem, K.; Calfapietra, C.; De Angelis, P.; Marek, M.V. Effect of season, needle age and elevated CO2 concentration on photosynthesis and Rubisco acclimation in Picea abies. Plant Physiol. Biochem. 2012, 58, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Norby, R.J.; Warren, J.M.; Iversen, C.M.; Medlyn, B.E.; McMurtrie, R.E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl. Acad. Sci. USA 2010, 107, 19368–19373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Way, D.A.; Oren, R.; Kroner, Y. The space-time continuum: The effects of elevated CO2 and temperature on trees and the importance of scaling. Plant Cell Environ. 2015, 38, 991–1007. [Google Scholar] [CrossRef]
- Davey, P.A.; Olcer, H.; Zakhleniuk, O.; Bernacchi, C.J.; Calfapietra, C.; Long, S.P.; Raines, C.A. Can fast-growing plantation trees escape biochemical down-regulation of photosynthesis when grown throughout their complete production cycle in the open air under elevated carbon dioxide? Plant Cell Environ. 2006, 29, 1235–1244. [Google Scholar] [CrossRef]
- Pendall, E.; Bridgham, S.; Hanson, P.J.; Hungate, B.; Kicklighter, D.W.; Johnson, D.W.; Law, B.E.; Luo, Y.Q.; Megonigal, J.P.; Olsrud, M.; et al. Below-ground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods, and models. New Phytol. 2004, 162, 311–322. [Google Scholar] [CrossRef]
- Iversen, C.M. Digging deeper: Fine-root responses to rising atmospheric CO2 concentration in forested ecosystems. New Phytol. 2010, 186, 346–357. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Polley, H.; Johnson, H.; Fay, P.; Sanabria, J. Initial response of evapotranspiration from tallgrass prairie vegetation to CO2 at subambient to elevated concentrations. Funct. Ecol. 2008, 22, 163–171. [Google Scholar]
- Kanniah, K.D.; Beringer, J.; Hurley, L. Exploring the link between clouds, radiation, and canopy productivity of tropical savannas. Agric. For. Meteorol. 2013, 182, 304–313. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Ψpre-dawn | Ψnoon | VC | J | Asat | Amax | GSmax | Emax | WUE | ΦPSII | D |
|---|---|---|---|---|---|---|---|---|---|---|---|
| TS | <0.001 | 0.036 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | 0.134 | <0.001 |
| [CO2] | 0.001 | 0.217 | <0.001 | 0.003 | 0.691 | 0.008 | 0.573 | 0.921 | <0.001 | 0.574 | 0.840 |
| WA | 0.002 | 0.015 | 0.211 | 0.022 | 0.017 | 0.066 | 0.970 | 0.910 | 0.008 | 0.363 | 0.279 |
| TS × [CO2] | 0.086 | 0.701 | 0.827 | 0.265 | <0.001 | 0.128 | 0.228 | 0.071 | 0.016 | 0.006 | <0.001 |
| TS × WA | 0.374 | 0.217 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.696 | 0.047 | 0.002 |
| [CO2] × WA | 0.374 | 0.956 | 0.245 | 0.191 | 0.016 | 0.452 | 0.002 | 0.010 | 0.040 | 0.566 | 0.973 |
| TS × [CO2]× WA | 0.086 | 0.783 | 0.156 | 0.910 | 0.010 | 0.137 | 0.006 | 0.016 | 0.005 | 0.615 | 0.234 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ofori-Amanfo, K.K.; Klem, K.; Veselá, B.; Holub, P.; Agyei, T.; Marek, M.V.; Grace, J.; Urban, O. Interactive Effect of Elevated CO2 and Reduced Summer Precipitation on Photosynthesis is Species-Specific: The Case Study with Soil-Planted Norway Spruce and Sessile Oak in a Mountainous Forest Plot. Forests 2021, 12, 42. https://doi.org/10.3390/f12010042
Ofori-Amanfo KK, Klem K, Veselá B, Holub P, Agyei T, Marek MV, Grace J, Urban O. Interactive Effect of Elevated CO2 and Reduced Summer Precipitation on Photosynthesis is Species-Specific: The Case Study with Soil-Planted Norway Spruce and Sessile Oak in a Mountainous Forest Plot. Forests. 2021; 12(1):42. https://doi.org/10.3390/f12010042
Chicago/Turabian StyleOfori-Amanfo, Kojo Kwakye, Karel Klem, Barbora Veselá, Petr Holub, Thomas Agyei, Michal V. Marek, John Grace, and Otmar Urban. 2021. "Interactive Effect of Elevated CO2 and Reduced Summer Precipitation on Photosynthesis is Species-Specific: The Case Study with Soil-Planted Norway Spruce and Sessile Oak in a Mountainous Forest Plot" Forests 12, no. 1: 42. https://doi.org/10.3390/f12010042