
Genome-Wide DNA Methylation in Policemen Working in Cities Differing by Major Sources of Air Pollution

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
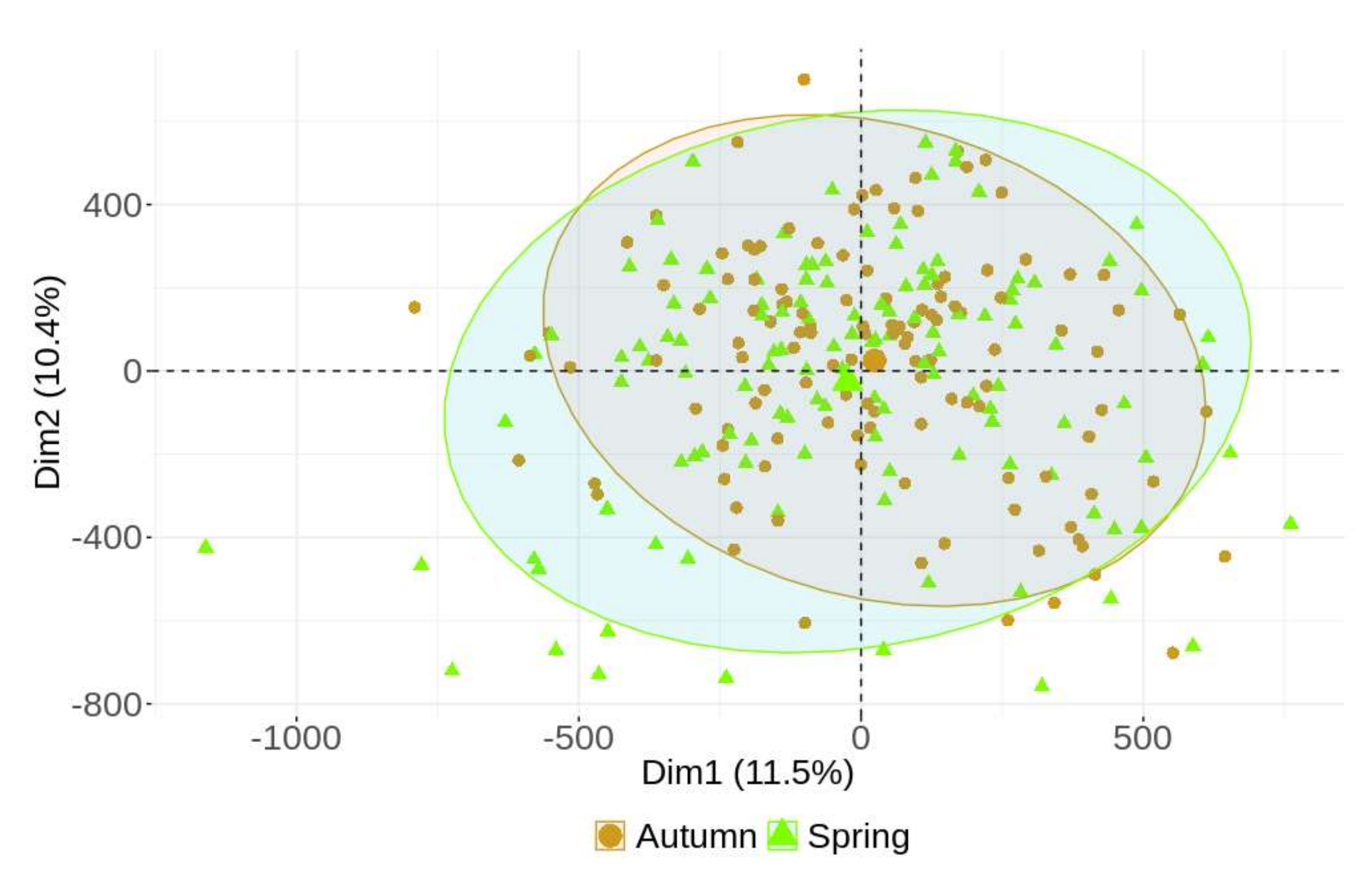
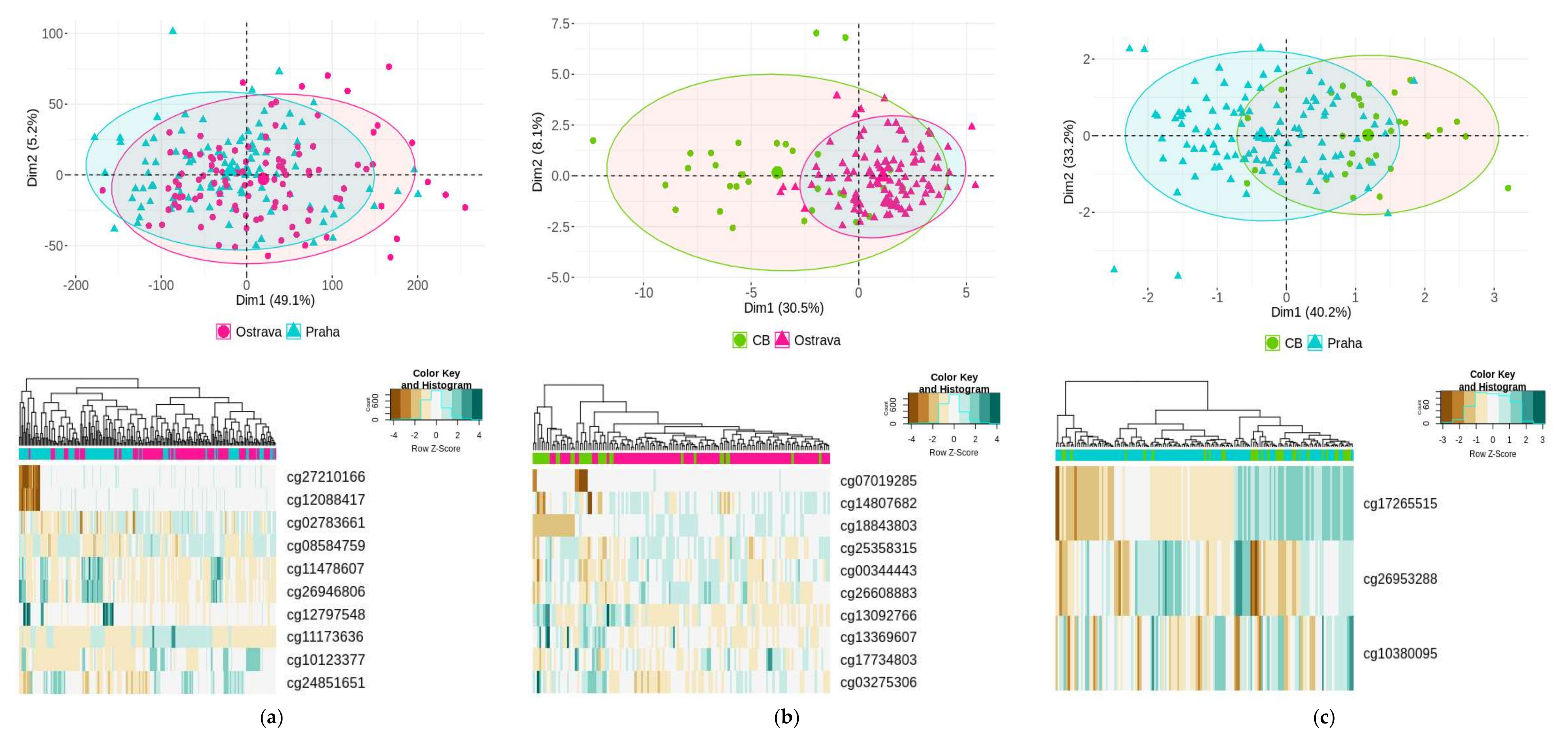
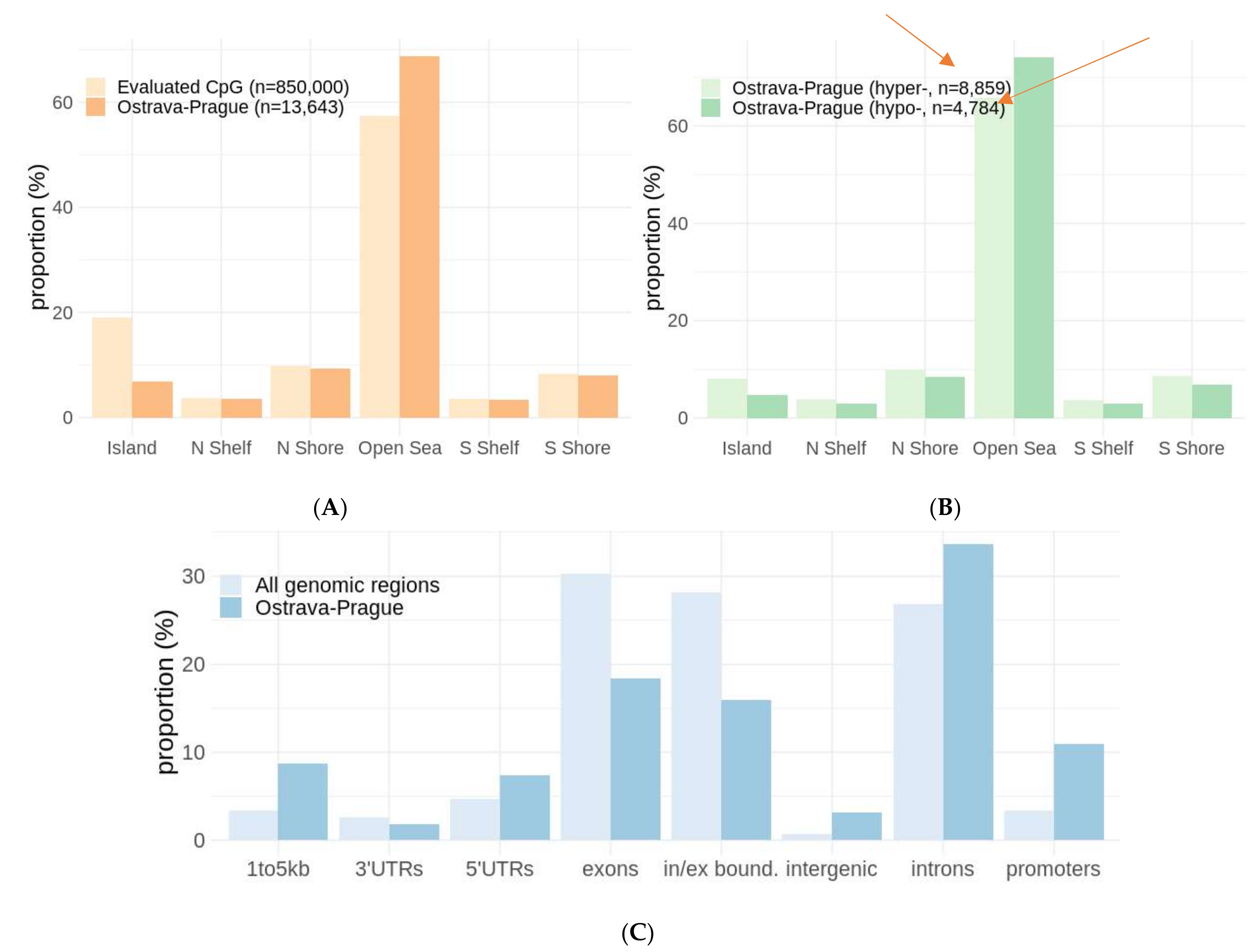
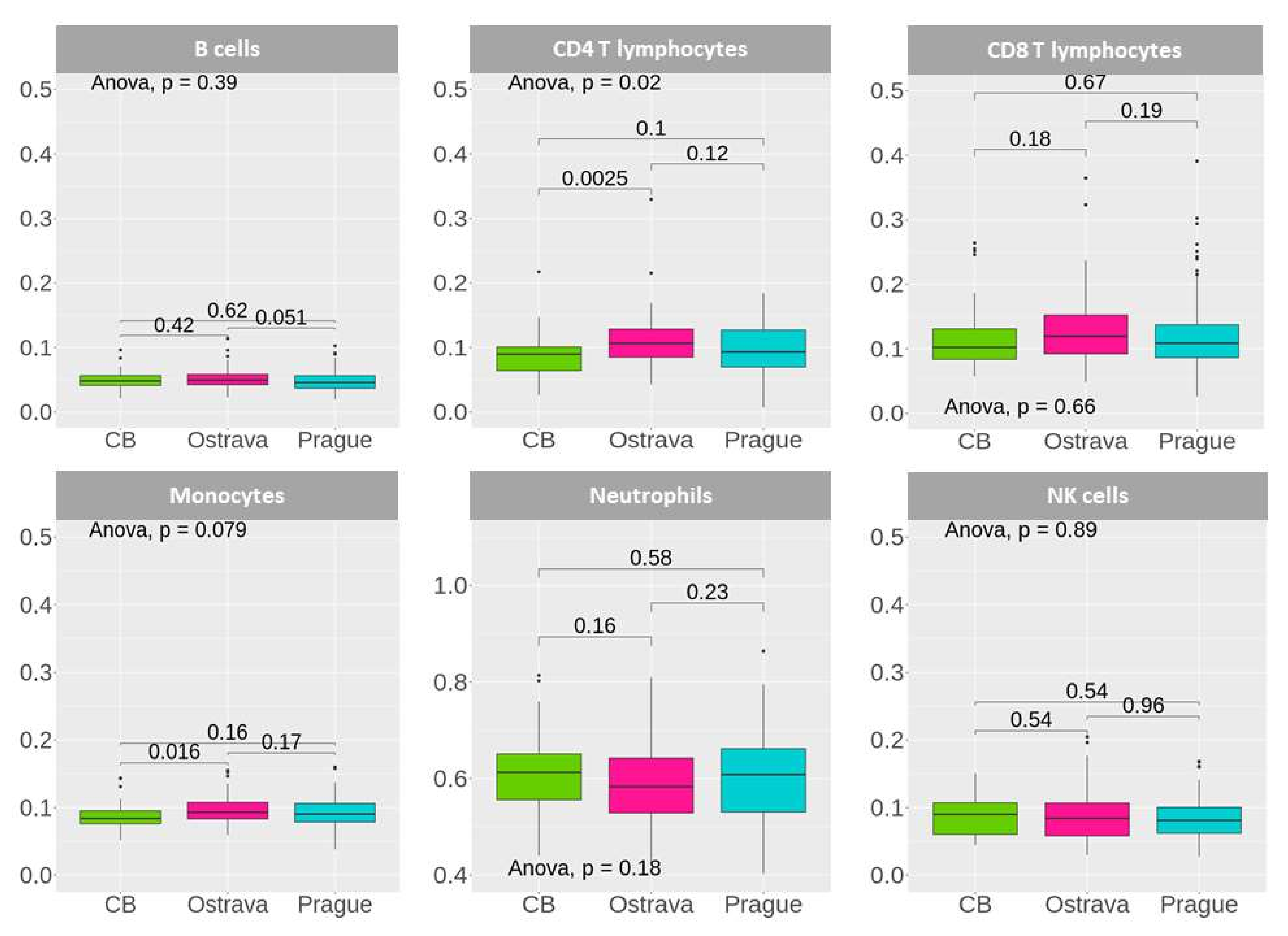
2. Results
2.1. Characteristics of Study Subjects
2.2. Air Pollution Monitoring
2.3. General DNA Methylation Profiling
2.4. Differentially Methylated Groups of Loci and Geneset Enrichment
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Air Pollution Monitoring
4.3. DNA Methylation Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3′UTR | 3′ Untranslated region |
| B[a]P | Benzo(a)pyrene |
| BCD | Bisulfite-converted DNA |
| BMI | Body mass index |
| CB | Ceske Budejovice |
| CpG | Cytosine-phosphate-Guanine dinucleotide |
| DML | Differentially methylated loci |
| DMG | Differentially methylated groups of loci |
| EDTA | Ethylenediaminetetraacetic acid |
| gDNA | Genomic DNA |
| EWAS | Epigenome-wide association study |
| FDR | False discovery rate |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| NHEJ | Non-homologous end joining |
| PCA | Principal component analysis |
| PM | Particulate matter |
| PM2.5 | Particulate matter less than 2.5 µm in diameter |
| SNPs | Single-nucleotide polymorphisms |
Appendix A

References
- Rider, C.F.; Carlsten, C. Air Pollution and DNA Methylation: Effects of Exposure in Humans. Clin. Epigen. 2019, 11, 131. [Google Scholar] [CrossRef] [PubMed]
- Curradi, M.; Izzo, A.; Badaracco, G.; Landsberger, N. Molecular Mechanisms of Gene Silencing Mediated by DNA Methylation. Mol. Cell. Biol. 2002, 22, 3157–3173. [Google Scholar] [CrossRef] [PubMed]
- Baccarelli, A.; Wright, R.O.; Bollati, V.; Tarantini, L.; Litonjua, A.A.; Suh, H.H.; Zanobetti, A.; Sparrow, D.; Vokonas, P.S.; Schwartz, J. Rapid DNA Methylation Changes after Exposure to Traffic Particles. Am. J. Respir. Crit. Care Med. 2009, 179, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, R.; Cai, J.; Cui, X.; Huang, N.; Kan, H. Short-Term Exposure to Fine Particulate Air Pollution and Genome-Wide DNA Methylation: A Randomized, Double-Blind, Crossover Trial. Environ. Int. 2018, 120, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Nwanaji-Enwerem, J.C.; Colicino, E. DNA Methylation-Based Biomarkers of Environmental Exposures for Human Population Studies. Curr. Environ. Health Rep. 2020, 7, 121–128. [Google Scholar] [CrossRef]
- Zeilinger, S.; Kühnel, B.; Klopp, N.; Baurecht, H.; Kleinschmidt, A.; Gieger, C.; Weidinger, S.; Lattka, E.; Adamski, J.; Peters, A.; et al. Tobacco Smoking Leads to Extensive Genome-Wide Changes in DNA Methylation. PLoS ONE 2013, 8, e63812. [Google Scholar] [CrossRef] [PubMed]
- Joubert, B.R.; Håberg, S.E.; Nilsen, R.M.; Wang, X.; Vollset, S.E.; Murphy, S.K.; Huang, Z.; Hoyo, C.; Midttun, Ø.; Cupul-Uicab, L.A.; et al. 450K Epigenome-Wide Scan Identifies Differential DNA Methylation in Newborns Related to Maternal Smoking during Pregnancy. Environ. Health Perspect. 2012, 120, 1425–1431. [Google Scholar] [CrossRef]
- Reese, S.E.; Zhao, S.; Wu, M.C.; Joubert, B.R.; Parr, C.L.; Håberg, S.E.; Ueland, P.M.; Nilsen, R.M.; Midttun, Ø.; Vollset, S.E.; et al. DNA Methylation Score as a Biomarker in Newborns for Sustained Maternal Smoking during Pregnancy. Environ. Health Perspect. 2017, 125, 760–766. [Google Scholar] [CrossRef]
- Liu, C.; Marioni, R.E.; Hedman, Å.K.; Pfeiffer, L.; Tsai, P.-C.; Reynolds, L.M.; Just, A.C.; Duan, Q.; Boer, C.G.; Tanaka, T.; et al. A DNA Methylation Biomarker of Alcohol Consumption. Mol. Psychiatry 2018, 23, 422–433. [Google Scholar] [CrossRef]
- Wright, R.O.; Schwartz, J.; Wright, R.J.; Bollati, V.; Tarantini, L.; Park, S.; Hu, H.; Sparrow, D.; Vokonas, P.; Baccarelli, A. Biomarkers of Lead Exposure and DNA Methylation within Retrotransposons. Environ. Health Perspect. 2010, 118, 790–795. [Google Scholar] [CrossRef]
- Boyne, D.J.; O’Sullivan, D.E.; Olij, B.F.; King, W.D.; Friedenreich, C.M.; Brenner, D.R. Physical Activity, Global DNA Methylation, and Breast Cancer Risk: A Systematic Literature Review and Meta-Analysis. Cancer Epidemiol. Biomark. Prev. 2018, 27, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Maghbooli, Z.; Hossein-Nezhad, A.; Adabi, E.; Asadollah-Pour, E.; Sadeghi, M.; Mohammad-Nabi, S.; Zakeri Rad, L.; Malek Hosseini, A.-A.; Radmehr, M.; Faghihi, F.; et al. Air Pollution during Pregnancy and Placental Adaptation in the Levels of Global DNA Methylation. PLoS ONE 2018, 13, e0199772. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.M. Epigenome-Wide Association Studies (EWAS): Past, Present, and Future. Methods Mol. Biol. 2015, 1238, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Gruzieva, O.; Xu, C.-J.; Breton, C.V.; Annesi-Maesano, I.; Antó, J.M.; Auffray, C.; Ballereau, S.; Bellander, T.; Bousquet, J.; Bustamante, M.; et al. Epigenome-Wide Meta-Analysis of Methylation in Children Related to Prenatal NO2 Air Pollution Exposure. Environ. Health Perspect. 2017, 125, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Gruzieva, O.; Xu, C.-J.; Yousefi, P.; Relton, C.; Merid, S.K.; Breton, C.V.; Gao, L.; Volk, H.E.; Feinberg, J.I.; Ladd-Acosta, C.; et al. Prenatal Particulate Air Pollution and DNA Methylation in Newborns: An Epigenome-Wide Meta-Analysis. Environ. Health Perspect. 2019, 127, 57012. [Google Scholar] [CrossRef]
- Jiang, C.-L.; He, S.-W.; Zhang, Y.-D.; Duan, H.-X.; Huang, T.; Huang, Y.-C.; Li, G.-F.; Wang, P.; Ma, L.-J.; Zhou, G.-B.; et al. Air Pollution and DNA Methylation Alterations in Lung Cancer: A Systematic and Comparative Study. Oncotarget 2016, 8, 1369–1391. [Google Scholar] [CrossRef]
- Sram, R.J.; Benes, I.; Binkova, B.; Dejmek, J.; Horstman, D.; Kotesovec, F.; Otto, D.; Perreault, S.D.; Rubes, J.; Selevan, S.G.; et al. Teplice Program—The Impact of Air Pollution on Human Health. Environ. Health Perspect. 1996, 104, 699–714. [Google Scholar]
- Jirik, V.; Machaczka, O.; Miturova, H.; Tomasek, I.; Slachtova, H.; Janoutova, J.; Velicka, H.; Janout, V. Air Pollution and Potential Health Risk in Ostrava Region—A Review. Cent. Eur. J. Public Health 2016, 24, S4–S17. [Google Scholar] [CrossRef]
- Sram, R.J.; Milcova, A.; Pastorkova, A.; Rossner, P.; Rossnerova, A.; Schmuczerova, J.; Spatova, M.; Svecova, V.; Topinka, J. European Hot Spot of Air Pollution by PM2.5 and Bap: Ostrava, Czech Republic. Epidemiology 2011, 22, S232. [Google Scholar] [CrossRef]
- Rossnerova, A.; Tulupova, E.; Tabashidze, N.; Schmuczerova, J.; Dostal, M.; Rossner, P.; Gmuender, H.; Sram, R.J. Factors Affecting the 27K DNA Methylation Pattern in Asthmatic and Healthy Children from Locations with Various Environments. Mutat. Res. 2013, 741–742, 18–26. [Google Scholar] [CrossRef]
- Honkova, K.; Rossnerova, A.; Pavlikova, J.; Svecova, V.; Klema, J.; Topinka, J.; Milcova, A.; Libalova, H.; Choi, H.; Veleminsky, M.; et al. Gene Expression Profiling in Healthy Newborns from Diverse Localities of the Czech Republic. Environ. Mol. Mutagenes. 2018, 59, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Rossner, P.; Tulupova, E.; Rossnerova, A.; Libalova, H.; Honkova, K.; Gmuender, H.; Pastorkova, A.; Svecova, V.; Topinka, J.; Sram, R.J. Reduced Gene Expression Levels after Chronic Exposure to High Concentrations of Air Pollutants. Mutat. Res. 2015, 780, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Rossner, P.; Uhlirova, K.; Beskid, O.; Rossnerova, A.; Svecova, V.; Sram, R.J. Expression of XRCC5 in Peripheral Blood Lymphocytes Is Upregulated in Subjects from a Heavily Polluted Region in the Czech Republic. Mutat. Res. 2011, 713, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Jirik, V.; Tomasek, L.; Fojtikova, I.; Janoš, T.; Stanovska, M.; Gunkova, P.; Dalecka, A.; Vrtkova, A.; Sram, R.J. Lifetime Carcinogenic Risk Proportions from Inhalation Exposures in Industrial and Non-Industrial Regions. Int. J. Environ. Res. Public Health 2021, 18, 13295. [Google Scholar] [CrossRef]
- Libalova, H.; Dostal, M.; Sram, R.J. Study of Gene Expression in Asthmatic Children Living in Localities with Differenct Extent of Air Pollution. Ochr. Ovzduší 2011, 23, 13–17. [Google Scholar]
- Kratenova, J.; Zejglicova, K.; Puklova, V. Respiratory Diseases in Children in the Moravian-Silesian Region in Comparison with the Results of National Monitoring. Hygiena 2018, 63, 116–121. [Google Scholar] [CrossRef]
- Zejglicova, K.; Puklova, V.; Kratenova, J.; Brabec, M.; Maly, M.; Tomek, M.; Urban, F.; Kubinova, R. Prevalence of Allergic Diseases and Respiratory Disorders in Children in Relation to Air Quality in the Moravian-Silesian Region. Hygiena 2019, 64, 45–51. [Google Scholar] [CrossRef]
- Stirzaker, C.; Taberlay, P.C.; Statham, A.L.; Clark, S.J. Mining Cancer Methylomes: Prospects and Challenges. Trends Genet. 2014, 30, 75–84. [Google Scholar] [CrossRef]
- Lanata, C.M.; Chung, S.A.; Criswell, L.A. DNA Methylation 101: What Is Important to Know about DNA Methylation and Its Role in SLE Risk and Disease Heterogeneity. Lupus Sci. Med. 2018, 5, e000285. [Google Scholar] [CrossRef]
- Rubes, J.; Sipek, J.; Kopecka, V.; Musilova, P.; Vozdova, M. Semen Quality and Sperm DNA Integrity in City Policemen Exposed to Polluted Air in an Urban Industrial Agglomeration. Int. J. Hyg. Environ. Health 2021, 237, 113835. [Google Scholar] [CrossRef]
- Ferrari, L.; Carugno, M.; Bollati, V. Particulate Matter Exposure Shapes DNA Methylation through the Lifespan. Clin. Epigenet. 2019, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Vojtisek-Lom, M.; Suta, M.; Sikorova, J.; Sram, R.J. High NO2 Concentrations Measured by Passive Samplers in Czech Cities: Unresolved Aftermath of Dieselgate? Atmosphere 2021, 12, 649. [Google Scholar] [CrossRef]
- Rossnerova, A.; Izzotti, A.; Pulliero, A.; Bast, A.; Rattan, S.I.S.; Rossner, P. The Molecular Mechanisms of Adaptive Response Related to Environmental Stress. Int. J. Mol. Sci. 2020, 21, 7053. [Google Scholar] [CrossRef] [PubMed]
- Rossnerova, A.; Honkova, K.; Chvojkova, I.; Pelclova, D.; Zdimal, V.; Hubacek, J.A.; Lischkova, L.; Vlckova, S.; Ondracek, J.; Dvorackova, S.; et al. Individual DNA Methylation Pattern Shifts in Nanoparticles-Exposed Workers Analyzed in Four Consecutive Years. Int. J. Mol. Sci. 2021, 22, 7834. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Lieber, M.R.; Yu, K.; Raghavan, S.C. Roles of Nonhomologous DNA End Joining, V(D)J Recombination, and Class Switch Recombination in Chromosomal Translocations. DNA Repair 2006, 5, 1234–1245. [Google Scholar] [CrossRef]
- Rossnerova, A.; Pokorna, M.; Svecova, V.; Sram, R.J.; Topinka, J.; Zölzer, F.; Rossner, P. Adaptation of the Human Population to the Environment: Current Knowledge, Clues from Czech Cytogenetic and “Omics” Biomonitoring Studies and Possible Mechanisms. Mutat. Res. Rev. Mutat. Res. 2017, 773, 188–203. [Google Scholar] [CrossRef]
- Miller, S.; Tsou, P.-S.; Coit, P.; Gensterblum-Miller, E.; Renauer, P.; Rohraff, D.M.; Kilian, N.C.; Schonfeld, M.; Sawalha, A.H. Hypomethylation of STAT1 and HLA-DRB1 Is Associated with Type-I Interferon-Dependent HLA-DRB1 Expression in Lupus CD8+ T Cells. Ann. Rheum. Dis. 2019, 78, 519–528. [Google Scholar] [CrossRef]
- Rebbeck, T.R. Molecular Epidemiology of the Human Glutathione S-Transferase Genotypes GSTM1 and GSTT1 in Cancer Susceptibility. Cancer Epidemiol. Biomark. Prev. 1997, 6, 733–743. [Google Scholar]
- Naccarati, A.; Soucek, P.; Stetina, R.; Haufroid, V.; Kumar, R.; Vodickova, L.; Trtkova, K.; Dusinska, M.; Hemminki, K.; Vodicka, P. Genetic Polymorphisms and Possible Gene-Gene Interactions in Metabolic and DNA Repair Genes: Effects on DNA Damage. Mutat. Res. 2006, 593, 22–31. [Google Scholar] [CrossRef]
- Levinsson, A.; Olin, A.-C.; Modig, L.; Dahgam, S.; Björck, L.; Rosengren, A.; Nyberg, F. Interaction Effects of Long-Term Air Pollution Exposure and Variants in the GSTP1, GSTT1 and GSTCD Genes on Risk of Acute Myocardial Infarction and Hypertension: A Case-Control Study. PLoS ONE 2014, 9, e99043. [Google Scholar] [CrossRef]
- Caubit, X.; Thoby-Brisson, M.; Voituron, N.; Filippi, P.; Bévengut, M.; Faralli, H.; Zanella, S.; Fortin, G.; Hilaire, G.; Fasano, L. Teashirt 3 Regulates Development of Neurons Involved in Both Respiratory Rhythm and Airflow Control. J. Neurosci. 2010, 30, 9465–9476. [Google Scholar] [CrossRef]
- Kajiwara, Y.; Akram, A.; Katsel, P.; Haroutunian, V.; Schmeidler, J.; Beecham, G.; Haines, J.L.; Pericak-Vance, M.A.; Buxbaum, J.D. FE65 Binds Teashirt, Inhibiting Expression of the Primate-Specific Caspase-4. PLoS ONE 2009, 4, e5071. [Google Scholar] [CrossRef] [PubMed]
- Guertin, D.A.; Sabatini, D.M. Defining the Role of MTOR in Cancer. Cancer Cell 2007, 12, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Weichhart, T.; Säemann, M.D. The Multiple Facets of MTOR in Immunity. Trends Immunol. 2009, 30, 218–226. [Google Scholar] [CrossRef]
- Kim, D.-H.; Sarbassov, D.D.; Ali, S.M.; King, J.E.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. MTOR Interacts with Raptor to Form a Nutrient-Sensitive Complex That Signals to the Cell Growth Machinery. Cell 2002, 110, 163–175. [Google Scholar] [CrossRef]
- Blandino-Rosano, M.; Bernal-Mizrachi, E. 2124-P: Raptor Levels Are Critical in the Adaptation of Beta Cells to High-Fat Diet. Diabetes 2020, 69, 2124. [Google Scholar] [CrossRef]
- Ni, Q.; Gu, Y.; Xie, Y.; Yin, Q.; Zhang, H.; Nie, A.; Li, W.; Wang, Y.; Ning, G.; Wang, W.; et al. Raptor Regulates Functional Maturation of Murine Beta Cells. Nat. Commun. 2017, 8, 15755. [Google Scholar] [CrossRef]
- Banyard, J.; Bao, L.; Hofer, M.D.; Zurakowski, D.; Spivey, K.A.; Feldman, A.S.; Hutchinson, L.M.; Kuefer, R.; Rubin, M.A.; Zetter, B.R. Collagen XXIII Expression Is Associated with Prostate Cancer Recurrence and Distant Metastases. Clin. Cancer Res. 2007, 13, 2634–2642. [Google Scholar] [CrossRef]
- Spivey, K.A.; Banyard, J.; Solis, L.M.; Wistuba, I.I.; Barletta, J.A.; Gandhi, L.; Feldman, H.A.; Rodig, S.J.; Chirieac, L.R.; Zetter, B.R. Collagen XXIII: A Potential Biomarker for the Detection of Primary and Recurrent Non-Small Cell Lung Cancer. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1362–1372. [Google Scholar] [CrossRef]
- Abbott, G.W. Biology of the KCNQ1 Potassium Channel. New J. Sci. 2014, 2014, e237431. [Google Scholar] [CrossRef]
- Rajender, S.; Avery, K.; Agarwal, A. Epigenetics, Spermatogenesis and Male Infertility. Mutat. Res. 2011, 727, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Travers, M.E.; Mackay, D.J.G.; Nitert, M.D.; Morris, A.P.; Lindgren, C.M.; Berry, A.; Johnson, P.R.; Hanley, N.; Groop, L.C.; McCarthy, M.I.; et al. Insights into the Molecular Mechanism for Type 2 Diabetes Susceptibility at the KCNQ1 Locus From Temporal Changes in Imprinting Status in Human Islets. Diabetes 2013, 62, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Li, Q.; Xie, H.; Chen, Z.J.; Borovitskaya, A.E.; Maclaren, N.K.; Notkins, A.L.; Lan, M.S. Identification of a Second Transmembrane Protein Tyrosine Phosphatase, IA-2beta, as an Autoantigen in Insulin-Dependent Diabetes Mellitus: Precursor of the 37-KDa Tryptic Fragment. Proc. Natl. Acad. Sci. USA 1996, 93, 2307–2311. [Google Scholar] [CrossRef]
- Den Dekker, H.T.; Burrows, K.; Felix, J.F.; Salas, L.A.; Nedeljkovic, I.; Yao, J.; Rifas-Shiman, S.L.; Ruiz-Arenas, C.; Amin, N.; Bustamante, M.; et al. Newborn DNA-Methylation, Childhood Lung Function, and the Risks of Asthma and COPD across the Life Course. Eur. Respir. J. 2019, 53. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, S.L.; Eliot, M.N.; Whitsel, E.A.; Huang, Y.-T.; Kelsey, K.T.; Marsit, C.J.; Wellenius, G.A. Maternal Residential Proximity to Major Roadways, Birth Weight, and Placental DNA Methylation. Environ. Int. 2016, 92, 43–49. [Google Scholar] [CrossRef]
- Sacchetti, P.; Carpentier, R.; Ségard, P.; Olivé-Cren, C.; Lefebvre, P. Multiple Signaling Pathways Regulate the Transcriptional Activity of the Orphan Nuclear Receptor NURR1. Nucleic Acids Res. 2006, 34, 5515–5527. [Google Scholar] [CrossRef]
- Doi, Y.; Oki, S.; Ozawa, T.; Hohjoh, H.; Miyake, S.; Yamamura, T. Orphan Nuclear Receptor NR4A2 Expressed in T Cells from Multiple Sclerosis Mediates Production of Inflammatory Cytokines. Proc. Natl. Acad. Sci. USA 2008, 105, 8381–8386. [Google Scholar] [CrossRef]
- Ruiz-Sánchez, E.; Yescas, P.; Rodríguez-Violante, M.; Martínez-Rodríguez, N.; Díaz-López, J.N.; Ochoa, A.; Valdes-Rojas, S.S.; Magos-Rodríguez, D.; Rojas-Castañeda, J.C.; Cervantes-Arriaga, A.; et al. Association of Polymorphisms and Reduced Expression Levels of the NR4A2 Gene with Parkinson’s Disease in a Mexican Population. J. Neurol. Sci. 2017, 379, 58–63. [Google Scholar] [CrossRef]
- Montarolo, F.; Perga, S.; Martire, S.; Navone, D.N.; Marchet, A.; Leotta, D.; Bertolotto, A. Altered NR4A Subfamily Gene Expression Level in Peripheral Blood of Parkinson’s and Alzheimer’s Disease Patients. Neurotox. Res. 2016, 30, 338–344. [Google Scholar] [CrossRef]
- Le, W.; Pan, T.; Huang, M.; Xu, P.; Xie, W.; Zhu, W.; Zhang, X.; Deng, H.; Jankovic, J. Decreased NURR1 Gene Expression in Patients with Parkinson’s Disease. J. Neurol. Sci. 2008, 273, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wei, L.; Tao, Q.; Deng, H.; Ming, M.; Xu, P.; Le, W. Decreased NURR1 and PITX3 Gene Expression in Chinese Patients with Parkinson’s Disease. Eur. J. Neurol. 2012, 19, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Zolochevska, O.; Figueiredo, M.L. Novel Tumor Growth Inhibition Mechanism by Cell Cycle Regulator Cdk2ap1 Involves Antiangiogenesis Modulation. Microvasc. Res. 2010, 80, 324–331. [Google Scholar] [CrossRef]
- Battum, E.Y.V.; Brignani, S.; Pasterkamp, R.J. Axon Guidance Proteins in Neurological Disorders. Lancet Neurol. 2015, 14, 532–546. [Google Scholar] [CrossRef]
- Antonell, A.; Lladó, A.; Altirriba, J.; Botta-Orfila, T.; Balasa, M.; Fernández, M.; Ferrer, I.; Sánchez-Valle, R.; Molinuevo, J.L. A Preliminary Study of the Whole-Genome Expression Profile of Sporadic and Monogenic Early-Onset Alzheimer’s Disease. Neurobiol. Aging 2013, 34, 1772–1778. [Google Scholar] [CrossRef]
- Ambroz, A.; Vlkova, V.; Rossner, P.; Rossnerova, A.; Svecova, V.; Milcova, A.; Pulkrabova, J.; Hajslova, J.; Veleminsky, M.; Solansky, I.; et al. Impact of Air Pollution on Oxidative DNA Damage and Lipid Peroxidation in Mothers and Their Newborns. Int. J. Hyg. Environ. Health 2016, 219, 545–556. [Google Scholar] [CrossRef]
- Sram, R.J.; Binkova, B.; Dostal, M.; Merkerova-Dostalova, M.; Libalova, H.; Milcova, A.; Rossner, P.; Rossnerova, A.; Schmuczerova, J.; Svecova, V.; et al. Health Impact of Air Pollution to Children. Int. J. Hyg. Environ. Health 2013, 216, 533–540. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A Simple Salting out Procedure for Extracting DNA from Human Nucleated Cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A Flexible and Comprehensive Bioconductor Package for the Analysis of Infinium DNA Methylation Microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef]
- McCartney, D.L.; Walker, R.M.; Morris, S.W.; McIntosh, A.M.; Porteous, D.J.; Evans, K.L. Identification of Polymorphic and Off-Target Probe Binding Sites on the Illumina Infinium MethylationEPIC BeadChip. Genom. Data 2016, 9, 22–24. [Google Scholar] [CrossRef]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Jaffe, A.E.; Storey, J.D. The Sva Package for Removing Batch Effects and Other Unwanted Variation in High-Throughput Experiments. Bioinformatics 2012, 28, 882–883. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xie, Z.; Le Pape, M.; Dye, T. An Evaluation of Statistical Methods for DNA Methylation Microarray Data Analysis. BMC Bioinform. 2015, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, R.G.; Sartor, M.A. Annotatr: Genomic Regions in Context. Bioinformatics 2017, 33, 2381–2383. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA Methylation Arrays as Surrogate Measures of Cell Mixture Distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Locality | Ostrava | Prague | CB |
|---|---|---|---|
| Number of samples/subjects | 108/54 | 110/55 | 32/16 |
| Age (years) | 40.4; 9.3 (42.0; 21–61) | 39.3; 9.2 (38.0; 23–63) | 38.0; 6.4 (38.0; 22–48) |
| Mean; SD (median; min–max) | |||
| BMI | 28.6; 4.1 (28.3; 20.4–44.8) | 28.4; 3.9 (28.7; 19.4–36.8) | 28.2; 3.7 (27.3; 23.2–41.0) |
| Mean; SD (median; min–max) | |||
| Occupation duration (years) | 13.9; 7.9 (14.0; 0.8–26.8) | 11.8; 6.7 (12.0; 1.1–25.9) | 10.2; 7.5 (10.0; 0.8–26.5) |
| Mean; SD (median; min–max) |
| (A) | |||||||
|---|---|---|---|---|---|---|---|
| Ostrava | Prague | CB | |||||
| Winter | Summer | Winter | Summer | Winter | Summer | ||
| PM2.5 | Mean; SD | 24.4; 19.1 | 11.3; 5.3 | 15.0; 13.4 | 9.1; 4.2 | 15.4; 11.9 | 9.7; 4.2 |
| µg/m3 | Median (min–max) | 19.8 (2.6–101.8) | 10.3 (3.6–40.5) | 10.4 (1.5–57.4) | 8.3 (2.0–22.2) | 12.3 (1.5–47.6) | 9.1 (3.3–22.2) |
| B[a]P | Mean; SD | 4.1; 3.3 | 0.3; 0.3 | 3.6; 3.0 | 0.1; 0.1 | 2.00; 1.01 | 0.1; 0.1 |
| ng/m3 | Median (min–max) | 2.6 (0.3–13.5) | 0.1 (0.0–1.3) | 1.6 (0.0–16.7) | 0.0 (0.0–0.5) | 1.7 (0.60–5.10) | 0.0 (0.0–0.9) |
| NO2 | Mean; SD | 19.9; 9.4 | 10.9; 3.70 | 19.5; 8.9 | 17.9; 5.5 | 17.6; 8.2 | 9.8; 2.6 |
| µg/m3 | Median (min–max) | 19.0 (4.5–51.5) | 10.3 (4.4–23.2) | 16.9 (4.7–47.9) | 18.3 (8.3–32.9) | 14.1 (4.8–44.4) | 9.8 (3.4–18.3) |
| (B) | |||||||
| PM2.5 (µg/m3) | 2016 | 2017 | 2018 | 2019 | |||
| Ostrava | Mean; SD | 22.2; 18.3 | 21.7; 15.9 | 22.9; 18.2 | 17.4; 14.4 | ||
| Prague | 16.5; 13.8 | 16.7; 12.2. | 18.0; 15.2 | 12.3; 9.9 | |||
| CB | 18.5; 14.8 | 14.6; 10.4 | 16.0; 13.6 | 12.8; 10.4 | |||
| B[a]P (ng/m3) | 2016 | 2017 | 2018 | 2019 | |||
| Ostrava | Mean; SD | 2.2; 0.9 | 2.5; 0.7 | 2.9; 0.9 | 2.0; 0.6 | ||
| Prague * | 0.8; 0.3 | 0.9; 0.3 | 0.8; 0.2 | 0.7; 0.2 | |||
| CB | 1.5; 0.5 | 1.3; 0.5 | 1.1; 0.3 | 1.2; 0.4 | |||
| NO2 (µg/m3) | 2016 | 2017 | 2018 | 2019 | |||
| Ostrava | Mean; SD | 16.4; 15.0 | 16.2; 13.6 | 17.2; 15.4 | 15.2; 13.5 | ||
| Prague # | 25.6; 24.0 | 31.0; 29.1 | 33.0; 31.8 | 28.6; 27.5 | |||
| CB | 15.7; 14.2 | 15.4; 13.7 | 14.9; 13.8 | 12.9; 6.9 | |||
| CpG Locus | Chromosome | Relation to Island * | Gene | log2FC | Adj p-Value | |
|---|---|---|---|---|---|---|
| (a) Ostrava–Prague | cg10123377 | 3 | Open Sea | -- | −1.92 | 8.30 × 10−4 |
| cg24851651 | 11 | S Shelf | CCS | −1.21 | 8.53 × 10−3 | |
| cg11173636 | 10 | Open Sea | RP11-170M17.1 | −1.19 | 4.04 × 10−3 | |
| cg27210166 | 17 | Open Sea | RPTOR | −0.90 | 2.55 × 10−3 | |
| cg08584759 | 10 | Island | C10orf47 | −0.87 | 8.30 × 10−4 | |
| cg12088417 | 17 | Open Sea | RPTOR | −0.86 | 7.05 × 10−4 | |
| cg02783661 | 12 | S Shelf | CCDC77 | −0.78 | 2.91 × 10−3 | |
| cg11478607 | 22 | Island | GSTT1 | 0.76 | 8.62 × 10−3 | |
| cg26946806 | 22 | S Shore | GSTT1 | 0.81 | 8.37 × 10−3 | |
| cg12797548 | 1 | Open Sea | NME7 | 0.83 | 6.69 × 10−4 | |
| (b) Ostrava–CB | cg13092766 | 2 | Island | BCL2L11 | −0.91 | 7.49 × 10−3 |
| cg13369607 | 19 | Island | SAFB2 | −0.63 | 5.44 × 10−3 | |
| cg03275306 | 15 | Open Sea | BCL2A1 | −0.53 | 3.63 × 10−3 | |
| cg00344443 | 11 | Island | AP000797.3 | 0.53 | 7.49 × 10−3 | |
| cg25358315 | 2 | Island | CAPG | 0.57 | 3.63 × 10−3 | |
| cg14807682 | 8 | Island | -- | 0.57 | 7.49 × 10−3 | |
| cg17734803 | 1 | Island | FOXD3-AS1 | 0.62 | 7.49 × 10−3 | |
| cg26608883 | 11 | Island | CALCB | 0.71 | 7.49 × 10−3 | |
| cg07019285 | 10 | Open Sea | CALML3-AS1 | 0.87 | 9.86 × 10−3 | |
| cg18843803 | 19 | Open Sea | TSHZ3 | 1.95 | 3.63 × 10−3 | |
| (c) Prague–CB | cg17265515 | 8 | Open Sea | ERICH1 | −1.59 | 5.05 × 10−3 |
| cg10380095 | 7 | Open Sea | -- | −0.31 | 5.05 × 10−3 | |
| cg26953288 | 8 | S Shore | BAI1 | 0.55 | 1.48 × 10−3 |
| (A) | |||
|---|---|---|---|
| Gene | CpG Probes | Adj. p-Value (Min–Max) | Importance |
| XRCC5 (N = 2: chr 2) | Hypo: cg23433242, cg01633232 | 5.62 × 10−3, 2.79 × 10−3 | DNA repair–non homologous end joining (NHEJ) |
| NR4A2 (N = 9: chr 2) | Hyper: cg14617996, cg11209121, cg11358945, cg00194126, cg03339537, cg06101180, cg14811105, cg20570611, cg13500877 | 4.61 × 10−3 – 9.60 × 10−3 | Autoimmune diseases, neurodegenerative diseases |
| CDK2AP1 (N = 9: chr 12) | Hyper: cg10289269, cg05760918, cg01660796, cg01247747, cg02017926, cg25326086, cg08696931, cg25630910, cg10411075 | 7.40 × 10−4 – 6.28 × 10−3 | Reduction in cell proliferation, Cell cycle termination |
| (B) | |||
| Gene | CpG Probes | Adj. p-Value (Min–Max) | Importance |
| RPTOR (N = 20: chr 17) | Hypo: cg12088417, cg17614805, cg27210166, cg00545580 Hyper: cg12654199, cg22386583, cg13549638, cg07856428, cg22091236, cg15230985, cg27457201, cg02675920, cg22652378, cg08129331, cg13159505, cg22255288, cg15781195, cg14343513, cg07702089, cg05098037 | 7.05 × 10−4 – 9.15 × 10−3 | RPTOR encodes part of a signaling pathway regulating cell growth which responds to nutrient and insulin levels. |
| COL23A1 (N = 17: chr 5) | Hypo: cg06485308, cg02119888 Hyper: cg08862479, cg17649857, cg24705158, cg06882830, cg04686319, cg18816996, cg21871330, cg21768539, cg25153741, cg24830898, cg05206633, cg08684511, cg26994283, cg24527008, cg00561180 | 1.16 × 10−3 – 9.08 × 10−3 | COL23A1 encodes a transmembrane nonfibrillar collagen. This kind of collagen has a single pass hydrophobic transmembrane domain. |
| KCNQ1 (N = 16: chr 11) | Hypo: cg21752270, cg00243281, cg24492694, cg06827779 Hyper: cg23762359, cg05472575, cg12141659, cg11887188, cg16465939, cg08741300, cg11700071, cg21130221, cg05438727, cg05993525, cg07618453, cg06485603 | 4.62 × 10−3 – 9.5 × 10−3 | This gene encodes a voltage-gated potassium channel required for the repolarization phase of the cardiac action potential. |
| PTPRN2 (N = 16, chr 17) | Hypo: cg04694534, cg06679384, cg13218485, cg01726608, cg12138780, cg18395809, cg16486501, cg06649856, cg15272173, cg04799270 Hyper: cg09656639, cg26208815, cg17291423, cg23634928, cg04007350, cg02704570 | 2.35 × 10−3 – 7.75 × 10−3 | Protein tyrosine phosphatase, receptor type N2 encodes a major islet autoantigen in type-1 diabetes. PTPRN2 plays an important role in the epigenetic regulation of metabolic diseases and cancers. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honkova, K.; Rossnerova, A.; Chvojkova, I.; Milcova, A.; Margaryan, H.; Pastorkova, A.; Ambroz, A.; Rossner, P., Jr.; Jirik, V.; Rubes, J.; et al. Genome-Wide DNA Methylation in Policemen Working in Cities Differing by Major Sources of Air Pollution. Int. J. Mol. Sci. 2022, 23, 1666. https://doi.org/10.3390/ijms23031666
Honkova K, Rossnerova A, Chvojkova I, Milcova A, Margaryan H, Pastorkova A, Ambroz A, Rossner P Jr., Jirik V, Rubes J, et al. Genome-Wide DNA Methylation in Policemen Working in Cities Differing by Major Sources of Air Pollution. International Journal of Molecular Sciences. 2022; 23(3):1666. https://doi.org/10.3390/ijms23031666
Chicago/Turabian StyleHonkova, Katerina, Andrea Rossnerova, Irena Chvojkova, Alena Milcova, Hasmik Margaryan, Anna Pastorkova, Antonin Ambroz, Pavel Rossner, Jr., Vitezslav Jirik, Jiri Rubes, and et al. 2022. "Genome-Wide DNA Methylation in Policemen Working in Cities Differing by Major Sources of Air Pollution" International Journal of Molecular Sciences 23, no. 3: 1666. https://doi.org/10.3390/ijms23031666