
Taxonomy and Biogeography of Andinia-Complex (Orchidaceae)


1
Department of Plant Taxonomy and Nature Conservation, University of Gdańsk, Wita Stwosza 59, 80-308 Gdańsk, Poland
2
Department of Evolutionary Genetics and Biosystematics, University of Gdańsk, Wita Stwosza 59, 80-308 Gdańsk, Poland
3
Department of Geobotany and Plant Ecology, Faculty of Biology and Environmental Protection, University of Lodz, Banacha 12/16, 90-237 Lodz, Poland
4
Department of Biodiversity Research, Global Change Research Institute AS CR, Bělidla 4a, 603 00 Brno, Czech Republic
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(5), 372; https://doi.org/10.3390/d14050372
Submission received: 3 February 2022
/
Revised: 28 April 2022
/
Accepted: 29 April 2022
/
Published: 7 May 2022
(This article belongs to the Special Issue Distribution and Diversity of Orchids)
Abstract
:The results of a revision of the orchid genus Andinia are presented. The proposed classification is based on outcomes of morphological and molecular studies. Additionally, the similarity of the climatic niches occupied by representatives of the Andinia-complex was calculated and visualized. The highest number of Andinia representatives is found in the Eastern Cordillera real montane forests, while Neooreophilus species are found in two ecoregions—the Eastern Cordillera real montane forests, and the Northwestern Andes. According to the level of endemism, a very unique orchid flora was noted in the Guajira–Barranquilla xeric scrub ecoregion, as well as in the Isthmian-Pacific and the Ucayali moist forests, where single endemic species occur. In the proposed classification, nine genera are recognized within the Andinia-complex, three of which (Xenosiella, Chicalia, and Minuscula) are described in this paper. A key to the identification of all genera is provided. Additionally, sections Amplectentes and Bilamellatae of Neooreophilus are elevated to the subgeneric rank, and subgenus Aenigma is here considered as a separate genus. A complete list of representatives of each taxon is provided, and new combinations are proposed accordingly. A new species of Neooreophilus from Colombia is described.

1. Introduction
The Neotropical subtribe Pleurothallidinae, comprising ca. 5000 species, is one of the largest groups within Orchidaceae [1,2]. The generic composition of this taxon has been studied for years [1,3,4,5], but the taxonomic concept of numerous genera is still debated [6,7,8,9,10] due to the enormous morphological diversity of pleurothallid orchids and the relatively poor molecular data which could be used to delineate well-defined taxa.
The genus Lepanthes described at the end of the XVIII century is one of several hyper-diverse genera within Pleurothallidinae [11]. In 1986, it was divided by Luer into four subgenera—L. subgen. Brachycladium Luer, L. subgen. Draconanthes Luer, L. subgen. Lepanthes, and L. subgen. Marsipanthes Luer. The former two were later considered as separated genera—Brachycladium Luer (Luer) [12] and Draconanthes Luer (Luer) [1], respectively. Brachcladium was, however, illegitimate, as the same name was previously used for a genus of fungus. For this reason, Archila and Higgins [13] proposed a substitute name, Oreophilus W.E. Higgins & Archila, for Brachycladium, but again, the authors made a nomenclatural mistake, and finally in 2009 a correct name, Neooreophilus Archila, was given to encompass pleurothallids characterized by long-repent, pendent habit, creeping rhizomes, leaves appearing alternating, and Lepanthes-like flowers [14]. The concept of Neooreophilus was accepted for eight years until Wilson et al. [15], based on phylogenetic analyses, proposed to group the genus representatives with Andinia (Luer) Luer, which in the suggested broad concept included also Masdevalliantha (Luer) Szlach. & Marg., and Xenosia Luer. Andinia was initially recognized as a subgenus of Salpistele Dressler and encompassed plants with repent habit, echinate ovaries, abbreviated petals, presence of callus on the lip disc, and large, auriculate lip lateral lobes. Pridgeon and Chase [4] later transferred to this genus also a single representative of Lueranthos Szlach. & Marg.
In this broad concept, morphological variation of Andinia s.l. is significant in aspects of vegetative and floral characters. Representatives of this complex are caespitose or rhizomatous plants with variously developed rhizome. Their leaves of various shape are glabrous or with ciliate or denticulate margins, and in numerous species they appear alternating and often overlapping. The inflorescence of these plants is single-flowered to racemose, sometimes reclining. The ovary is glabrous to papillose, verrucose, echinate, or spiculate. Sepals are spiculate or glabrous, variously connate, while the petals vary in size (from microscopic to subequal to sepals) and form (suborbicular to variously lobed and trifurcate). In addition, the lip form significantly differs among the Andinia-complex. It is entire to variously lobed, sometimes with an appendix, with or without callus. The gynostemium is usually (but not always) footless, with elongated or reduced rostellum.
The discrepancy between the genetic and morphological and/or ecological data together with the question about the precision of using a barcoding approach were recognized as serious problems in taxonomy of numerous organisms [16,17]. As summarized by Wiens et al. [18], while the rates of genome sequencing increase, scientists should continue to construct classification systems using morphological data. A reason supporting morphological analyses is that DNA of many taxa, including numerous Andinia s.l., have not been sequenced, and many species remain known from a single specimen that was collected decades ago. Moreover, because various issues may result in the creation of false but statistically well-supported molecular phylogenies [18], it is still important to develop accurate, morphology-based phylogenies. Noteworthily, a typical set of morphological characters is information obtained from various unlinked genes, while a molecular dataset is often linked and inherited as a single unit [19].
The substantial differences in the morphology within Andinia sensu Wilson et al. [15] and the obvious discontinuum in numerous characters prompted us to revise the classification of species included in this genus. Moreover, we tested if species classified in Andinia s.l. exhibit any ecological differences. Based on the confrontation of morphological, molecular, and ecological data, we propose a new taxonomic concept of Andinia s.l.
2. Materials and Methods
2.1. Phylogenetic Analyses
For phylogenetic reconstruction, we applied the ITS sequences and the matK gene (70/40), representing taxa from six genera of the Andinia-complex: Andinia s.str., Lueranthos, Masdevalliantha, Pleurothallis, Neooreophilus, and Xenosia. Laelia was used as an outgroup. All sequences were downloaded from GenBank (https://www.ncbi.nlm.nih.gov/genbank/, accessed on 1 June 2021), and the accession numbers were placed on the cladograms together with the names of the species. We performed three matrixes, two for single markers (ITS, matK), and combined both these markers as well. The sequences were automatically aligned by SeaView [20], choosing the MUSCLE algorithm [21]. The generated matrices were analyzed under maximum likelihood (ML) and Bayesian Inference (BI). The ML/Transfer bootstrap expectation workflow analysis was performed using RAxML-HPC2 [22] by searching for the best-scoring ML tree with the GTR+I+G as the best fit substitution model. The branch support values (BS) were calculated with RaxML by halting bootstrapping automatically under the autoMRE criterion. For bootstrap support levels, we considered the following percentage values of 50–70% as weak, 71–85% as moderate, and >85% as strong [23].
The BI was performed using MrBayes v. 3.2.7a [24] with GTR+I+G as the selected evolutive model for all matrices. An evolutionary model was determined with MrModeltest 2.2 [25] and selected according to the Akaike Information Criterion (AIC). Each analysis consisted of two independent runs of four chains, one hot and three cold chains, and were started from different random trees to ensure that individual runs had converged to the same result. We used 10 million generations per runs with a sampling frequency of 100. Split frequencies below 0.01 were used to check for convergence, and the effective sample size (ESS) for each run was checked in Tracer v. 1.7.1 [26]. Twenty-five percent of trees were excluded as burn-in. Saved trees were summarized in a majority rule consensus tree created with nodal confidence assessed by posterior probabilities (PP), which were considered strongly supported when equal to or higher than 0.95. The trees obtained in both methods were edited with FigTree v.1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/ accessed on 1 June 2021) and Inkscape (https://inkscape.org/release/inkscape-1.0.2/ accessed on 1 June 2021).
2.2. Taxonomic Studies
The database of diagnoses and original illustrations of 18 Andinia s.str., 52 Neooreophilus, 3 Xenosia, 2 Masdevalliantha, and single Lueranthos species was compiled. A total of over 300 herbarium specimens and liquid preserved flowers of orchids included by Wilson et al. [15] in Andinia deposited in AMES, BM, COL, JAUM, K, MO, NY, U, US, UGDA, and W [27] was examined according to the standard procedures. Additionally, plants cultivated in Ecuagenera, Mundiflora, Ecuaflora, and the private collection of Ramiro Medina Trejo were analyzed. Every studied herbarium sheet was photographed, and the data from the labels were taken. Both vegetative and generative characters of each plant were examined. The form and surface of leaves were studied first. Then the construction of the inflorescence and the shape and size of the floral bracts were examined. The morphology of flowers was studied after their boiling using a stereomicroscope. The database of vegetative and floral characters of all species was compiled to facilitate comparison of Andinia-complex representatives (Annex S1).
To propose the comprehensive classification of Andinia s.l., we used morphological data gathered in this research as well as outcomes of the phylogenetic analyses conducted for the sequences downloaded from GenBank.
2.3. Variation of Climatic Preferences and Biogeographical Analyses
The database of localities for species representing the Andinia-complex was compiled. Only localities which should be localized based on information included in herbarium specimen labels were included in the distribution maps (Figure 1) created using ArcGis 10.6 (Esri, Redlands, CA, USA). We were able to obtain precise (precision of 10 km) coordinates for 93 various locations and assign 122 records to specific terrestrial ecoregions [28]. The datasets are presented as Annexes S2 and S3. To visualize climatic niches of the studied taxa, multivariate analyses were performed. A preliminary Detrended Correspondence Analysis (DCA) showed relatively low values of gradient length (<3 SD units). Therefore, a principal component analysis (PCA) was used as the most appropriate method for examine species variation within different climatic spaces and to account for the multicollinearity nature of the initial variables. Analysis was carried out based on 12 of the 19 available bioclimatic variables (Table 1 [29]), also used in ENM analysis.
The analyses were carried out with the division into studied genera and species, where more than one record was recorded. For each vector, the determination coefficient R2 and p-value were calculated using a permutation test with 999 iterations. To determine the bioclimatic variables which differentiate the studied orchids the most, canonical variate analysis (CVA) was applied in order to reduce the dataset by selecting only the factors that showed the strongest discrimination. The significance of bioclimatic differences between the studied taxa was evaluated using Wilk’s λ with 10,000 permutations. In turn, the Bray–Curtis similarity between samples was calculated to classify the terrestrial ecoregions, in which species occur at sites, and the resulting matrix was then used in the cluster analysis UPGMA (unweighted pair-group average method). The Shannon–Wiener index was calculated to compare species diversity between terrestrial ecoregions. All analyses were performed using PAST v. 3.20 software [30].
3. Results
3.1. Phylogeny
3.1.1. ITS Matrix
On the tree obtained by Bayesian inference for the ITS marker, we could distinguish six main and strongly supported phylogenetic groups (Figure 2, clades I–VI).
Clade I (pp = 1.00) was represented by only one species, Andinia spiralis, whereas the second group (clade II, Figure 2) included samples of representatives of Andinia longiserpens, combined with Andinia xenion (pp = 0.99). Clade III, also with a high value of posterior probability (pp = 0.98), was represented by members of Neooreophilus, and according to the results of analysis of the ITS sequence, this clade was fragmented into three suballiances. The following three groups (clades IV–VI, Figure 2) included representatives of Andinia s.l. with a high value of probability. The first one (clade IV) included A. trimytera and A. vestigipetala, while clade V encompassed A. vestigipetala, A. lappacea, A. pensilis, and A. cf. dielsii. A. vestigipetala was sampled by two sequences (KP012349.1 and MK294775.1) placed in two different clades. Probably it could be the result of an incorrectly labelled sample, misidentification, or contamination of the sample. The last clade with high pp value comprised three species: A. schizopogon, A. dalstroemii, and A. pogonion.
The topology of the tree (Figure 3) obtained for the same marker but based on the ML method was slightly different. Samples representing the following species did not form a consistent clade: A. schizopogon, A. dalstroemii, and A. pogonion. A. spiralis (clade a), A. longiserpens, and A. xenion (clade b) clustered together in a strongly supported group (clade I, BS = 89). On the ML tree, we could distinguish three better supported groups of taxa (clades II–IV). Clade II (BS = 90) included A. vestigipetala, A. lappacea, A. pensilis, and A. cf. dielsii and overlapped with clade V on the BI tree (Figure 2). Group III (Figure 3) was formed by the same taxa as clade V on the Bayesian inference tree (Figure 2). Clade IV (Figure 3) corresponded as well to the results based on the BI method (Figure 2, clade III), and included representatives of Neooreophilus.
3.1.2. matK Matrix
Both Bayesian inference (Figure 4) and maximum likelihood analysis (Figure 5) performed for matK provided quite interesting results—relationships between particular clades were not solved. However, all clades had high values of posterior probability and bootstrap support. Species contents of all of these groups were similar to the results obtained for the ITS marker.
3.1.3. Combined Matrix
The topology of trees obtained for the combined matrix (ITS + matK) (Figure 6 and Figure 7) was similar to that based on the ITS marker. We could distinguish three main clades (I–III, Figure 6 and Figure 7). Two of these included representatives of A. xenion and A. longiserpens (clade I), and the other was represented by A. spiralis (clade II). However, within the last one (clade III, Figure 6), on the tree based on BI analysis, we could recognize the following three subgroups: A, B, and C. The first one (A) included representatives of Neooreophilus (pp = 1.00), and the next one (B, Figure 6) included samples of A. pensilis, A. dielsii, A. lappacea, and A. vestigipetala. The last group—C—with A. schizopogon, A. pogonion, and A. dalstroemii has also a high value of probability (Figure 6, pp = 1.00). On the maximum likelihood tree, clade III (Figure 7) split into two subgroups: A and B. Within the first of these (A) we could also distinguish two subclades (C and D) with strong bootstrap support. Subclade C included Neooreophilus representatives (BS = 100), and subclade D included A. pensilis, A. dielsii, A. lappacea, and A. vestigipetala (BS = 93). A. trimytera was also placed in group A (Figure 7) as a sister to subclade D. The second group—B—with A. schizopogon, A. pogonion, and A. dalstroemii also had strong bootstrap support (Figure 7, BS = 1.00).
3.2. Climatic Niche Similarity
The general similarity of the climatic niches occupied by genera within the Andinia-complex is visible in the PCA graph (Figure 8) and explained 96.13% of variance. Variables with the greatest contributions enhanced in the overall analysis were: the temperature seasonality (bio4); the annual precipitation (bio12); the precipitation of the warmest quarter (bio18); and the precipitation of the coldest quarter (bio19). The main shift in differentiation of climatic niches was along the PCA axis 2, where Andinia, Aenigma, Masdevalliantha, and Lueranthos were separated on the left side of the diagram. However, the climatic niches for all genera overlapped in this plot. There were no clear boundaries between the studied genera in terms of their preferred climatic conditions. In turn, the canonical variate analysis (CVA) showed a highly statistically significant differential value for the studied genera with respect to the bioclimatic variables (Wilks λ = 0.200; F(72;577) = 2.762; p < 0.001). The canonical variate axes (56.24 and 20.00% of explained variance, respectively) distinguished specimens from each other along the environmental gradient derived from bioclimatic variables, similar to the PCA analysis (Figure 9).
In the next PCA analysis including species within the Andinia-complex, the first axis distinguished Masdevalliantha longiserpens, Neooreophilus chaoae, N. monilius, N. montis-rotundus, and Xenosia xenion (above the axis) from Andinia dielsii, A. dalstroemii, A. pogonion, A. pentamytera, and N. lupulus (below the axis). Furthermore, the second axis separated Aenigma schizopogon, Lueranthos vestigipetalus, Masdevalliantha longiserpens, M. masdevalliopsis, and N. stalactites on the left side of the diagram (Figure 10). Variables with the greatest contributions enhanced in the analysis were: the annual precipitation (bio12); the precipitation of the wettest month (bio13); the precipitation of the warmest quarter (bio18); and the precipitation of the coldest quarter (bio19); and also the temperature seasonality (bio4) (Table 2). The same variables indicated in the above analysis were significant contributions to the discrimination of the studied species. This analysis distinguished most species of Neooreophilus (e.g., N. chaoae, N. ciliaris, N. hippocrepicus, N. lunaris, N. lupulus, N. micropetalus, N. montis-rotundus, N. nummularius) on the right side of the diagram (Figure 11). The described shift in discrimination of climatic niches was along Axis 2 (Wilks λ = 0.002; F(240;651) = 2.295; p < 0.001).
3.3. Biogeography
Two main groups were recognized in the UPGMA dendrogram illustrating the similarity between analyzed ecoregions in the occupied climatic niches (Figure 12). One of them included mountain forests of the Eastern and Oriental Cordillera, the Northwestern Andean montane forests, the Northern Andean páramo, and the Cauca Valley montane forests ecoregions. The second group was comprised of the Magdalena Valley montane forests and the Napo moist forests. The Peruvian Yungas remained separate.
The highest values of the Shannon–Wiener index for taxa belonging to the Andinia-complex were obtained in montane forests of the Eastern Cordillera (H′ = 1.26) and the Northwestern Andean montane forests (H′ = 1.08). The highest number of Andinia representatives was found in the Eastern Cordillera real montane forests (29 species), while Neooreophilus species were found in two ecoregions—the Eastern Cordillera real montane forests, and the Northwestern Andean montane forest (20 and 8 species, respectively). According to the level of endemism, a very unique orchid flora was noted in the Guajira–Barranquilla xeric scrub ecoregion, as well as in the Isthmian-Pacific and the Ucayali moist forests, where single endemic species occur (N. triangularis, A. lappacea, A. tingomariana, respectively).
4. Discussion
4.1. Total Evidence Classification
The problem of non-monophyletic taxa is rather common in plant classification, and still numerous recognized genera are not monophyletic [31,32,33,34]. Paraphyletic groups are often acknowledged as separated genera mostly because their morphological and ecological characters prevented taxonomist from lumping them into larger, monophyletic taxa. Ideally, a “good” genus should be recognizable in molecular, morphological, and also ecological attributes. Unfortunately, the availability of data on various molecular markers in orchids is still relatively poor and most phylogenetic studies rely on just several barcode loci—ITS, trnL, rbcL, matK, psbA-trnH, and psbK [35,36,37]. Incompatibilities between classification systems derived from morphological and molecular analyses, and between phylogenetic trees based on various subsets of molecular sequences has become widespread [38,39]. Tree topology conflicts can be explained either by hybridization or incomplete lineage sorting [40]. The incongruencies between morphological and molecular data were observed in numerous orchids, e.g., with the complex Erycina–Stacyella–Psygmorchis [41], genus Otoglossum [42], numerous oncidioid orchids [43], and pleurothallids [8,44]. Hopefully, the development and availability of more advanced techniques of DNA analyses will bring some conclusions into Orchidaceae taxonomy [45].
4.2. Classification of Andinia
The genus Andinia has been recently revised and re-circumscribed by Wilson et al. [15]. The authors described the genus as follows: “Plants caespitose or rhizomatous in habit; in the latter the rhizome repent, creeping or pendulous (similar to species of Brachionidium). The inflorescence mostly successively multi-flowered, with only one flower open at a time. Ovaries glabrous to echinate. The flowers of some species similar to those of Lepanthes. The petals mostly very much abbreviated compared to the sepals. The lip three-lobed (very shallowly in a few species), with the mid-lobe modified into an appendix in many species, and the lateral lobes frequently surrounding the column. Only a couple of species do not have an apical anther and stigma, but all have drop-like pollinaria, with a bubble-like viscidium”. We decided to cite expressis verbis Wilson’s et al. [15] definition of Andinia s.l. to make evident the problem in identification the unique combination of characters of this genus in its widest concept.
The habit detected in Andinia s.l. is not unique at all, as was previously suggested by the authors. There can be distinguished two main habit types. The first one includes species with dangling rhizome, with short, pendulous ramicaul enclosed in typical, lepanthiform sheath(s) with ciliate or echinate margins. They produce orbicular to elliptic leaves, often with ciliate margins and/or upper surface. All of them have lepanthiform flowers with more or less connate sepals, and altered petals covered by various kinds of ciliae or hairs. The lip is simple in form, ecallose, canaliculated, and enclasping the gynostemium. The gynostemium is also lepanthiform, i.e., footless, terete, with erect anther lying on the dorsal surface of the column, and elongate, subulate rostellum. The stigmatic surface is subapical, horizontal, which facilitates grabbing the pollen mass. As thus can be seen, species of this group are similar to the representatives of Lepanthes, and in fact for a long time all of them have been accommodated in this genus. Archila and Higgins [13] were the first who emphasized differences between this group of species versus other Lepanthes, giving them a rank of a separate genus (Oreophilus W. Higgins & Archila), but for nomenclatural reason changing it soon to Neooreophilus [14]. Except for morphological background, the genus is supported by phylogenetic analyses in which all species belonging here create a monophyletic clade, although with various degrees of support.
Lepanthiform sheaths can be found also in Lepanthes and Trichosalpinx. Moreover, flowers of Lepanthes and Neooreophilus are similar in having basally connate sepals which are often widely spread and forming a kind of triangle, modified petals, and surface of various flower segments covered by ciliae or other protuberances. According to Wilson et al.’s [15] phylogenetic analyses, however, these genera and Neooreophilus are only distantly related. It is interesting to note that flowers of Lepanthes–Trichosalpinx are borne below, whereas in Neooreophilus they are above or upon the leaf blade. Phylogenetic studies can suggest independent origin of similar sheaths and other characters in both Neooreophilus and Lepanthes–Trichosalpinx. In our opinion it is necessary to consider as well a hybrid origin of the genus Neooreophilus. The putative parents could be Lepanthes and Andinia species. It would be interesting to study sequences of other markers than matK, ITS, or rbcL in a wide spectrum of Pleurothallidinae.
While Neooreophilus was not well-sampled, several morphologically consistent groups can be recognized in the phylogenetic tree. The first subclade (B3) (Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7) includes samples of species with ascending inflorescence borne above the leaf surface. Closely related N. compositus, N. persimilis, N. pilosellus, and N. platysepalus are characterized by hirsute or ciliate–denticulate leaves and cucullate dorsal sepal. The group composed by N. ciliaris and N. lynnianius, which are characterized by glabrous leaves, is sister to N. werneri, which has hirsute or ciliate–denticulate leaves and not cucullate dorsal sepals. The generitype of Neooreophilus, N. nummularius (B4) (Figure 2, Figure 3, Figure 6 and Figure 7), characterized by transversely ovate to trapeziform lip without an appendix, is sister to N. stalactites (B5) (Figure 2, Figure 3, Figure 6 and Figure 7), which is the only representative of the genus having a bilaminate lip with an appendix.
The second large group of species is relatively uniform as the habit is considered and completely different from the aforementioned one. The plants are usually small to even tiny with short, creeping, or ascending rhizome. A short, erect ramicaul is enclosed in tight sheaths. Leaf is narrow, oblong to oblanceolate, always glabrous. Usually the few-flowered inflorescence is erect, rarely reclining, and longer than leaf. Both flower segments and reproductive structures are much more diversified than in Neooreophilus, and it is difficult to detect a common character to all of the species belonging here. This is not surprising, as according to the results of molecular analyses, they are paraphyletic, unlike Neooreophilus, which is monophyletic and strongly supported.
The clade B1 (Figure 2, Figure 3, Figure 6 and Figure 7) is much more complicated. It is represented in our study by A. pensilis, A. dielsii, A. lappacea, and A. vestigipetala. This clade is strongly supported on the trees obtained based on both ITS and combined matrices (Figure 2, Figure 3, Figure 6 and Figure 7), while on the cladograms performed for the matK matrix, the relation between these taxa are unsolved (Figure 4 and Figure 5). In fact, it is very difficult to find a common character for such set of species. The only one we could detect are relatively small or even rudimentary petals. It is much easier to single out features characterized for each of them separately. A. pensilis, to which we can also classified morphologically very similar A. dielsii (a generitype of Andinia) and the recently described A. tingomariana, have a very peculiar gynostemium form, which can be compared to that found in Neooreophilus and which could be described as lepanthiform, i.e., footless, erect, terete, with erect anther lying on its upper surface. The erect rostellum is much elongated, elliptic to oblong, and much exceeding the anther. The stigma is ventral. In all aforementioned species, the lip is basally (or even subapically in A. tingomariana) connate with the gynostemium, more or less 3-lobed with both lateral lobes being upcurved and enclasping the gynostemium.
The subclade 1 (Figure 6 and Figure 7), including A. pensilis and A. dielsii, is sister to the pair of sample representatives A. lappacea and A. vestigipetala (Figure 6 and Figure 7), and it has both high value of probability (pp = 0.99) and bootstrap support (BS = 93). A similar situation we observed also on the trees preformed on the ITS marker (clade V, Figure 2 and Figure 3).
Andinia vestigipetala is unique in Pleurothallidinae in having erect, terete gynostemium, which is connate with rudimentary petals combined with simple, canaliculated lip, glabrous, free sepals, and glabrous ovary. A. lappacea is characterized by connate lateral sepals, and a subquadrate, ciliate lip, which is basally reflexed and apically emarginated. Sepals are densely ciliate–papillate. The gynostemium is short, clavate, slightly sigmoid, and basally enclasped by the lip. The ovary is echinate. We cannot find any mutual character for this pair except vestigial petals. The subclade A. pensilis, A. vestigipetala, and A. lappacea is weakly supported with Neooreophilus and A. trimytera.
However, based on the similarity in morphology of the flowers, the following species can be classified here as well: A. hystricosa, A. panica, A. sunchubambensis, and A. wayqechensis. All of them can be characterized by subsimilar sepals, of which both lateral ones are connate together in the lower half, prominent petals (except A. trimytera, where they are obscure), and peculiar lip. The lip is subsessile, strongly reflexed near the middle, with its basal part enclasping the gynostemium. Its apical part is deeply 3-lobed, with all lobes being subsimilar, more or less elongate. The central part of the lip is adorned by an elevated callus. The gynostemium is also uniform, i.e., footless, elongate, clavate, apically somewhat bent forward, and the apical clinandrium is obscure, the anther is apical and bent forward, and the rostellum is triangular–ligulate and bent over the stigmatic surface, which is ventral. The ovary is always hispid or muricate. All of these characters are shared by two additional species, A. pentamytera and A. ibex, despite some differences in their lip morphology (e.g., additional lobules in the sinus between middle and lateral lobes in the former, and two minute calli on the lip with widely spread lateral lobes in the latter one).
The taxa of group A (clade IV Figure 2 and clade C Figure 6) is represented by A. pogonion, A. schizopogon, and A. dalstroemii. In our opinion, two other species should be classified with them, i.e., A. hirtzii and A. uchucayensis, as they share many common characters. In all of them the ovary is echinate, lateral sepals are basally connate, petals are prominent, linear–lanceolate, acute to long–acuminate or even caudate, and the lip is sessile, gently reflexed near the middle, enclasping the gynostemium. The lip is suborbicular to elliptic–cordate, apically apiculate, or acute with central thickening. Its upper surface is more or less glandular or warty. The gynostemium is footless, rather short, clavate, somewhat sigmoid, the anther is suberect, the rostellum is short, suberect, and obtuse at the apex, and the ventral stigma is deeply concave. In all species of this group, inflorescence is few-flowered and usually longer than leaf, which is of course not a unique character, as it can be found elsewhere in Andinia.
Andinia spiralis and A. xenion have been classified to the genus Xenosia by Luer (2004), and A. longiserpens to Masdevalliantha by Szlachetko and Margońska [46]. Xenosia as originally described included three species, with Pleurothallis xenion as the generitype. Masdevalliantha included M. masdevalliopsis and M. longiserpens only. Xenosia as proposed by Luer [47] appears to be a paraphyletic taxon in the light of molecular analyses. This impression is intensified by morphology. In the case of this clade we have been dealt a kind of puzzle. The genus Xenosia can be characterized by a gynostemium structure, which is elongate, slender, clavate, slightly arcuate, with narrow apical clinandrium, apical, ventral anther, ligulate, incumbent rostellum, and short, baton-like column foot. This is the only mutual characteristic for the species of this genus, as they are different in other aspects of the flower structure. Inflorescence of X. spiralis–X. macrorhiza is a few-flowered raceme, longer than leaf, with obtuse sepals, lateral sepals connate almost to the apex, sagittate lip apically papillate, with oblong, central callus. Inflorescence of X. xenion is single-flowered, shorter than leaf, the sepals are caudate, free to the base, and the lip is basally 3-lobed with an oblong central callus. Its apical half is densely hairy. The gynostemium morphology and especially the connection between lip and column food recall the situation found in other Xenosia species. The common character of Masdevalliantha is a unique gynostemium morphology, which is short, massive, with long column foot and shelf-like, and massive process elevating rostellum. Especially interesting is the connection between lip and column foot, as the lip is attached on the ventral surface of the column foot, the apex of which is free and becoming accommodated by the basal lip cavity. We have not found such characteristics elsewhere in Andinia s.l. species. Flower segments are similar to those found in X. xenion, except lateral sepals, which in both Masdevalliantha species are basally shortly connate.
Analysis of the distribution of all samples in the climatic space was carried out, where Andinia, Aenigma, Masdevalliantha, and Lueranthos were separated. The distribution of all observations in the two-dimensional climatic space indicates that the samples are widespread across the whole climatic space defined by the annual precipitation (bio12) and the temperature seasonality (bio4).
The question arises, what is the sense in creating genera which although monophyletic in terms of sequence of two molecular markers, are impossible to define in terms of morphology? The partial answer can be found in Chase et al. [48]. The authors stated that their “general philosophy in developing the classification of Orchidaceae has been to minimize the number of tribes in order to make the system as simple as possible”. The next question is why they wish to minimize the number of tribes and other taxa? The answer is dumbfounding. They argued that “five is a reasonable number of subfamilies and is easily remembered by everyone”. This argument is difficult to challenge, but we dare to assume that if anyone is not able to remember six or seven subfamilies, that five is also too much to learn. Therefore Chase et al. [48] “have tended to reduce well-supported sister tribes to a single tribe”. We can add that even such taxa are ill-defined. From the other papers they published, we can guess that it is a general rule they try to implement into the orchid classification on various taxonomic levels and is followed by other authors. In this case, it is less important whether the group can be defined in terms of morphology or not. Much more important is whether they are monophyletic in terms of one or another molecular marker, and it is completely unimportant whether it is possible to propose a key to determination of taxa distinguished in such an odd manner. Moreover, Górniak et al. [39] was trying to prove that the topology of phylogenetic tree depends on the number of markers used in analyses and different taxonomic units could be proposed based on different tree topologies obtained.
To the best of our knowledge, the molecular marker used in analysis shows the phylogeny of this particular marker, and its extrapolation to the pattern of phylogeny of the entire organisms seems to be malfeasance. Furthermore, different markers can show different evolutionary paths which quite often are incongruent. In this case, which marker is more important?
Because all the above-mentioned morphologically uniform species groups form separated clades according to the results of phylogenetic analyses conducted by Wilson et al. [15], we propose to split Andinia s.l. into several genera, easily definable in terms of morphology and hence recognizable: Aenigma, Andinia, Chicalia, Lueranthos, Masdevalliantha, Minuscula, Neooreophilus, Xenosia, and Xenosiella. Most of these taxa were previously considered as valid genera, and we believe that there is no reason to lump them into a single ill-defined genus. The taxonomic decisions presented here were made primarily on the basis of morphology, with DNA data demonstrating congruence with the morphological distinctions.
4.3. Taxonomic Treatment
Key to genera of Andinia-complex
1. Gynostemium footless or foot rudimentary … 2
1* Gynostemium with prominent column-foot … 6
2. Plants pendent, leaves appearing alternating and often overlapping … 1. Neooreophilus
2* Plants ascending or caespitose, leaves not appearing alternating and overlapping … 3
3. Inflorescence single-flowered … 4. Chicalia
3* Inflorescence few-flowered … 4
4. Petals prominent … 2. Aenigma
4* Petals much smaller than sepals … 5
5. Lip erect, canaliculate, petals rudimentary, minutely papillate along margins, fused with the base of the column part and tightly adnate to it … 5. Lueranthos
5* Lip and petals not as above … 3. Andinia
6. Ovary muricate … 6. Minuscula
6* Ovary glabrous ... 7
7. Inflorescence few-flowered … 9. Xenosiella
7* Inflorescence single-flowered … 8
8. Lip motile, base with prominent cavity accommodating free apex of the column foot … 7. Masdevalliantha
8* Lip deeply 3-lobed at the base, firmly fixed on the apex of the column foot … 8. Xenosia
1. Neooreophilus Archila
Revista Guatemalensis 12(2): 73. 2009; Generitype: Neooreophilus nummularius (Rchb.f.) Archila [≡ Lepanthes nummularia Rchb. f.]. ≡ Lepanthes sect. Brachycladae Rchb. f., Xenia Orchid. 1: 142. 1856. ≡ Lepanthes subgen. Brachycladium Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 15: 31. 1986. ≡ Oreophilus W. E. Higgins & Archila, Revista Guatemal. 12(2): 73. 2009, nom. illeg. ≡ Penducella Luer & Thoerle, Orchid Digest 74(2): 68. 2010, nom. illeg.
Plants pendent, delicate, with rarely branching rhizome, rooting occasionally. Ramicaul obscure, terete, enclosed in 1–2 lepanthiform sheaths. Leaf much longer than ramicaul, elliptic to suborbicular, hispid to ciliate all over, all glabrous, with prominent venation. Few-flowered inflorescence usually shorter than leaf. Flowers typically lepanthiform, with sepals dissimilar or subsimilar in form and size, variously connate, lateral ones sometimes almost completely united together, more or less papillate or ciliate. Petals distinctly smaller than sepals, sometimes rudimentary, variously formed, longer than wide, or wider than long, entire or lobed, ciliate, papillate or with other protuberances. Lip shortly clawed to sessile, ecallose, with more or less auriculate base, lateral lobes upcurved, enveloping the gynostemium, ciliate or papillate. Gynostemium short, erect, terete, column foot missing, stigma subapical, horizontal, rostellum elongate, subulate or digitate, much exceeding the anther, erect or upcurved, anther erect, dorsal, ellipsoid, pollinia clavate, viscidium large, plate-like.
Three clearly separated assemblages in Neooreophilus can be distinguished based on morphological data and outcomes of molecular analyses, and these groups are presented here as subgenera. To facilitate identification of representatives within the largest subgenus, Amplectentes, this taxon was further divided into sections. However, it is important to emphasize the poor sampling of Neooreophilus that complicates comparison of morphological data with phylogenetic relationships. Based on similarity of related species, the more complicated division of the genus could be conducted based, for example, on petal form. More comprehensive molecular study is required to propose formal sectional division of the largest subgenus of Neooreophilus.
Key to subgenera of Neooreophilus
1. Inflorescence reclining, borne upon the leaf surface … 2
1* Inflorescence ascending, borne above the leaf surface … 1.1. Neooreophilus subgen. Amplectentes
2. Lip bilaminate with an appendix …. 1.2. Neooreophilus subgen. Bilamellatae
2* Lip transversely ovate to trapeziform in outline, without an appendix ... 1.3. Neooreophilus subgen. Neooreophilus
1.1. Subgenus Amplectentes (Luer) Kolan. & Szlach., stat. et comb. nov.
Basionym: Lepanthes sect. Amplectentes Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 52: 3. 1994. ≡ Andinia sect. Amplectentes (Luer) Karremans & S. Vieira-Uribe, Phytotaxa 295(2): 123. 2017. Type: Lepanthes pilosella Rchb. f., Flora 69: 556. 1886.
This is the largest subgenus of Neooreophilus. Three species groups can be distinguished within this subgenus.
Neooreophilus ciliaris-group
This group is characterized by glabrous leaves. Species included in this group:
- Neooreophilus ariasianus (Luer & L.Jost) Archila, Revista Guatemalensis 12(2): 73 (-74). 2009.
- Neooreophilus chaoae S.V.Uribe & L.Jost, Lankesteriana 15(3): 213. 2015.
- Neooreophilus chelosepalus (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 76. 2009.
- Neooreophilus ciliaris (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 76. 2009.
- Neooreophilus cordilabius (Luer) Archila, Revista Guatemalensis 12(2): 77. 2009.
- Neooreophilus dentatus Archila, Revista Guatemal., 17(1): 41. 2014.
- Neooreophilus destitutus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 78. 2009.
- Neooreophilus lunaris (Luer) Archila, Revista Guatemalensis 12(2): 81. 2009.
- Neooreophilus lynnianus (Luer) Archila, Revista Guatemalensis 12(2): 81. 2009.
- Neooreophilus macroticus (Luer & Dalström) Archila, Revista Guatemalensis 12(2): 82. 2009.
- Neooreophilus ortizianus S.V.Uribe & Thoerle, Orquideologia 28(2): 135 (-139; fig.; photogr.). 2011.
- Neooreophilus pendens (Garay) Archila, Revista Guatemalensis 12(2): 84. 2009.
- Neooreophilus pholeter (Luer) Archila, Revista Guatemalensis 12(2): 85.
- Neooreophilus ricii (Luer & R.Vásquez) Archila, Revista Guatemalensis 12(2): 86. 2009.
- Neooreophilus triangularis (Luer) Archila, Revista Guatemalensis 12(2): 87. 2009.
- Neooreophilus vieira-perezianus P.Ortiz, Orquideologia 28(1): 7 (-10; Figure 1; photogr.). 2011.
1.1.2. Neooreophilus pilosellus group
This group is characterized by hirsute or ciliate–denticulate leaves and cucullate dorsal sepal. It includes:
- Neooreophilus cardiochilus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 75. 2009.
- Neooreophilus chilopsis (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 76. 2009.
- Neooreophilus compositus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 77. 2009.
- Neooreophilus erepsis (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 78. 2009.
- Neooreophilus lunatocheillus Tobar & Archila, Revista Guatemalensis 15(2): 26, Figure 3. 2012.
- Neooreophilus mongei Tobar & Archila, Revista Guatemalensis 15(2): 23, Figure 2. 2012.
- Neooreophilus octocornutus (Luer) Archila, Revista Guatemalensis 12(2): 84. 2009.
- Neooreophilus persimilis (Luer & Sijm) Archila, Revista Guatemalensis 12(2): 84. 2009.
- Neooreophilus phalicus Tobar & Archila, Revista Guatemalensis 15(2): 20, Figure 1. 2012.
- Neooreophilus pilosellus (Rchb.f.) Archila, Revista Guatemalensis 12(2): 85. 2009.
- Neooreophilus platysepalus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 85. 2009.
- Neooreophilus tridactylus (Luer) Archila, Revista Guatemalensis 12(2): 88. 2009.
- Neooreophilus ursulus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 88. 2009.
- Neooreophilus viebrockianus (Luer & L.Jost) Archila, Revista Guatemalensis 12(2): 89. 2009.
- Neooreophilus villosus (Løjtnant) Archila, Revista Guatemalensis 12(2): 89. 2009.
Neooreophilus medinae Kolan. & Szlach., sp. nov. TYPE: Colombia. Dept. Putumayo. Vereda Balsayaco, near San Andres. Alt. 1984 m. 7 September 2016. R. Medina T. S14/28 (holotype, JAUM!; MEDEL!—photos).
[urn:lsid:ipni.org:names:xxxxxxx-x]
Species similar to N. platysepalus, N. ursulus, and N. pilosellus, distinguished by relatively shortly connate lateral sepals and prominent lip auricles. Moreover, from N. platysepalus and N. pilosellus, the new entity differs additionally by the lip and externally densely ciliate dorsal sepal.
Epiphytic plant, up to 15 cm long in total; ramicauls spaced 4–5 mm, each segment enclosed by two imbricating, membranaceous, infundibular sheaths, ciliate along the ribs, with dilated, ciliated ostia. Ramicauls 0.8–1.1 mm long, enclosed by a single membranaceous, infundibular sheath ciliate along the ribs, with dilated, ciliate ostia. Leaves shortly petiolate; petiole 0.7–1 mm long; blade 9–10 mm long, 3.5–4 mm wide, elliptic to ovate-elliptic, subacute to acute, densely ciliate, 3-nerved, lateral veins sometimes dichotomous. Inflorescence borne upon the surface of leaf, single-flowered; floral bract about 0.5 mm long, infundibular, oblique, acuminate, membranaceous; pedicel slender, persistent, 1.5–2 mm long; ovary ciliate, about 1 mm long. Flower small, sepals light yellow with dark red stripes, dorsal sepal mottled with purple along veins, petals light yellow with purple stripe, lip light yellow with 3 purple veins. Dorsal sepal concave in the natural position, 7.5 × 6 mm when expanded, broadly obovate, with a small, obtuse apiculus, 5- or 7-veined, externally densely ciliate. Lateral sepals 7 × 4 mm, connate for about 4 mm, narrowly ovate, obtuse, 2-veined, externally glabrous. Petals 2–2.1 × 0.4 mm, oblong–lanceolate to linear–lanceolate, basally cuneate, obtuse, 1-veined, glabrous. Lip embracing the column in the natural position, 1.7 × 1.5 mm when expanded, subsessile, lamina subrectangular with prominent basal projections 0.7 × 0.5 mm, apical part truncate with a small, obtuse apiculus and here glandular–ciliate, margin glabrous, disc 3-veined, glabrous. Gynostemium about 2 mm long, terete, anther and stigma apical (Figure 13 and Figure 14).
Etymology: Dedicated to Ramiro Medina Trejo, an orchid enthusiast who collected and cultivated new species.
Distribution, habitat and ecology: So far, Neooreophilus medinae is known only from the type locality in southern Colombia, where it was found on the mossy trunk and large branches of the tree in the pasture at the altitude of almost 2000 m. The population consists of about 50 specimens and its individuals grow sympatrically with specimens of N. pseudocaulescens. Flowering occurs in September.
Notes. Neooreophilus medinae resembles Colombian N. platysepalus, N. pilosellus, and N. ursulus, but the prominent basal projections of the lip allow one to easily distinguish it from the aforementioned species. In both N. pilosellus and N. ursulus, the lateral sepals are long-connate, free only at the apices. The most similar species is N. platysepalus known from the Colombian department of Antioquia, which is characterized by the glabrous sepals and small lip that is wider than long (1–1.25 × 1.5 mm), subquadrate to transversely subcordate, rounded to subtruncate at the apex, and minute basal projections. The lateral sepals of this species are connate in the basal 3/4–4/5.
Conservation status. Considering that about 50 specimens were found, the threat category according to IUCN [49] criterion D swings between Critically Endangered (CR) and Endangered (EN), while for the definition of the category according to criterion B data are missing on possible decline and fluctuations. Therefore, more in field research would be useful for the definition of distribution, population size, locations (sensu IUCN) and threats. In fact, new described species with a restricted distribution could be attributed to different IUCN categories, as Critically Endangered (CR) according to criterion B [50], or Vulnerable (VU) under criterion D [51,52].
1.1.3. Neooreophilus werneri-group
This group is characterized by hirsute or ciliate–denticulate leaves, not cucullate dorsal sepal, and variously connate, spreading lateral sepals. It includes:
- Neooreophilus catellus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 75. 2009.
- Neooreophilus dactylus (Garay) Archila, Revista Guatemalensis 12(2): 78. 2009.
- Neooreophilus exiguus (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 79. 2009.
- Neooreophilus geminipetalus (Luer & J.Portilla) Archila, Revista Guatemalensis 12(2): 79. 2009.
- Neooreophilus hippocrepicus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 80. 2009.
- Neooreophilus irrasus (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 80. 2009.
- Neooreophilus lueri (S.V.Uribe & Karremans) Kolan. & Szlach., comb. nov.
- Basionym: Andinia lueri S.V.Uribe & Karremans, Orquideologia 33: 116. 2016.
- Neooreophilus lupulus (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 81. 2009.
- Neooreophilus micropetalus (L.O.Williams) Archila, Revista Guatemalensis 12(2): 82. 2009.
- Neooreophilus monilius (Luer & R.Escobar) Archila, Revista Guatemalensis 12(2): 83. 2009.
- Neooreophilus montis-rotundus (P.Ortiz) Archila, Revista Guatemalensis 12(2): 83. 2009.
- Neooreophilus pseudocaulescens (L.B.Sm. & S.K.Harris) Archila, Revista Guatemalensis 12(2): 86. 2009.
- Neooreophilus rotundus Archila, Revista Guatemal. 17(1): 42. 2014.
- Neooreophilus sibundoyensis Kolan., Ann. Bot. Fenn. 50(3): 170. 2013.
- Neooreophilus sudamericanus Archila, Revista Guatemal., 17(1): 40. 2014.
- Neooreophilus werneri (Luer) Archila, Revista Guatemalensis 12(2): 89. 2009.
Incertæ sedis
Due to the lack of sufficient information about morphological characters of the following species we were not able to assign them into any of the groups listed above:
- Neooreophilus auriculatus Archila, Revista Guatemal. 17(1): 44. 2014.
- Neooreophilus caveroi (D.E. Benn. & Christenson) Archila, Revista Guatemal. 12(2): 76. 2009.
- Neooreophilus roseus Archila, Revista Guatemal. 17(1): 43. 2014.
1.2. Subgenus Bilamellatae (Luer) Kolan. & Szlach., stat. et comb. nov.
Basionym: Lepanthes sect. Bilamellatae Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 52: 3. 1994; Type: Lepanthes stalactites Luer & Hirtz, Lindleyana 2(2): 105. 1987.
This group is characterized by the inflorescence reclining, borne upon the leaf surface and bilaminate lip with an appendix.
- Neooreophilus stalactites (Luer & Hirtz) Archila, Revista Guatemalensis 12(2): 87. 2009.
1.3. Subgenus Neooreophilus
This group is characterized by the inflorescence reclining, borne upon the leaf surface and transversely ovate to trapeziform lip without an appendix.
- Neooreophilus nummularius (Rchb.f.) Archila, Revista Guatemalensis 12(2): 83. 2009.
- Neooreophilus obesus (S.V.Uribe & Karremans) Kolan. & Szlach., comb. nov.
Basionym: Andinia obesa S.V.Uribe & Karremans, Lankesteriana 17: 311.2017.
2. Aenigma (Luer) Szlach. & Kolan., stat. et comb. nov.
Basionym: Pleurothallis subgen. Aenigma Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 20: 26. 1986. ≡ Andinia subgen. Aenigma (Luer) Karremans & M. Wilson, Phytotaxa 295(2): 121. 2017. Generitype: Aenigma schizopogon (Luer). Szlach. & Kolan. [≡ Pleurothallis schizopogon Luer].
All species small, with somewhat elongate and ascending rhizome, producing ramicauls in short distance. Ramicaul short, terete, enveloped in 1 tight sheath. Leaf oblong, glabrous. Inflorescence longer than leaf, few-flowered, with flowers opening in succession. Ovary muricate, pedicel glabrous. Lateral sepals connate in the lower half, more or less acuminate, glabrous, or hispid. Petals prominent, linear–lanceolate, acuminate, as long as sepals, but distinctly narrower. Lip sessile, firmly connate with the column foot, more or less reflexed just above the base, enveloping the gynostemium, with prominent calli in the center, suborbicular to pentagonal in outline, shortly apiculate in the apex. Gynostemium erect to somewhat arcuate, clavate, column foot rudimentary, stigma subapical, rostellum blunt, short, rounded at the apex, anther subapical to subventral, suberect, ovoid–ellipsoid. The genus includes:
- Aenigma dalstroemii (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis dalstroemii Luer, Orchideer 5: 52. 1984.
- Aenigma hirtzii (Luer) Szlach. & Kolan., comb. nov.
Basionym: Andinia hirtzii Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 103: 275. 2005.
- Aenigma pogonion (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis pogonion Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 52: 61. 1994.
- Aenigma schizopogon (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis schizopogon Luer, Lindleyana 16(4): 251. 2001.
- Aenigma uchucayensis (A.Doucette & J.Portilla) Szlach. & Kolan., comb. nov.
Basionym: Andinia uchucayensis A.Doucette & J.Portilla, Orchids (Lindleyana) 86(1): 72. 2017.
3. Andinia (Luer) Luer
Monogr. Syst. Bot. Missouri Bot. Gard. 79: 5. 2000. ≡ Salpistele subgen. Andinia Luer, (Monogr. Syst. Bot. Missouri Bot. Gard. 39: 124. 1991; Generitype: Andinia dielsii (Mansf.) Luer [≡ Lepanthes dielsii Mansf.].
Plants with elongate rhizome, producing ramicauls in prominent intervals. Ramicaul short, terete, enclosed in 1–2 tight sheaths. Leaf oblong elliptic, glabrous. Multiflowered inflorescence much longer than leaf. The flowers produced in succession, broadly opened. Ovary muricate, or glabrous, pedicellate. Sepals subsimilar in size and form, shortly acuminate, mucronate, ciliate, or glabrous along margins, lateral sepals connate in the lower half or free to the base. Petals much smaller than sepals. Lip simple, 3-lobed, basally or subapically united with the gynostemium, lateral lobes upcurved, enveloping the gynostemium, with prominent callus in the center or ecallose. Gynostemium erect, slender, terete, column foot missing, stigma ventral, emarginate, anther subdorsal, erect, ovoid, rostellum elongate, ligulate, erect, much exceeding the anther.
The genus includes:
- Andinia dielsii (Mansf.) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 79: 6. 2000.
- Andinia pensilis (Schltr.) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 79: 6. 2000.
- Andinia tingomariana A.G. Diaz & Mark Wilson, Phytotaxa 361(2): 223–227. 2018.
4. Chicalia Szlach. & Kolan., gen. nov.
Generitype: Chicalia lappacea (Luer) Szlach. & Kolan. [≡ Pleurothallis lappacea Luer]
Etymology: In reference to Chical, Carchi, Ecuador, where the type specimen of P. lap-pacea was collected.
Plants very small, subcaespitose. Ramicaul obscure, enclosed in 1 tight sheath. Leaf elliptic, glabrous. Single-flowered inflorescence longer than leaf, produces relatively large, broadly opened flower. Ovary densely muricate, pedicellate. Sepals subsimilar in size, papillate all over except central and basal parts, lateral sepals connate together almost to the apex forming a kind of platform, supporting lip. Petals rudimentary, with ciliate margins. Lip sessile, subquadrate in outline, reflexed just above the base, enveloping partially the gynostemium, with prominently emarginate apex. Its surface ciliatae–glandular, except the central and apical portions. Gynostemium rather short, clavate, subsigmoid, with obscure column foot, anther apical, incubment ellipsoid, rostellum subapical, short, obscure, stigma subapical.
The genus includes:
- Chicalia lappacea (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis lappacea Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 79: 129, f. 5. 2000.
5. Lueranthos Szlach. & Marg.
Polish Bot. J. 46(2): 117. 2001 [2002]; Generitype: Lueranthos vestigipetalus (Luer) Szlach. & Marg. [≡ Pleurothallis vestigipetala Luer].
Plants with elongate, ascending rhizome. Ramicaul short, terete, enclosed in tight sheath. Inflorescence few-flowered, somewhat longer than leaf. Flowers produced in succession with glabrous ovary and broadly opened sepals free to the base. Sepals similar in size and form. Petals rudimentary, minutely papillate along margins, fused with the base of the column part and tightly adnate to it. Lip erect, canaliculate, oblong–elliptic, ecallose, simple, more or less papillate all over. Gynostemium erect, terete, slender, elongate, footless, with subapical, incumbent anther and short rostellum. Anther oblong ovoid. Stigma apical, horizontal.
This monotypic taxon includes:
- Lueranthos vestigipetalus (Luer) Szlach. & Marg., Polish Bot. J. 46(2): 117. 2001 [2002].
6. Minuscula Szlach. & Kolan., gen. nov.
Generitype: Minuscula trimytera (Luer & R.Escobar) Szlach. & Kolan. [≡ Pleurothallis trimytera Luer & R. Escobar].
Etymology: From minusculus (Lat.)—small, tiny. An allusion to the size of species of this group.
Plants miniature, with very short, creeping rhizome or caespitose. Ramicaul obscure, terete, enclosed in 1 tight sheath. Leaf elliptic, shortly apiculate, glabrous. The inflorescence longer than leaf; 1-(2)-flowered. Flower broadly opened. Ovary muricate, pedicellate. Sepals subsimilar in form and size, glabrous, papillate or glandular in the basal part or all over, lateral sepals connate in the lower half, occasionally free. Petals shorter and narrower than sepals, occasionally rudimentary. Lip shortly clawed, transversely elliptic, deeply 3-lobed at the apex, geniculately bent forward, thickened in the center, concave below, rarely calli obscure. Gynostemium slightly sigmoid, clavate, column foot short, apical clinandrium obscure, anther apical, incumbent, oblong ovoid, stigma ventral, rostellum ventral, triangular, more or less incumbent.
The genus includes:
- Minuscula hystricosa (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis hystricosa Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 52: 54. 1994.
- Minuscula ibex (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis ibex Luer, Selbyana 5(2): 168 (–169). 1979.
- Minuscula panica (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis panica Luer & Dalström, Monogr. Syst. Bot. Missouri Bot. Gard. 61(3): 6. 1996.
- Minuscula pentamytera (Luer) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis pentamytera Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 52: 58. 1994.
- Minuscula sunchubambensis (A. Doucette & Janovec) Szlach. & Kolan., comb. nov.
Basionym: Andinia sunchubambensis A. Doucette & Janovec, Internet Orchid Sp. Photo Encycl. Nomencl. Notes 2016: f. 1A–G, 2–3. 2016.
- Minuscula trimytera (Luer & R.Escobar) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis trimytera Luer & R. Escobar, Orquideología 16: 34. 1983.
7. Masdevalliantha (Luer) Szlach. & Marg.
Polish Bot. J. 46(2): 117. 2001. ≡ Pleurothallis subgen. Masdevalliantha Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 20: 44. 1986. ≡ Andinia subgen. Masdevalliantha (Luer) Karremans & M. Wilson, Phytotaxa 295(2): 125. 2017; Generitype: Masdevalliantha masdevalliopsis (Luer) Szlach. & Marg. [≡ Pleurothallis masdevalliopsis Luer].
Subcaespitose plant, with 1-flowered inflorescence longer than leaf. Ramicaul short, terete, with tight sheath. Flowers broadly opened. Sepals subsimilar in form and size, long–caudate, lateral sepals connate basally. Petals orbicular to obtriangular, much smaller than sepals. Lip motile, more or less 3-lobed, base with prominent cavity accommodating free apex of the column foot, apical part of the middle lobe hairy or hispid, callus single, oblong, or bifid. Gynostemium short, massive, suberect, rostellum elongate, oblong–triangular, perpendicular to the column axis, column foot long, upcurved with the lip attached below its apex, which is placed in the cavity at the lip base. Anther subapical, suberect, ovoid, relatively small. Stigma ventral.
The genus includes:
- Masdevalliantha longiserpens (C. Schweinf.) Szlach. & Marg., Polish Bot. J. 46(2): 117. 2001.
- Masdevalliantha masdevalliopsis (Luer) Szlach. & Marg., Polish Bot. J. 46(2): 117. 2001.
8. Xenosia Luer
Monogr. Syst. Bot. Missouri Bot. Gard. 95: 265. 2004; Generitype: Xenosia xenion (Luer & R. Escobar) Luer [≡ Pleurothallis xenion Luer & R. Escobar].
Etymology: In reference to the similarity to the genus Xenosia.
Plants miniature. Rhizome shortly repent. Ramicaul filiform, tightly enclosed in 1 sheath. Leaf linear–lanceolate, glabrous. Inflorescence shorter than leaf, single-flowered. Flowers broadly opened. Ovary glabrous, pedicellate. Sepals subsimilar in form and size, free to the base, long–caudate. Petals deltoid, prominently smaller than sepals. Lip deeply 3-lobed at the base, firmly fixed on the apex of the column foot, with basal lateral lobes obliquely rhombic, and oblong callus in the center, the middle lobe densely pubescent. Gynostemium slender, elongate, terete, apically somewhat arcuate. Apical clinandrium narrow. Column foot elongate, robust. Anther apical, incumbent, ovoid. Rostellum ventral, ligulate, incumbent. Stigma ventral.
The genus includes one species:
- Xenosia xenion (Luer & R. Escobar) Luer, Monogr. Syst. Bot. Missouri Bot. Gard. 95: 265. 2004.
9. Xenosiella Szlach. & Kolan., gen nov.
Generitype: Xenosiella spiralis (Ruiz & Pav.) Szlach. & Kolan. [≡ Humboltia spiralis Ruiz & Pav.]
Plants with elongate, ascending rhizome. Pseudobulbs terete, short, enclosed in tight sheath. Leaf ligulate, glabrous. Inflorescence shorter than leaf, few-flowered. Flowers tubular borne in succession. Ovary glabrous, pedicellate. Sepals subsimilar in size and form, obtuse, glabrous, lateral sepals connate together in the basal 2/3, forming a kind of platform below lip. Petals oblong–obovate, much smaller than sepals. Lip firmly attached to the column foot, deltoid–ovate in outline, canaliculate, obscurely 3-lobed, with oblong callus in the center and papillate apical lobe. Gynostemium elongate, slender, clavate, with prominent, slightly upcurved column foot. Anther ventral, ellipsoid, pollinia 2, ovoid. Rostellum ligulate, obtuse, incumbent. Viscidium plate-like. Stigma ventral.
The two species classified in this genus are probably synonymic as the only noticeable difference between them is the lip base (cuneate in X. spiralis and rounded-broadly cuneate in X. macrorhiza). The genus includes:
- Xenosiella macrorhiza (Lindl.) Szlach. & Kolan., comb. nov.
Basionym: Pleurothallis macrorhiza Lindl., Monogr. Syst. Bot. Missouri Bot. Gard. 105: 233–234, f. 185. 2006.
- Xenosiella spiralis (Ruiz & Pav.) Szlach. & Kolan., comb. nov.
Basionym: Humboltia spiralis Ruiz & Pav., Syst. Veg. Fl. Peruv. Chil. 1: 237. 1798.
5. Conclusions
Molecular analyses clearly show a monophyly of the genus Andinia s.l. The genus is intangible, however, in morphological terms. High diversity in the flower and gynostemium structure can be interpreted as an adaptation to various pollination systems. We do not have sufficient information on pollination strategy of the species of the Andinia-complex, but we can assume that multifarious structure of the flowers can be attractive to different species of insects. The differences in the temperature seasonality, the annual precipitation, and the precipitation of the warmest and the coldest quarters, which differentiate species of the Andinia-complex, can also affect a species composition of potential pollinators. In our opinion, separation of monophyletic, smaller, but morphologically well-supported genera is unreservedly legitimate. It appeared that they represents closely related taxa which were derived from a common ancestor. Unfortunately, it is difficult to define the relationship between these groups. It is likely that each evolved independently from the other. Perhaps this was due to climatic factors or the result of adaptation to different habitats.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14050372/s1, Annex S1: Morphological characters of Andinia-complex representatives; Annex S2: List of localities of Andinia-complex representatives with bioclims values for each record; Annex S3: Ecoregions corresponding to localities of Andinia-complex representatives.
Author Contributions
Conceptualization, D.L.S. and M.K.; methodology, D.L.S., M.D., A.N. and M.K.; software, M.D., A.N. and M.K.; validation, D.L.S. and M.K. formal analysis, D.L.S., M.D., A.N. and M.K.; investigation, D.L.S., M.D., A.N. and M.K.; resources, D.L.S. and M.K.; data curation, D.L.S., M.D., A.N. and M.K.; writing—original draft preparation, D.L.S., M.D., A.N. and M.K.; writing—review and editing, D.L.S., M.D., A.N. and M.K.; visualization, M.D., A.N. and M.K.; supervision, D.L.S. and M.K. project administration, D.L.S. and M.K.; funding acquisition, D.L.S. and M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education, Youth, and Sports of CR within the CzeCOS program, grant number LM2018123.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article and supplementary material.
Acknowledgments
The curators and staff of the cited herbaria are thanked for their kind hospitality and assistance during visits. We are grateful to José “Pepe” Portilla, Ivan Portilla, Mario Portilla, Magali Portilla, and Iván Acaro for access to their greenhouses and assistance during our visits. We would like to thank Ramiro Medina for providing data from his orchid cultivation and to Alexander Hirtz for assistance during excursions to Ecuador. We are grateful to Natalia Olędrzyńska for preparing a line drawing of a new species.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Luer, C.A. Icones Pleurothallidinarum I: Systematics of the Pleurothallidinae (Orchidaceae). Monogr. Syst. Bot. Mo. Bot. Gard. 1986, 15, 1–81. [Google Scholar]
- Karremans, A.P. Genera Pleurothallidinarum: An updated phylogenetic overview of the Pleurothallidinae. Lankesteriana 2016, 16, 219–241. [Google Scholar] [CrossRef] [Green Version]
- Pridgeon, A.M.; Solano, R.; Chase, M.W. Phylogenetic relationships in Pleurothallidinae (Orchidaceae): Combined evidence from nuclear and plastid DNA sequences. Am. J. Bot. 2001, 88, 2286–2308. [Google Scholar] [CrossRef] [PubMed]
- Pridgeon, A.M.; Chase, M.W. A phylogenetic reclassification of Pleurothallidinae (Orchidaceae). Lindleyana 2001, 16, 235–271. [Google Scholar]
- Higgins, W.E. Pleurothallidinae: How many genera. In Proceedings of the 19th World Orchid Conference, Miami, FL, USA, 23–27 January 2008; pp. 425–430. [Google Scholar]
- Barros, F.; Barberena, F.F.V.A. Nomenclatural notes and new combinations on Anathallis and Specklinia (Orchidaceae). Rodriguésia 2010, 61, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Karremans, A. Nomenclatural notes in the Pleurothallidinae (Orchidaceae): Stelis. Phytotaxa 2015, 203, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Karremans, A.; Bogarin, D.; Pupulin, F.; Luer, C.; Gravendeel, B. The glandulous Specklinia: Morphological convergence versus phylogenetic divergence. Phytotaxa 2015, 218, 101–127. [Google Scholar] [CrossRef]
- Karremans, A.; Rincon-Gonzalez, M. Nomenclatural notes in the Pleurothallidinae (Orchidaceae): Apoda-prorepentia. Phytotaxa 2015, 238, 174–182. [Google Scholar] [CrossRef]
- Pupulin, F.; Karremans, A.; Gravendeel, B. A reconsideration of the empusellous species of Specklinia (Orchidaceae: Pleurothallidinae) in Costa Rica. Phytotaxa 2012, 63, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bogarin, D.; Perez-Escobar, O.A.; Groenenberg, D.; Holland, S.D.; Karremans, A.P.; Lemmon, E.M.; Lemmon, A.R.; Pupulin, F.; Smets, E.; Gravendeel, B. Anchored hybrid enrichment generated nuclear, plastid and mitochondrial markers resolve the Lepanthes horrida (Orchidaceae: Pleurothallidinae) species complex. Mol. Phylogenet. Evol. 2018, 129, 27–47. [Google Scholar] [CrossRef]
- Luer, C.A. Icones Pleurothallidinarum XXVII. Miscellaneous new taxa and combinations. Monogr. Syst. Bot. Mo. Bot. Gard. 2005, 103, 307–309. [Google Scholar]
- Archila, F.; Higgins, W.E. Oreophilus: A new genus in the Pleurothallidinae Lindl. (Orchidaceae). Selbyana 2008, 29, 202–208. [Google Scholar]
- Archila, F. Neooreophilus Archila: A new genus in the Pleurothallidinae Lindl. (Orchidaceae). Rev. Guatemalensis 2009, 12, 71–90. [Google Scholar]
- Wilson, M.; Frank, G.; Jost, L.; Pridgeon, A.; Vieira-Uribe, S.; Karremans, A. Phylogenetic analysis of Andinia (Pleurothallidinae; Orchidaceae) and a systematic re-circumscription of the genus. Phytotaxa 2017, 295, 101–131. [Google Scholar] [CrossRef]
- Stepanović, S.; Kosovac, A.; Krstić, O.; Jović, J.; Toševski, I. Morphology versus DNA barcoding: Two sides of the same coin. A case study of Ceutorhynchus erysimi and C. contractus identification. Insect Sci. 2016, 23, 638–648. [Google Scholar] [CrossRef] [Green Version]
- Rejlová, L.; Böhmová, A.; Chumová, Z.; Hořčicová, Š.; Josefiová, J.; Schmidt, P.-A.; Trávníček, P.; Urfus, T.; Vít, P.; Chrtek, J. Disparity between morphology and genetics in Urtica dioica (Urticaceae). Bot. J. Linn. Soc. 2020, 195, 606–621. [Google Scholar] [CrossRef]
- Wiens, J.J. The Role of Morphological Data in Phylogeny Reconstruction. Syst. Biol. 2004, 53, 653–661. [Google Scholar] [CrossRef]
- Hills, D.M.; Wiens, J.J. Molecules versus morphology in systematics: Conflicts, artifacts, and misconceptions. In Phylogenetic Analysis of Morphological Data; Wiens, J.J., Ed.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 1–19. [Google Scholar]
- Galtier, N.; Gouy, M.; Gautier, C. SEAVIEW and PHYLO_WIN: Two graphic tools for sequence alignment and molecular phylogeny. Comput. Appl. Biosci. 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kores, P.; Molvray, M.; Weston, P.; Hopper, S.; Brown, A.; Cameron, K.; Chase, M. A phylogenetic analysis of Diurideae (Orchidaceae) based on plastid DNA sequence data. Am. J. Bot. 2001, 88, 1903–1914. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Huelsenbeck, J. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A. MrModeltest V2; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A.; Drummond, A.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian Phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. 2021. Available online: http://sweetgum.nybg.org/science/ih/ (accessed on 1 June 2021).
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth: A new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Fick, S.; Hijmans, R. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Pichardo-Marcano, F.J.; Nieto-Blázquez, M.E.; MacDonald, A.N.; Galeano, G.; Roncal, J. Phylogeny, historical biogeography and diversification rates in an economically important group of Neotropical palms: Tribe Euterpeae. Mol. Phylogenet. Evol. 2019, 133, 67–81. [Google Scholar] [CrossRef]
- Koutroumpa, K.; Theodoridis, S.; Warren, B.H.; Jiménez, A.; Celep, F.; Doğan, M.; Romeiras, M.M.; Santos-Guerra, A.; Fernández-Palacios, J.M.; Caujapé-Castells, J.; et al. An expanded molecular phylogeny of Plumbaginaceae, with emphasis on Limonium (sea lavenders): Taxonomic implications and biogeographic considerations. Ecol. Evol. 2018, 8, 12397–12424. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Tang, C.C.; Thomas, D.C.; Couvreur, T.L.P.; Saunders, R.M.K. A mega-phylogeny of the Annonaceae: Taxonomic placement of five enigmatic genera and support for a new tribe, Phoenicantheae. Sci. Rep. 2017, 7, 7323. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, E.; Bruneau, A.; Hughes, C.E.; de Queiroz, L.P.; Lewis, G.P. A new generic system for the pantropical Caesalpinia group (Leguminosae). PhytoKeys 2016, 71, 1–160. [Google Scholar] [CrossRef]
- Xiang, X.G.; Hu, H.; Wang, W.; Jin, X.H. DNA barcoding of the recently evolved genus Holcoglossum (Orchidaceae: Aeridinae): A test of DNA barcode candidates. Mol. Ecol. Resour. 2011, 11, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Raskoti, B.B.; Ale, R. DNA barcoding of medicinal orchids in Asia. Sci. Rep. 2021, 11, 23651. [Google Scholar] [CrossRef] [PubMed]
- Parveen, I.; Singh, H.; Malik, S.; Raghuvanshi, S.; Babbar, S. Evaluating five different loci (rbcL, rpoB, rpoC1, matK, and ITS) for DNA barcoding of Indian orchids. Genome 2017, 60, 665–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Soltis, D.E.; Soltis, P.S.; Zhu, X.; Burleigh, J.G.; Chen, Z. Deep phylogenetic incongruence in the angiosperm clade Rosidae. Mol. Phylogenet. Evol. 2015, 83, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Górniak, M.; Szlachetko, D.L.; Olędrzyńska, N.; Naczk, A.M.; Mieszkowska, A.; Boss, L.; Ziętara, M.S. Species Phylogeny versus Gene Trees: A Case Study of an Incongruent Data Matrix Based on. Int. J. Mol. Sci. 2021, 22, 11393. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Ellstrand, N.C.; Arnold, M. What Can Molecular and Morphological Markers Tell Us About Plant Hybridization? Crit. Rev. Plant Sci. 1993, 12, 213–241. [Google Scholar]
- Kolanowska, M.; Szlachetko, D.L. Notes on Erycina-complex with descriptions of new Colombian species. Plant Syst. Evol. 2013, 300, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Szlachetko, D.L.; Kolanowska, M. Generic delimitation of the genera Brevilongium and Otoglossum (Orchidaceae) with description of new species from Colombia and Panama. Plant Biosyst. 2014, 148, 1148–1159. [Google Scholar] [CrossRef]
- Kolanowska, M.; Szlachetko, D. Problems with generic delimitation in the Odontoglossum complex (Orchidaceae, Oncidiinae) and an attempt for a solution. Plant Syst. Evol. 2016, 302, 203–217. [Google Scholar] [CrossRef]
- Karremans, A.P. To be, or not to be a Stelis. Lankesteriana 2019, 19, 281–343. [Google Scholar] [CrossRef]
- Favre, F.; Jourda, C.; Besse, P.; Charron, C. Genotyping-by-Sequencing Technology in Plant Taxonomy and Phylogeny. In Molecular Plant Taxonomy. Methods in Molecular Biology; Besse, P., Ed.; Humana: New York, NY, USA, 2021; pp. 167–178. [Google Scholar]
- Szlachetko, D.L.; Margońska, H.B. Genera et species orchidalium 3. Pol. Bot. J. 2001, 46, 113–121. [Google Scholar]
- Luer, C.A. Icones Pleurothallidinarum XXVI. Pleurothallis subgenus Acianthera and three allied subgenera. A second century of new species of Stelis of Ecuador. Epibator, Ophidion, Zootrophion. Addenda to Brachionidium, Dracula, Lepanthes, Platystele, Pleurothallis, Porroglossum, and Masdevallia. New genera and combinations. Monogr. Syst. Bot. Mo. Bot. Gard. 2004, 95, 1–265. [Google Scholar]
- Chase, M.; Cameron, K.; Freudenstein, J.; Pridgeon, A.; Salazar, G.; Van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef] [Green Version]
- International Union for Conservation of Nature (IUCN). Guidelines for Using the IUCN Red List Categories and Criteria. Version 15. Available online: https://nc.iucnredlist.org/redlist/content/attachment_files/RedListGuidelines.pdf (accessed on 23 March 2022).
- Wagensommer, R.P.; Venanzoni, R. Geranium lucarinii sp. nov. and re-evaluation of G. kikianum (Geraniaceae). Phytotaxa 2021, 489, 252–262. [Google Scholar] [CrossRef]
- Blasco, F.A.; Rubite, R.R.; Cortes, J.C.; Alejandro, G.J.D. Begonia lanuzaensis (sect. Petermannia, Begoniaceae) a new species from Surigao del Sur, Mindanao Island, Philippines. Phytotaxa 2021, 523, 203–207. [Google Scholar] [CrossRef]
- Swanepoel, W.; Cauwer, V.d.; Van Wyk, A.E.A. A new rheophytic species of Syzygium (Myrtaceae) from the lower Kunene River of Angola and Namibia. Phytotaxa 2021, 491, 281–290. [Google Scholar] [CrossRef]
Figure 1.
Distribution of studied genera. Base map of ecoregions developed by Olson [28].
Figure 1.
Distribution of studied genera. Base map of ecoregions developed by Olson [28].

Figure 2.
Majority-rule consensus tree for Andinia s.l. obtained in Bayesian analysis from ITS1-5.8S-ITS2 sequences. Values above branches represent posteriori probabilities; values with pp < 0.9 are indicated with an asterisk. The main and strong supported clades on the tree are marked with the numbers I–VI. Discussed groups of species are marked as A–C2.
Figure 2.
Majority-rule consensus tree for Andinia s.l. obtained in Bayesian analysis from ITS1-5.8S-ITS2 sequences. Values above branches represent posteriori probabilities; values with pp < 0.9 are indicated with an asterisk. The main and strong supported clades on the tree are marked with the numbers I–VI. Discussed groups of species are marked as A–C2.

Figure 3.
Phylogenetic tree performed based on maximum likelihood analysis inferred from the ITS matrix using ML/Transfer bootstrap expectation with 1000 bootstrap replicates in RaxML. Values at each node represent percent bootstrap support; bootstrap percentages less than 50% are not shown. The main and strong supported clades on the tree are marked with the numbers I–IV. However, the subclades are tagged as a and b. The main and strong supported clades on the tree are marked with the numbers I–IV. Discussed groups of species are marked as A–C2.
Figure 3.
Phylogenetic tree performed based on maximum likelihood analysis inferred from the ITS matrix using ML/Transfer bootstrap expectation with 1000 bootstrap replicates in RaxML. Values at each node represent percent bootstrap support; bootstrap percentages less than 50% are not shown. The main and strong supported clades on the tree are marked with the numbers I–IV. However, the subclades are tagged as a and b. The main and strong supported clades on the tree are marked with the numbers I–IV. Discussed groups of species are marked as A–C2.

Figure 4.
Majority-rule consensus tree obtained in Bayesian analysis of part of matK gene for Andinia s.l. Values above branches represent posteriori probabilities; values with pp < 0.9 are indicated with an asterisk. The main and strong supported clades on the tree are marked with the numbers I–IX. Discussed groups of species are marked as A–C2.
Figure 4.
Majority-rule consensus tree obtained in Bayesian analysis of part of matK gene for Andinia s.l. Values above branches represent posteriori probabilities; values with pp < 0.9 are indicated with an asterisk. The main and strong supported clades on the tree are marked with the numbers I–IX. Discussed groups of species are marked as A–C2.

Figure 5.
Phylogenetic tree performed based on maximum likelihood analysis inferred from the matK matrix using ML/Transfer bootstrap expectation with 1000 bootstrap replicates in RaxML. Values at each node represent percent bootstrap support; bootstrap percentages less than 50% are not shown. The main and strong supported clades on the tree are marked with the numbers I–III. The subclades are tagged as 1–6. Discussed groups of species are marked as A–C1.
Figure 5.
Phylogenetic tree performed based on maximum likelihood analysis inferred from the matK matrix using ML/Transfer bootstrap expectation with 1000 bootstrap replicates in RaxML. Values at each node represent percent bootstrap support; bootstrap percentages less than 50% are not shown. The main and strong supported clades on the tree are marked with the numbers I–III. The subclades are tagged as 1–6. Discussed groups of species are marked as A–C1.

Figure 6.
Majority-rule consensus tree for Andinia s.l. obtained in Bayesian analysis from combined matrix (ITS + matK). Values above branches represent posteriori probabilities; values with pp < 0.9 are indicated with an asterisk. The main and strong supported clades on the tree are marked with the numbers I–III. The subclades are tagged as capitals A–C. Discussed groups of species are marked as A–C2.
Figure 6.
Majority-rule consensus tree for Andinia s.l. obtained in Bayesian analysis from combined matrix (ITS + matK). Values above branches represent posteriori probabilities; values with pp < 0.9 are indicated with an asterisk. The main and strong supported clades on the tree are marked with the numbers I–III. The subclades are tagged as capitals A–C. Discussed groups of species are marked as A–C2.

Figure 7.
Phylogenetic tree performed based on maximum likelihood analysis inferred from the combined ITS-matK dataset using ML/Transfer bootstrap expectation with 1000 bootstrap replicates in RaxML. Values at each node represent percent bootstrap support; bootstrap percentages less than 50% are not shown. The main and strong supported clades on the tree are marked with the numbers I–III. However, the subclades are tagged as capitals A–D. Discussed groups of species are marked as A–C2.
Figure 7.
Phylogenetic tree performed based on maximum likelihood analysis inferred from the combined ITS-matK dataset using ML/Transfer bootstrap expectation with 1000 bootstrap replicates in RaxML. Values at each node represent percent bootstrap support; bootstrap percentages less than 50% are not shown. The main and strong supported clades on the tree are marked with the numbers I–III. However, the subclades are tagged as capitals A–D. Discussed groups of species are marked as A–C2.

Figure 8.
Principal component analysis (PCA) of the climatic variables present in the studied genera among the Andinia-complex. Variables with the greatest contributions are presented as vectors.
Figure 8.
Principal component analysis (PCA) of the climatic variables present in the studied genera among the Andinia-complex. Variables with the greatest contributions are presented as vectors.

Figure 9.
Canonical variate analysis (CVA) of the studied genera among the Andinia-complex along the climatic gradient. The relative contributions of bioclimatic factors are shown as vectors. Variables with low discriminating impact are omitted.
Figure 9.
Canonical variate analysis (CVA) of the studied genera among the Andinia-complex along the climatic gradient. The relative contributions of bioclimatic factors are shown as vectors. Variables with low discriminating impact are omitted.

Figure 10.
Principal component analysis (PCA) of the climatic variables present in the studied species. Variables with the greatest contributions are presented as vectors.
Figure 10.
Principal component analysis (PCA) of the climatic variables present in the studied species. Variables with the greatest contributions are presented as vectors.

Figure 11.
Canonical variate analysis (CVA) of the studied species along the climatic gradient. The relative contributions of bioclimatic factors are shown as vectors. Variables with low discriminating impact are omitted.
Figure 11.
Canonical variate analysis (CVA) of the studied species along the climatic gradient. The relative contributions of bioclimatic factors are shown as vectors. Variables with low discriminating impact are omitted.

Figure 12.
The cluster analysis UPGMA of the studied biogeographical ecoregions in terms of bioclimatic variables.
Figure 12.
The cluster analysis UPGMA of the studied biogeographical ecoregions in terms of bioclimatic variables.

Figure 13.
Neooreophilus medinae Kolan. & Szlach.—dissected perianth. (A)—sepals, (B)—dorsal sepal, (C)—petal, (D)—lip, (E)—gynostemium. Drawn by N. Olędrzyńska from the holotype.
Figure 13.
Neooreophilus medinae Kolan. & Szlach.—dissected perianth. (A)—sepals, (B)—dorsal sepal, (C)—petal, (D)—lip, (E)—gynostemium. Drawn by N. Olędrzyńska from the holotype.

Figure 14.
Neooreophilus medinae Kolan. & Szlach.—plant in situ. (A)—plant habit, (B)—plants hanging on a branch covered by mosses in natural habitat.
Figure 14.
Neooreophilus medinae Kolan. & Szlach.—plant in situ. (A)—plant habit, (B)—plants hanging on a branch covered by mosses in natural habitat.


{kind=link}
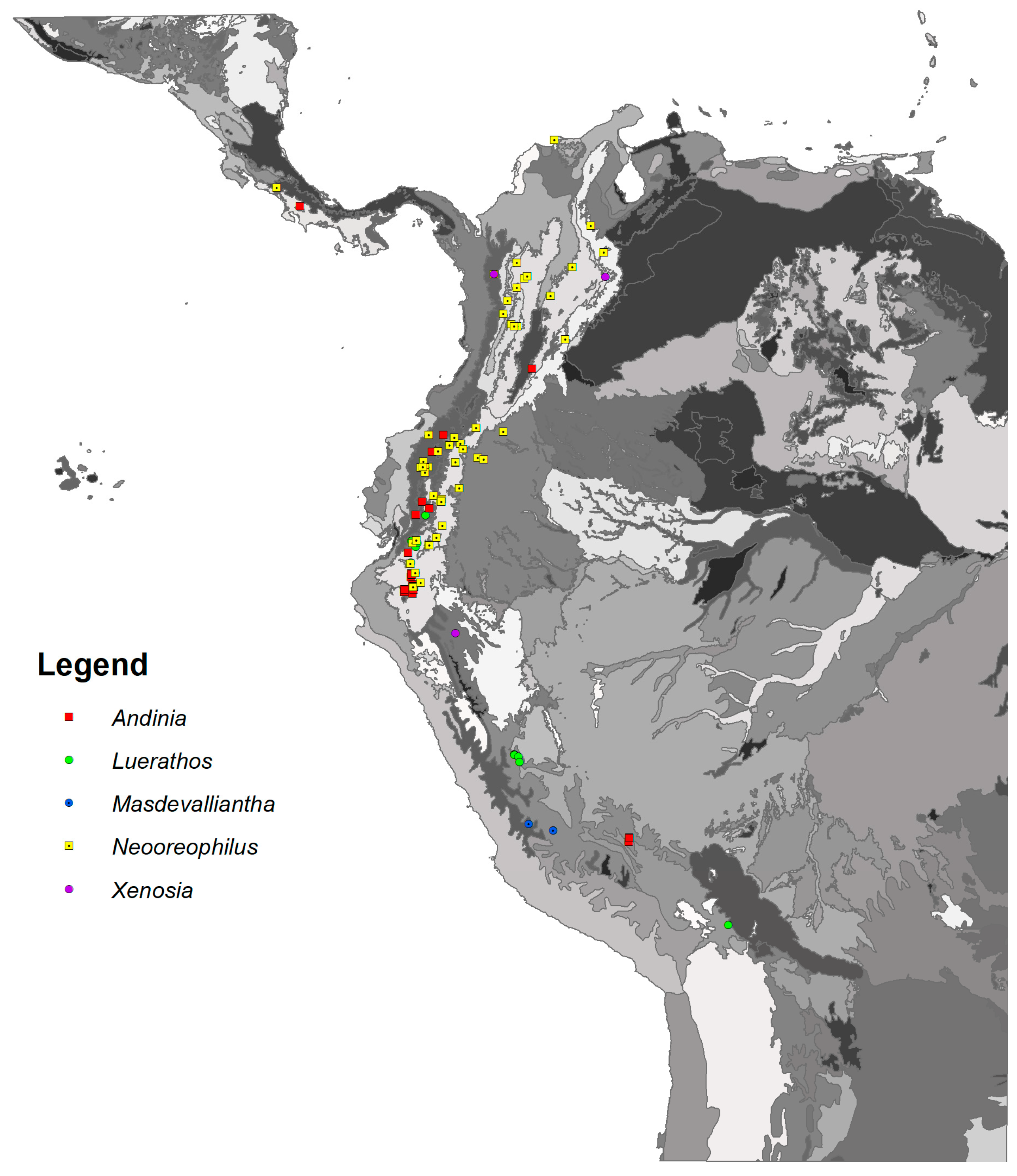{kind=link}
{kind=link}
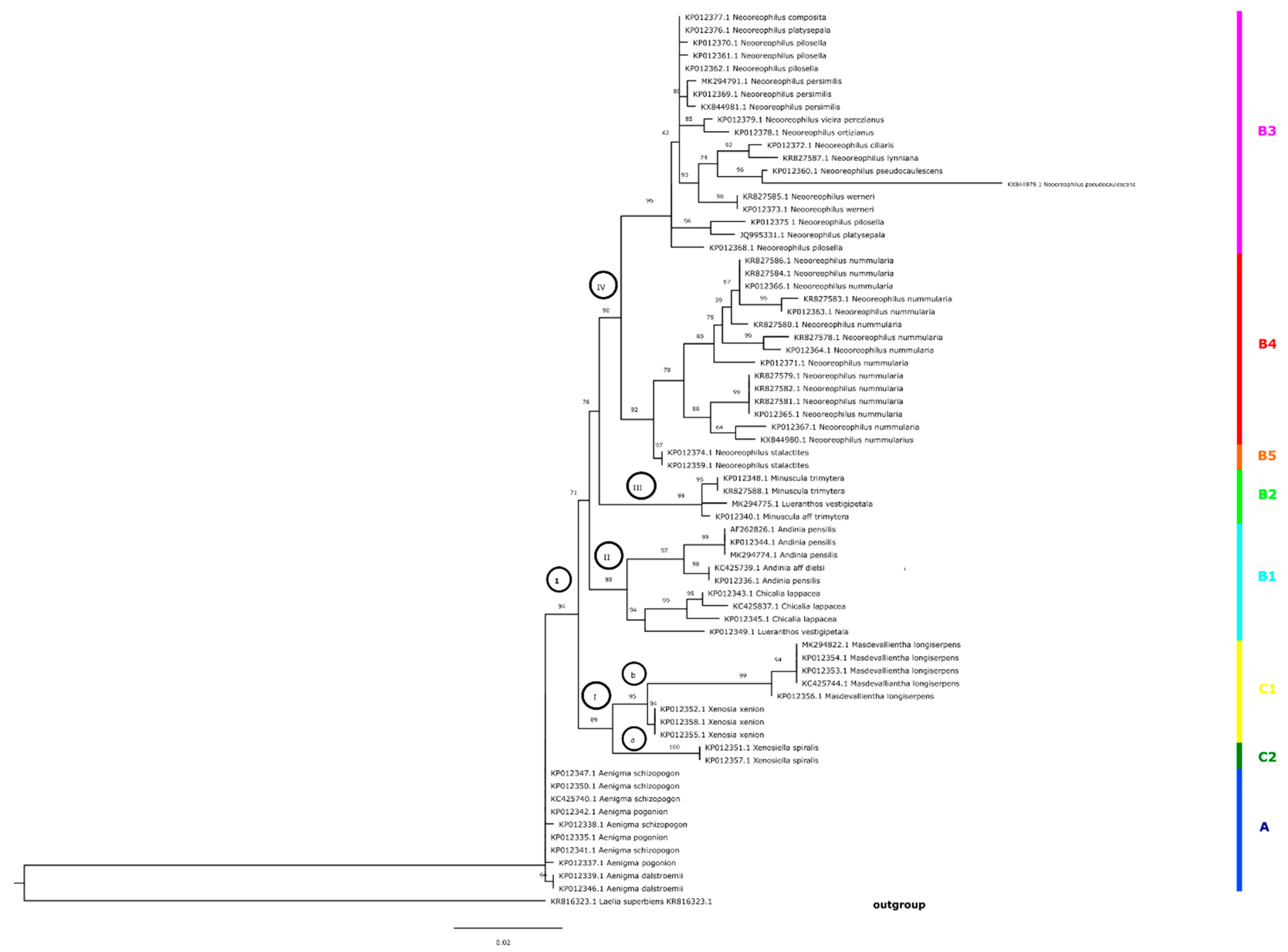{kind=link}
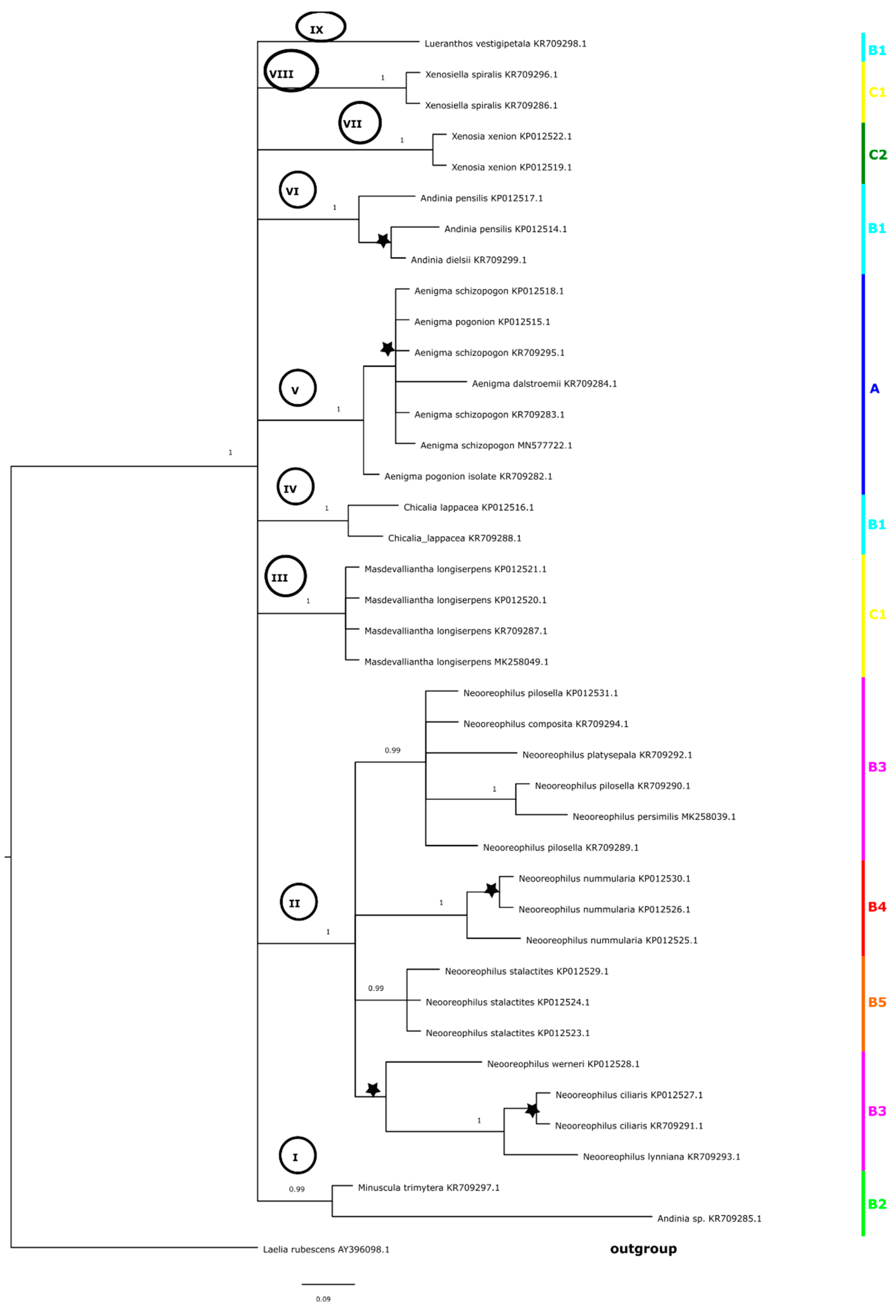{kind=link}
{kind=link}
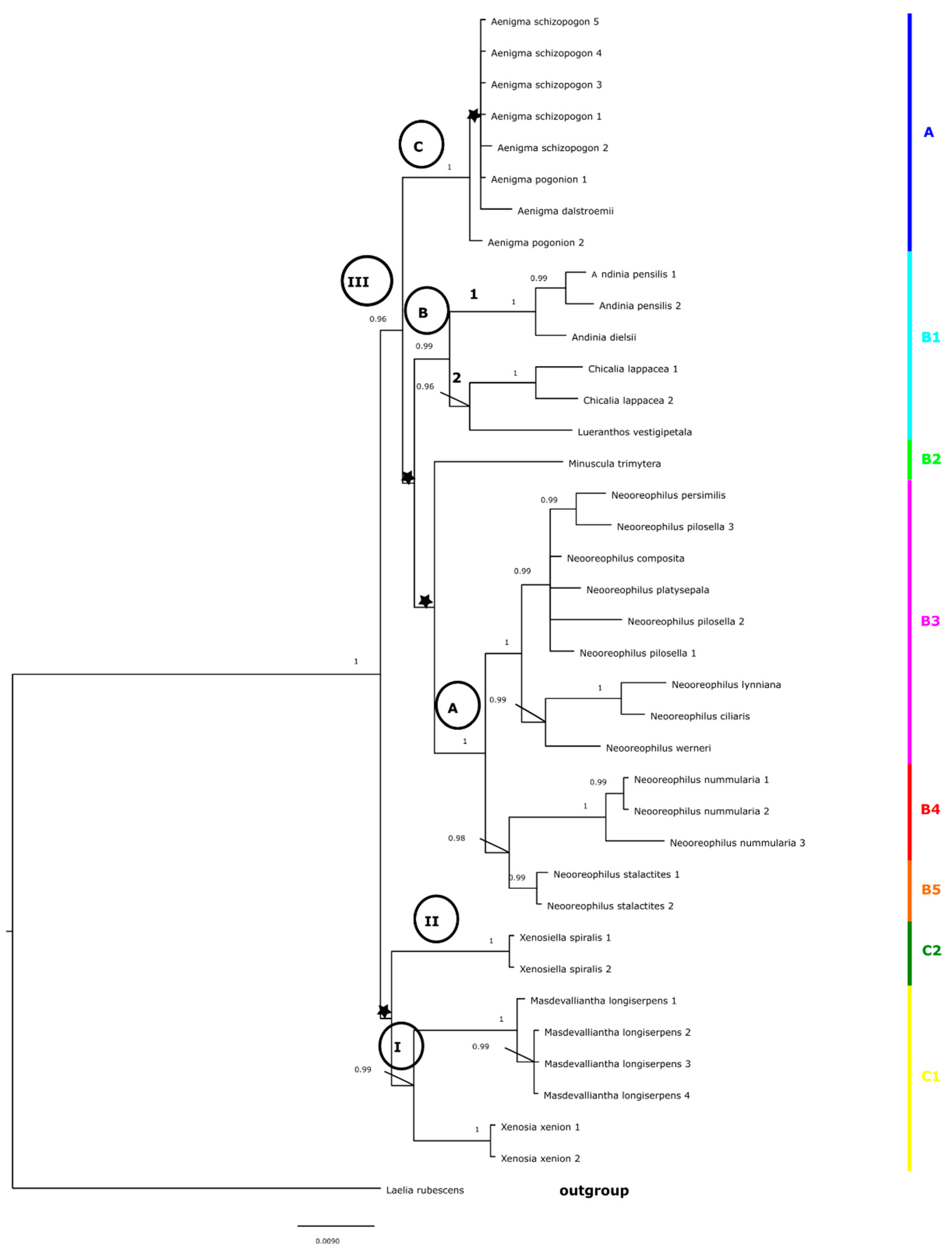{kind=link}
{kind=link}
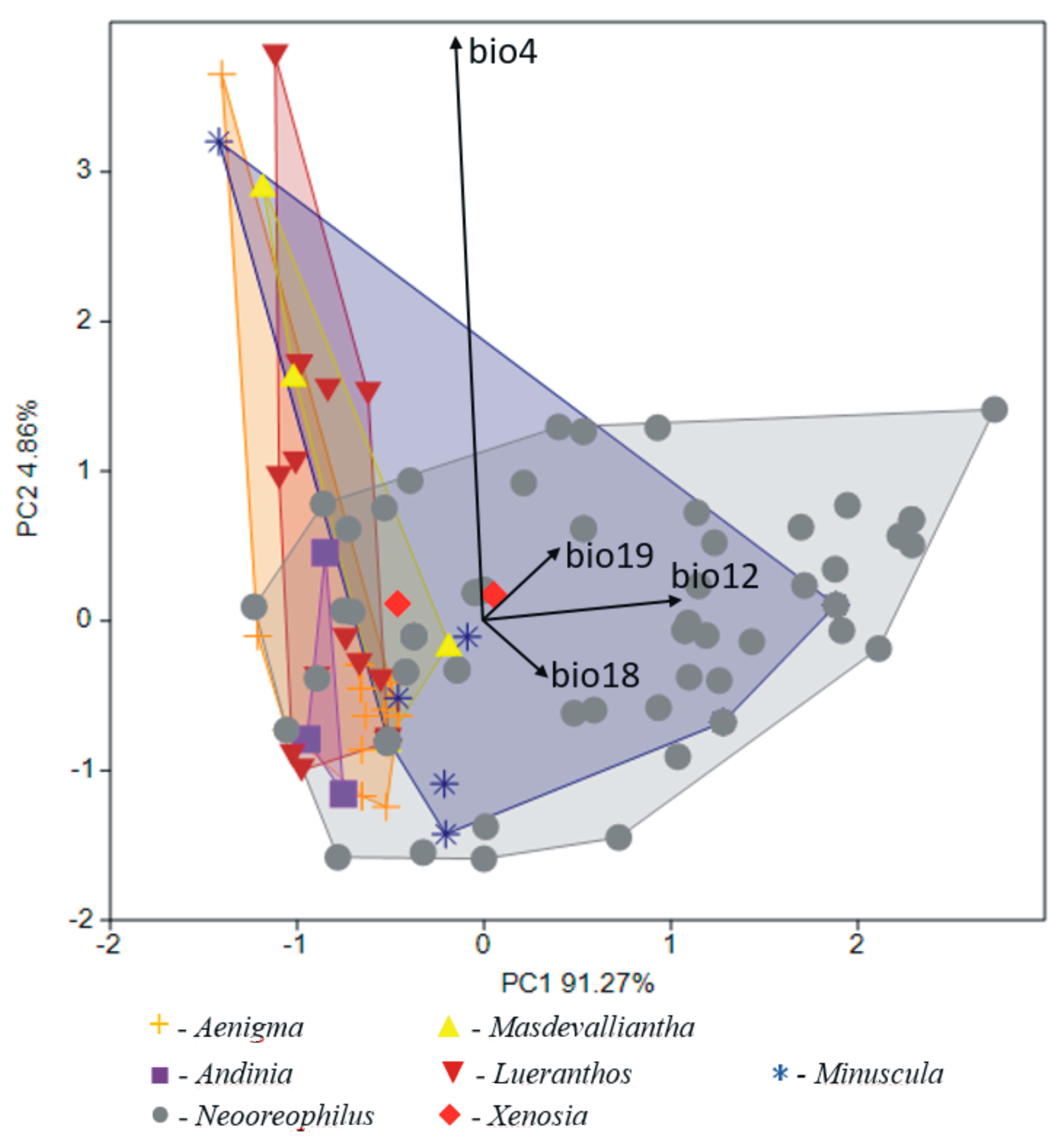{kind=link}
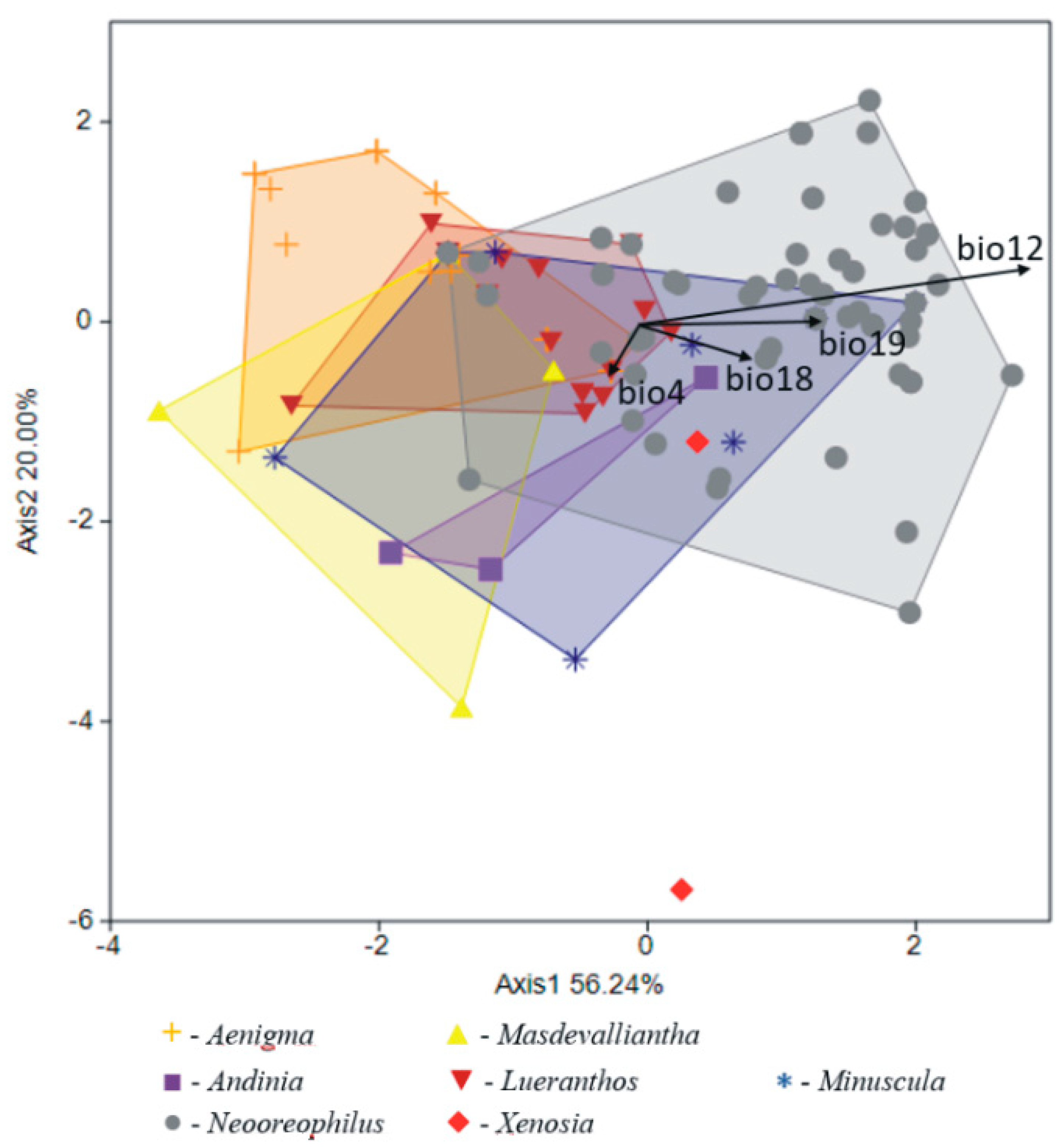{kind=link}
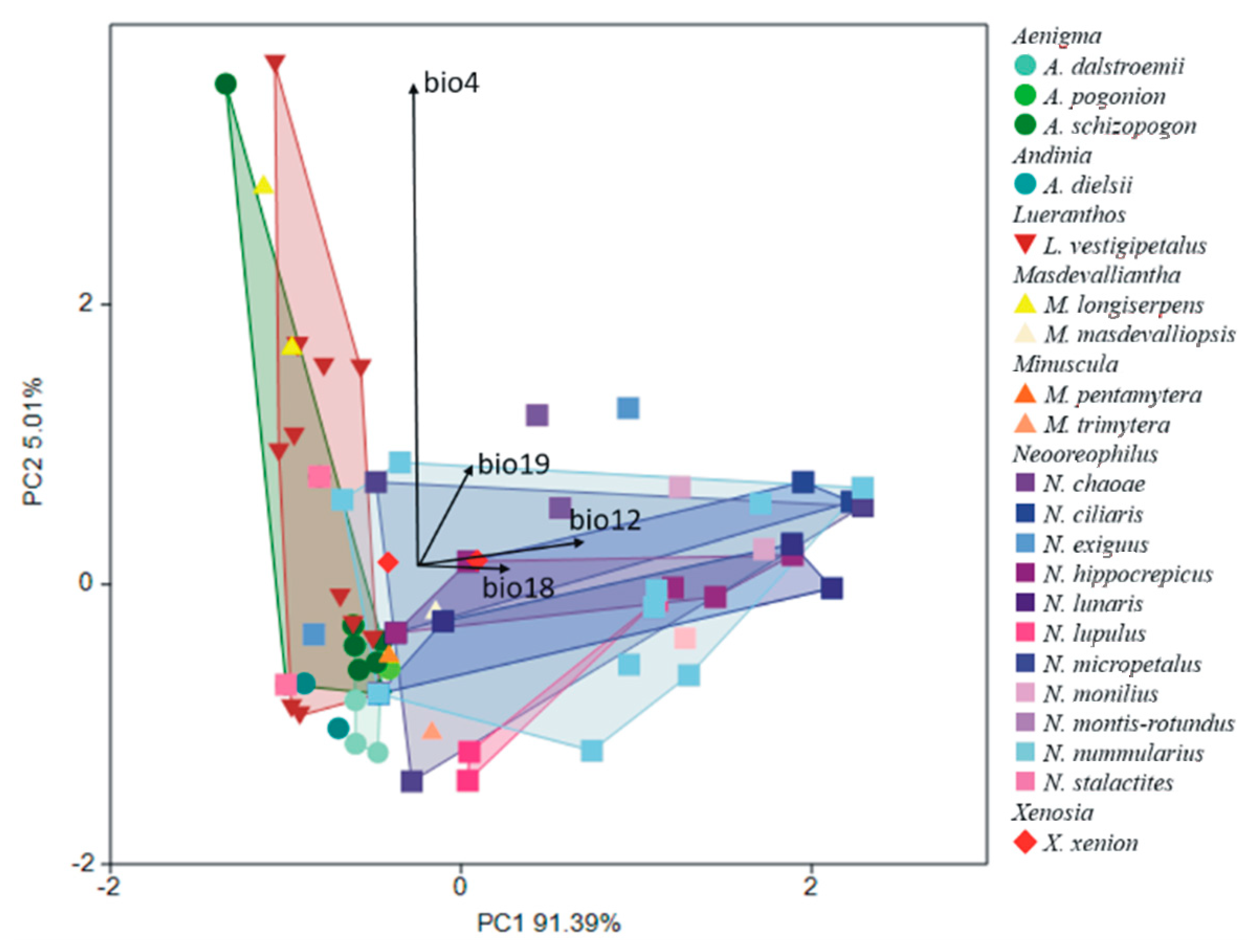{kind=link}
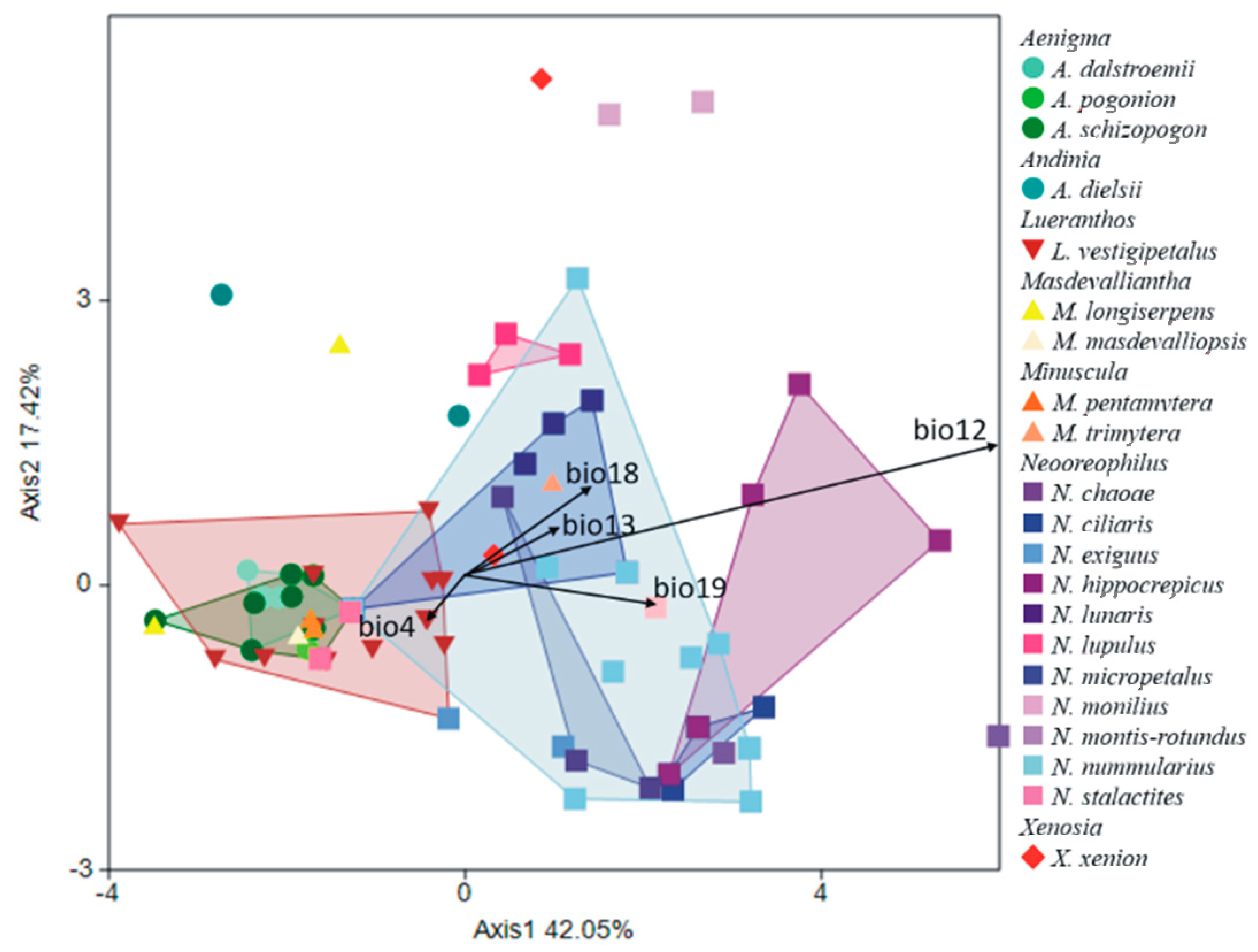{kind=link}
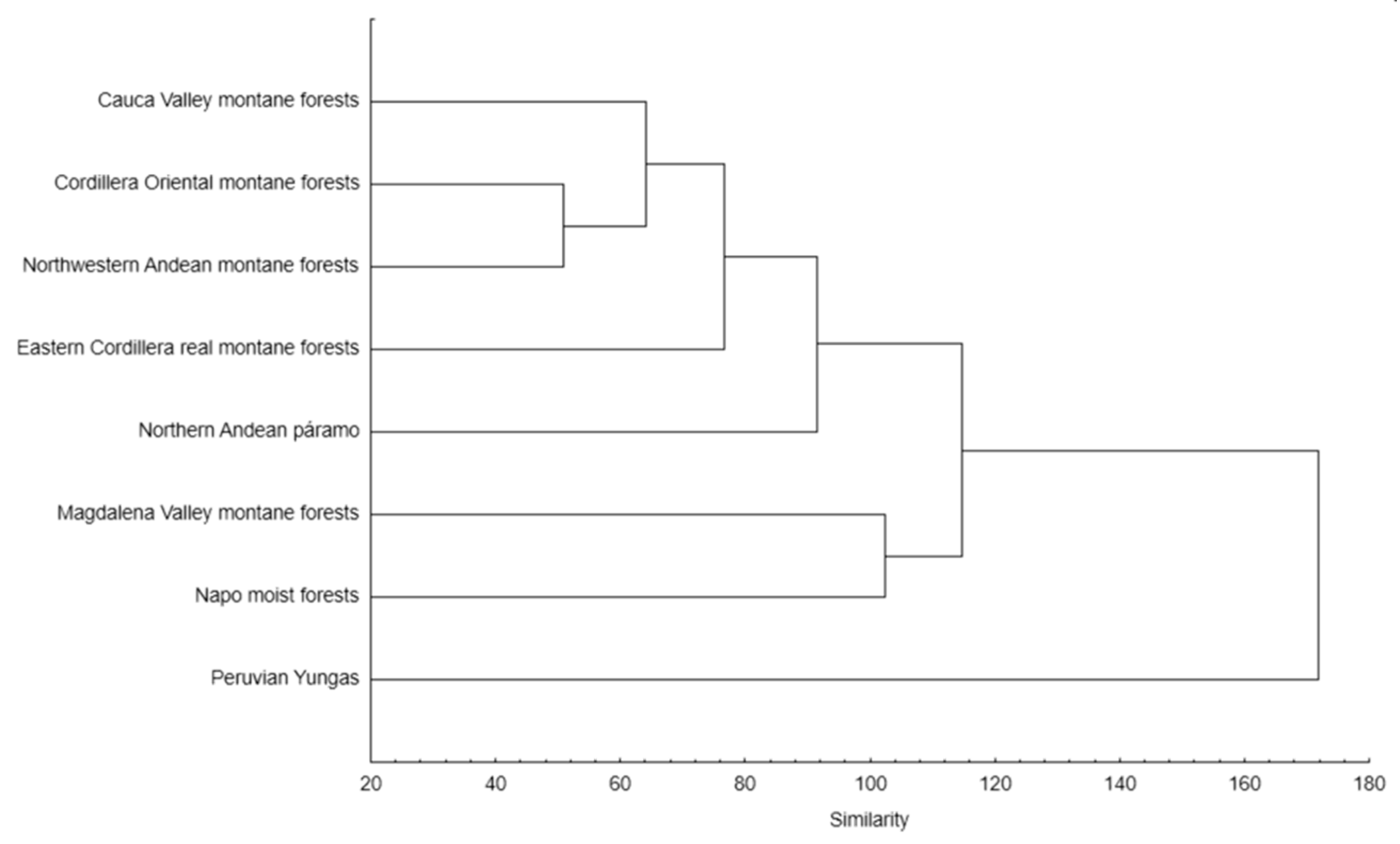{kind=link}
{kind=link}
{kind=link}
Table 1.
Codes of climatic variables used in this study.
| Code | Description |
|---|---|
| bio1 | Annual Mean Temperature |
| bio2 | Mean Diurnal Range = Mean of Monthly (max temp − min temp) |
| bio3 | Isothermality (bio2/bio7) × 100 |
| bio4 | Temperature Seasonality (standard deviation × 100) |
| bio5 | Max Temperature of Warmest Month |
| bio8 | Mean Temperature of Wettest Quarter |
| bio12 | Annual Precipitation |
| bio13 | Precipitation of Wettest Month |
| bio14 | Precipitation of Driest Month |
| bio15 | Precipitation Seasonality (Coefficient of Variation) |
| bio18 | Precipitation of Warmest Quarter |
| bio19 | Precipitation of Coldest Quarter |
Table 2.
Variables fitted to the PCA analysis focusing on climatic niche similarity of the studied species and determination coefficients (R2), where level of significance is marked (ns—not significant; *—p < 0.05; **—p < 0.01; ***—p < 0.001).
Table 2.
Variables fitted to the PCA analysis focusing on climatic niche similarity of the studied species and determination coefficients (R2), where level of significance is marked (ns—not significant; *—p < 0.05; **—p < 0.01; ***—p < 0.001).
| Variables | PC1 | PC2 | R2 |
|---|---|---|---|
| bio1 | 0.034 | −0.009 | 0.019 ns |
| bio2 | −0.009 | 0.034 | 0.019 *** |
| bio3 | 0.001 | −0.015 | 0.112 *** |
| bio4 | −0.061 | 0.992 | 0.046 ns |
| bio5 | 0.029 | 0.014 | 0.960 *** |
| bio8 | 0.032 | −0.005 | 0.996 *** |
| bio12 | 0.910 | 0.030 | 0.435 ns |
| bio13 | 0.104 | −0.017 | 0.451 ns |
| bio14 | 0.044 | 0.011 | 0.310 ns |
| bio15 | −0.066 | 0.011 | 0.026 ns |
| bio18 | 0.221 | −0.006 | 0.201 ns |
| bio19 | 0.322 | 0.112 | 0.412 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szlachetko, D.L.; Dudek, M.; Naczk, A.; Kolanowska, M. Taxonomy and Biogeography of Andinia-Complex (Orchidaceae). Diversity 2022, 14, 372. https://doi.org/10.3390/d14050372
AMA Style
Szlachetko DL, Dudek M, Naczk A, Kolanowska M. Taxonomy and Biogeography of Andinia-Complex (Orchidaceae). Diversity. 2022; 14(5):372. https://doi.org/10.3390/d14050372
Chicago/Turabian StyleSzlachetko, Dariusz L., Magdalena Dudek, Aleksandra Naczk, and Marta Kolanowska. 2022. "Taxonomy and Biogeography of Andinia-Complex (Orchidaceae)" Diversity 14, no. 5: 372. https://doi.org/10.3390/d14050372
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.