1. Introduction
A crucial step in fertilization is the sperm–oocyte contact that allows the two gametes to fuse and create a zygote. Many questions remain open about the molecules that bring the gametes’ membranes together. For several years, only a few proteins were considered essential for gamete adhesion/fusion: first, the CD9 tetraspanin [
1,
2,
3], then the sperm IZUMO1 immunoglobulin [
4] and the GPI-AP JUNO, which is the IZUMO1 oocyte receptor [
5]. Over the past two years, six sperm factors essential for mammalian fertilization, SPACA6, TMEM95, SOF1, FIMP and DCST1/DCST2, have been identified using the CRISPR /Cas9 approach [
6,
7,
8,
9] (reviewed in [
10]). However, this multiplication of factors does not seem to be sufficient to explain the gamete adhesion/fusion process, first because the mechanisms of action of these different actors are not fully known, particularly with regard to the most recently identified molecules, and because there are certainly other molecules to be discovered. Very recently, MAIA, also known as Fc receptor-like 3, has been demonstrated to supersede JUNO as IZUMO1 receptor during human fertilization [
11]. These latest findings reinforce the idea of the existence of large and organized molecular complexes on each of the gamete plasma membranes, encouraging not only the identification of new players but also the reconsideration of some that have been sidelined. Integrins are included in the latter category during the fertilization process. Indeed, these were first considered essential; in particular α6β1 integrin has been described as a receptor for sperm ADAMs [
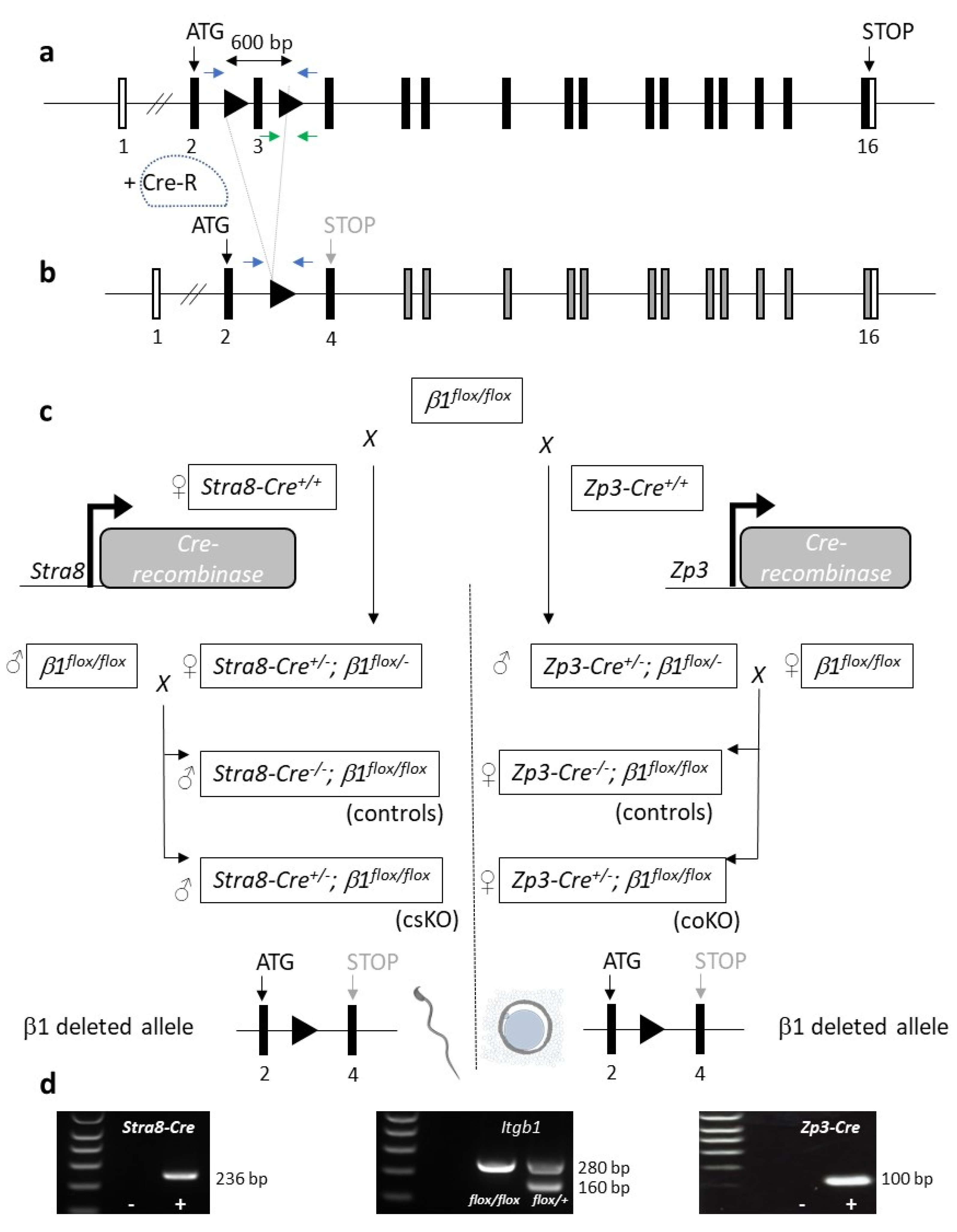
12]. Conditional invalidation experiments with the
Itga6 gene [
13] and then of the
Itgb1 gene [
14] at the oocyte level showed that they were dispensable since the oocytes deleted from one or the other of these genes were fertilizable and female mice carrying one of these deletions were normally fertile.
In order to understand this apparent contradiction, we hypothesized that the presence of the β1 integrin subunit could be necessary on the membrane of one of the two gametes as occurs in myoblasts or other cells that fuse [
15].
At first, we used synaptonemal complex protein 1 (
Sycp1)-Cre and integrin subunit beta1 (
Itgb1) floxed gene mice mating to generate sperm conditional knockout (KO) mouse, but this mouse did not show good excision efficiency at the point where males reproduced normally, with either wild-type (WT) or
Itgb1 KO oocyte (
Zp3-Cre +/− Itgb1 flox/flox) females, although in vitro, sperm showed a low fertilization rate compared with controls.
Zp3-Cre, where the Cre is expressed under the control of the promoter of the
Zp3 gene encoding one of the three proteins of the zona pellucida (ZP) in mice, is highly effective as described [
14], and accordingly, we never found integrin β1 protein on conditional KO oocytes. On the contrary, integrin β1 protein on the sperm of this
Sycp1-Cre +/− Itgb1 flox/flox model, that is a knockdown (KD), could still be detected with immunofluorescence and western blot [
16]. We have identified two elements that could explain the presence of residual integrin β1 protein on KD sperm: first, the low efficiency of the Cre recombinase expressed under the control of the
Sycp1 promoter, especially when combined with the distance between the two Lox sites (~28 kb) in the first used
Itgb1 floxed mice. Therefore, the question of whether the role of integrin β1 is essential or not in fertilization remains unanswered.
Here, we used a
Stra8-Cre mouse [
17] and a
Itgb1 floxed gene mouse with a smaller distance (~0.6 kb) between the two LoxP sites [
18]. Even though conditional KO (cKO) males continued to reproduce with a small decrease in the limit of significance when mated with WT females, there were no births when cKO males were mated with
Itgb1 KO females. On ultrasonography, we observed the presence of embryos and their peri-implantation lethality phenotype, which was previously described for total
Itgb1 gene KO [
19,
20]. Our findings indicate that integrin β1 is not essential for fertilization at the point of sperm–oocyte adhesion/fusion. However, there is no doubt about integrin β1′s participation during this process, as in its absence, in vitro fertilization rates were low, and sperm accumulated in the perivitelline space (PVS). In order to explain the present results, we propose that integrin β1could be resubstituted by another β integrin subunit, resulting in either another known integrin heterodimer or a novel dimerization. Nevertheless, these putative integrin heterodimers lacking β1 seem not functional during postimplantation embryo development, confirming that β1 integrin is essential during postimplantation.
3. Discussion
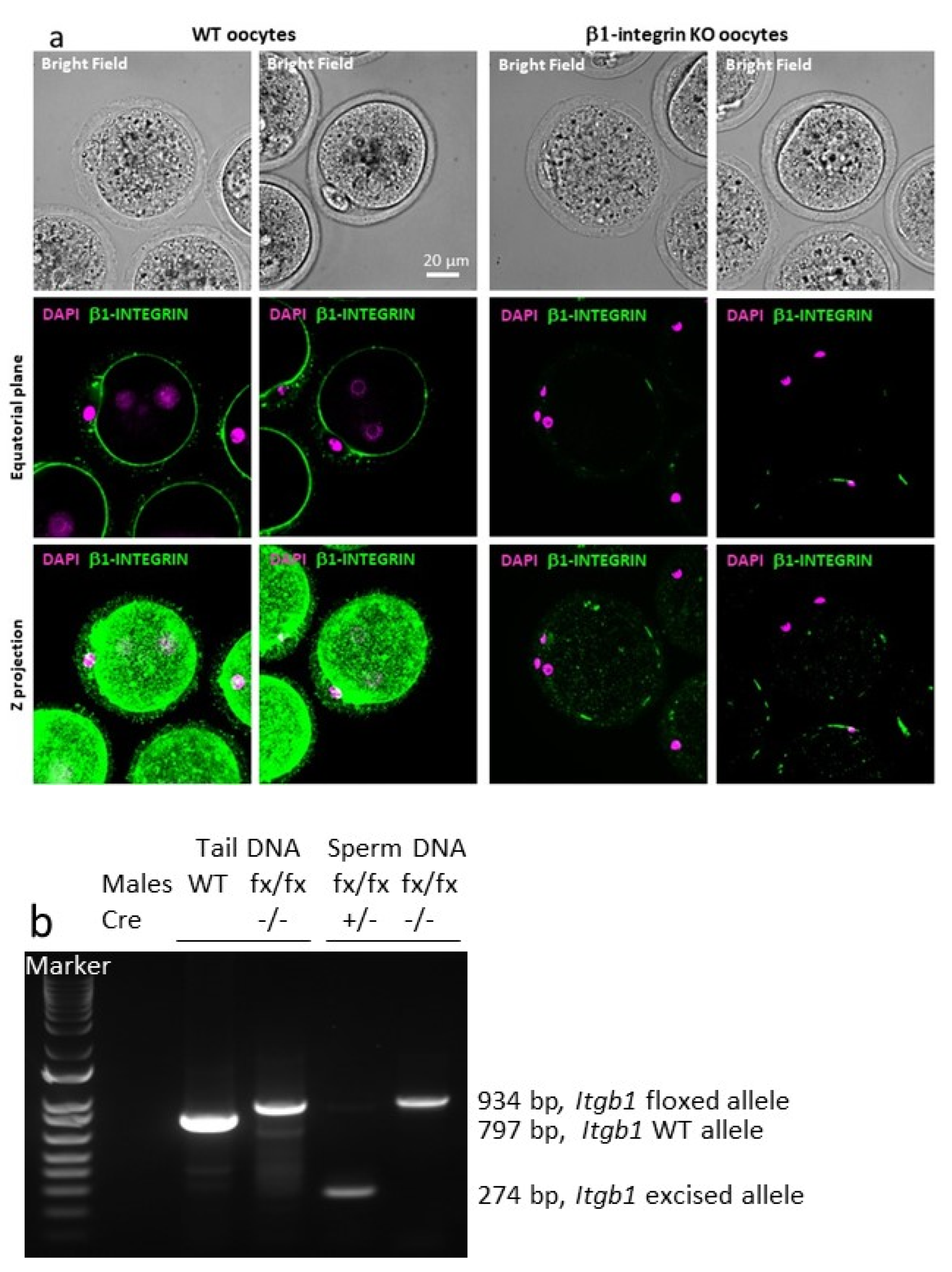
To study the function of integrin β1 during in vivo fertilization, we intended to generate
Itgb1 sperm conditional KO mice. Due to the partial efficiency of the Cre activity under the control of the promoter of the male germ cells specific gene,
Sycp1, we obtained a
Itgb1 Knock-down mouse line in the first place. Therefore, we could not clearly determine the role of integrin β1 in fertilization. It was suggested that the inefficiency of the Cre was due to the distance between the two LoxP sites (~28 kb).Therefore, we used a
Stra8-Cre mouse [
17] and another
Itgb1 floxed gene mouse with a smaller distance (~0.6 kb) between LoxP sites [
18]. This developed mouse line was also used in crossing with the
Zp3-Cre mouse line to obtain oocyte-conditional KO mice. The effectiveness of Cre under
Zp3 promoter control was already shown by us and others [
14,
16], and we further confirmed it by immunofluorescence. Indeed, oocytes from more than 10 conditional KO mice were tested, and 100% of the oocytes showed no integrin β1 signal, unlike the controls. For the males, we performed PCR on sperm DNA. Indeed, the male germ line is the only one to have both the floxed β1 alleles and the Cre expressed under the control of the
Stra8 promoter. PCR showed only the presence of the excised alleles indicating a very high efficiency of the Cre recombinase.
At this point, this
Stra8-Cre +/− Itgb1 flox/flox male model appeared to be the right tool for revealing the function of sperm integrin β1 during in vivo fertilization. The fact that
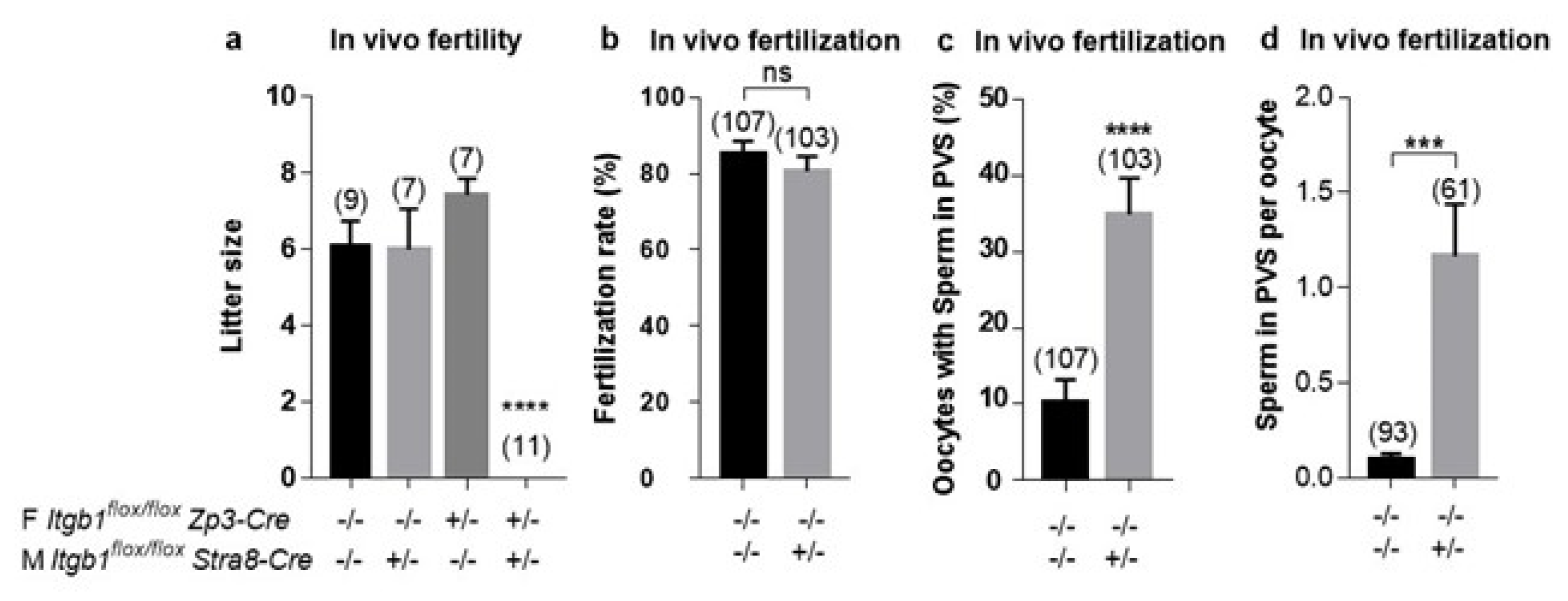
Itgb1-conditional KO males continued to reproduce normally when they were mated with WT females indicated either that sperm integrin β1 was not necessary for in vivo fertilization or that oocyte integrin β1 could compensate for sperm-originated integrin β1 absence. In order to test the oocyte compensation hypothesis, we mated sperm-conditional KO males with
Itgb1-oocyte KO females. Indeed, we did not obtain any births. Here again, two explanations were possible: the observed sterility could have been due to either a defect in fertilization or a defect in development. Using the same mating scheme, we retrieved oocytes from females that exhibited a vaginal plug the day after mating. The fertilization rates were the same in the KO group as in the controls, thus demonstrating that the absence of the integrin β1 subunit in both sperm and oocytes was not an obstacle to fertilization. The only anomaly that we noted was the presence of spermatozoa in the PVS, with a higher frequency than in the controls. This phenotype indicated that sperm lacking integrin β1 could take longer to fuse with the oocyte. The possible delay in triggering the cortical reaction, which normally follows fusion and prevents polyspermy [
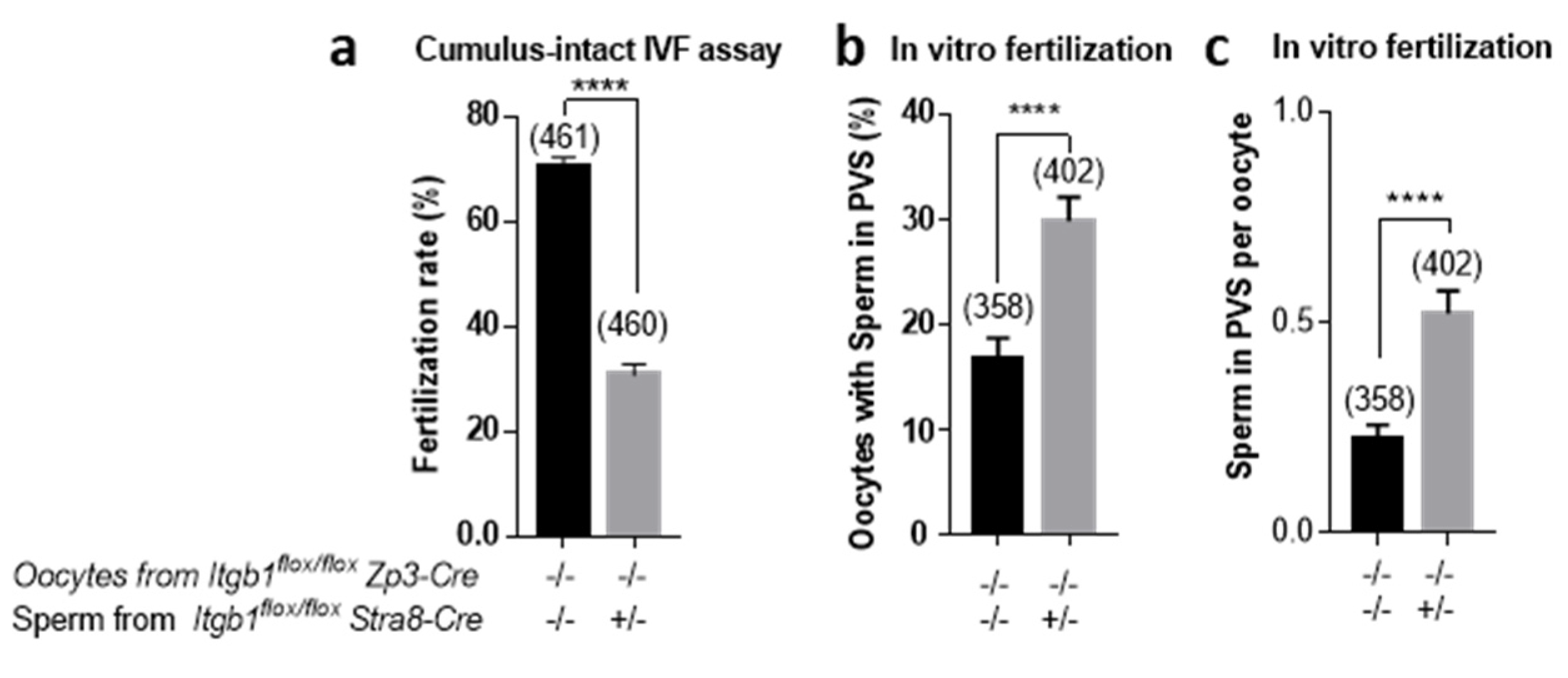
21], could explain the entry of additional sperm into PVS. These results are in favor of a theory of participation of β1 integrin in fertilization in terms of optimization. In support of this hypothesis, in vitro fertilization, where conditions are not optimal as in vivo, showed a significant decrease in fertilization rates when control oocytes were inseminated by cKO sperm, in addition to a greater accumulation of cKO spermatozoa in PVS.
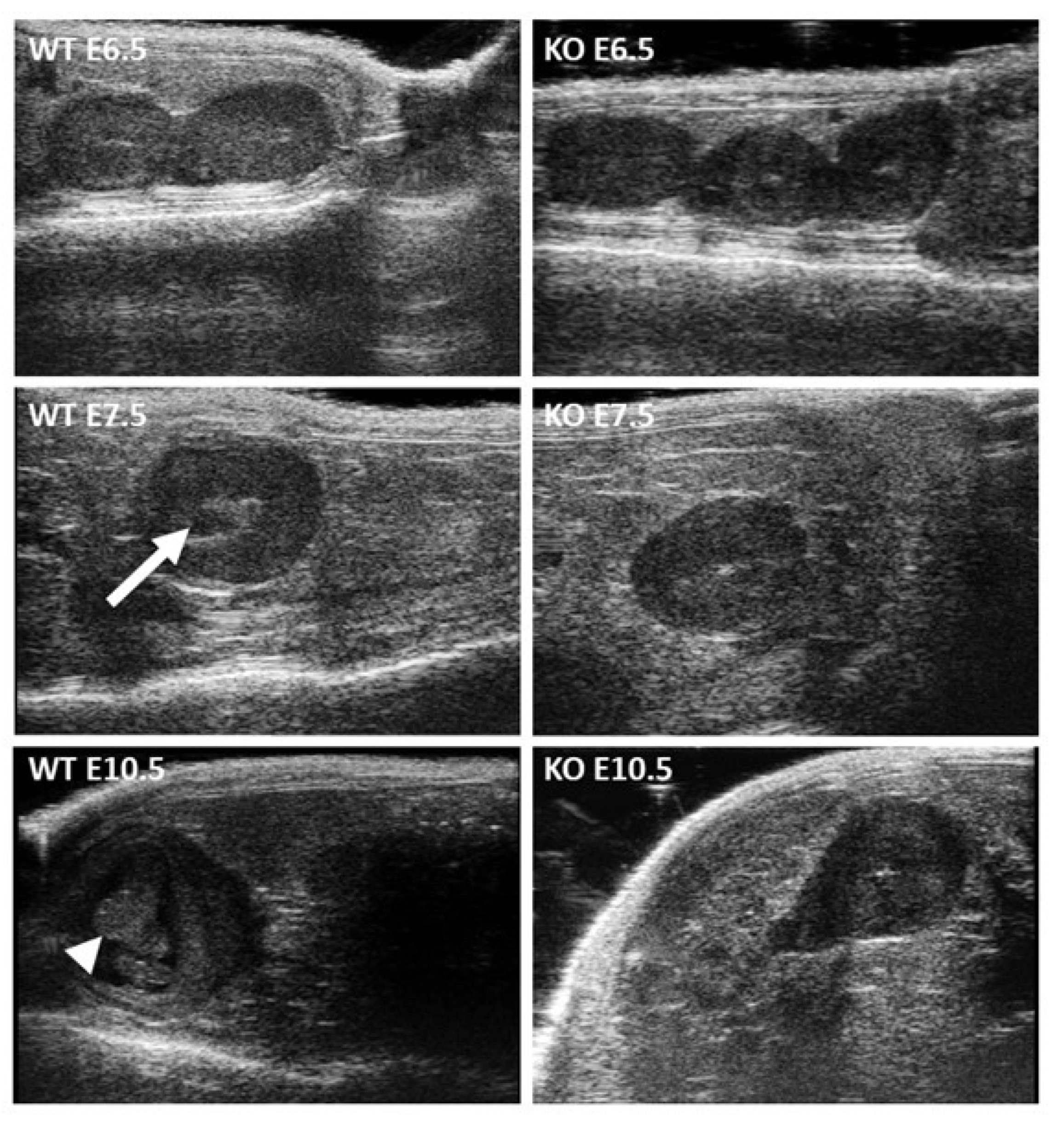
The hypothesis that remained to be tested in order to explain sterility was that of embryonic lethality. With ultrasonography, we observed the presence of embryos and their peri-implantation lethality phenotype, which was previously described for total
Itgb1 gene KO [
19,
20]. These findings indicated two points. First, the Cre under the control of the
Stra8 promoter was more efficient than that under the control of the
Sycp1 promoter that we had used before [
16]. Among the thirty pups generated for ultrasonography experiments, all resorbed; that is, none had inherited an intact (still floxed)
Itgb1 gene. Second, integrin β1 participates as well, but it is not essential for fertilization to take place, at least not in vivo. A hypothesis that could explain this apparent contradiction is related to a possible redundancy either by the de novo expression of another beta integrin subunit or by a new dimerization in the absence of the β1 integrin subunit. This hypothesis could also explain why the in vitro fertilization rate was lower when Cre was less effective using the
Sycp1-Cre mouse line [
16]. We propose that when Cre is strongly efficient (
Stra8-Cre), there is low or no integrin β1 mRNA and protein expression, setting up up a functional compensatory integrin heterodimer system in the absence of integrin β1 subunit. Further investigations are needed to verify this redundancy hypothesis. For example, comparative studies of the transcriptome and/or the proteome of gametes (oocytes and spermatozoa) lacking beta1 and control gametes that express integrin β1 could make it possible to detect genes or proteins expressed only in the context of the absence of integrin β1 subunit.
4. Materials and Methods
4.1. Ethics Statement
All animal experiments were performed in accordance with national guidelines for the care and use of laboratory animals. Authorizations were obtained from local (C2EA-34, Comité d’éthique en matière d’expérimentation animale Paris Descartes) and governmental ethical review committees via APAFiS Application (Autorisation de projet utilisant des animaux à des fins scientifiques), Authorization APAFIS #14124-2017072510448522 v26, A. Ziyyat (2018–2023).
4.2. Generation of Oocyte and Sperm Itgb1 Conditional Knockout Mice
The floxed
Itgb1 gene mice [
18] (JAX stock #004605) were mated with the transgenic mice expressing the Cre recombinase under the control of the
Zp3 promoter [
14,
22] (JAX stock #003651) or the
Stra8 promoter [
17] (JAX stock #017490). Heterozygous mice for
floxed Itgb1 and
Zp3-Cre or
Stra8-Cre were mated with
floxed Itgb1 mice to generate mice homozygous for the floxed
Itgb1 gene and expressing Cre. Female mice with both the
Zp3-Cre transgene and homozygous floxed
Itgb1 gene (
Zp3-Cre +/− Itgb1 flox/flox) were used for in vivo mating. Male mice with the
Stra8-Cre transgene and homozygous floxed
Itgb1 gene (
Stra8-Cre +/− Itgb1 flox/flox) were used for in vivo mating and IVF assays. For all experiments, controls used were
Stra8- or
Zp3-
Cre −/− Itgb1 flox/flox males or females respectively.
Genotyping of mice was performed by PCR amplification (GoTaq
® DNA Polymerase, Promega, Madison, WI, USA) on DNA extracted from tail biopsies (NucleoSpin
® Tissue, Macherey-Nagel, Düren, Germany) using the following primers: 5′- CGGCTCAAAGCAGAGTGTCAGTC -3′ and 5′- CCACAACTTTCCCAGTTAGCTCTC-3′ for floxed
Itgb1 gene detection and one or the other of these two pairs of primers: 5′- GCGGTCTGGCAGTAAAAACTATC -3′ and 5′- GTGAAACAGCATTGCTGTCACTT-3′ or 5′- AGATGCCAGGACATCAGGAACCTG-3′ and 5‘-ATCAGCCACACCAGACACAGAGATC-3′ for
Cre gene detection under the control of
Zp3 or
Stra8 promoters. The first primer pair gives an amplification of 160 bp for WT
Itgb1 and 280 bp for floxed
Itgb1 (
Figure 1a, green arrows). The second and the third ones give amplimers of 100 bp or 236 bp respectively.
4.3. Immunofluorescence of Mouse Embryos
Zona-intact early embryos from control or conditional KO mice were fixed during 20 min in 4% formaldehyde at room temperature (RT) and washed three times in PBS-1% BSA. For detection of the integrin β1, embryos were incubated with the primary antibody (MB1.2 at 20 µg/mL; Chemicon International, Temecula, CA, USA) for 1 h at RT followed by Alexa Fluor® 488 goat anti-rat IgG secondary antibody incubation (10 µg/mL, Molecular Probes, Invitrogen, Illkirch, France). Embryos were then incubated with 5µg/mL Hoechst 33342 (Invitrogen H3570) in PBS-1% BSA for 30 min and washed in PBS-1% BSA and mounted in PBS in Nunc™ Lab-Tek™ Chamber Slide System (Thermo Fisher Scientific, Illkirch, France). Control immunofluorescent studies were performed using isotype (IgG2a) (Serotec-Bio-Rad, Marnes-la-Coquette, France) as primary antibody or Alexa Fluor® 488 goat anti-rat IgG alone. Observations and acquisitions were carried out on an inverted spinning disk microscope using oil-immersion objectives X40. Final images were processed, Z-projected upon request, and merged using ImageJ.
4.4. PCR on Mouse Tail and Sperm DNA
DNA was extracted from sperm pellets and mouse tail biopsies (NucleoSpin
® Tissue, Macherey-Nagel, Düren, Germany). The amplification was performed thanks to primers located upstream of the LoxP5′ in intron 2 (5′-TATAACCCGCAGAACAATAGG-3′) and downstream of LoxP3′ in intron 3 (5′-CCACAACTTTCCCAGTTAGCTCTC-3′) and GoTaq
® DNA Polymerase (
Figure 1a, blue arrows). Amplimers of 797, 934 or 274 bp were obtained for the WT, floxed or excised allele respectively.
4.5. In Vivo Mating, Gamete Preparation and in vitro Fertilization
Four groups of mating were used: KO males (i.e., males expressing Cre under the control of the Stra8 promoter at the heterozygous state) with KO females (i.e., females expressing Cre under the control of the Zp3 promoter at the heterozygous state), KO males with control females, control males with KO females and control males with control females. All mice were homozygous for the Itgb1 floxed gene. Mice were aged from 8 to 14 weeks and were housed as one male and one female per cage. In each group, litter size was assessed and compared to other groups.
WT and conditional KO female mice (5–8 week-old) were superovulated with 5 IU pregnant mare serum gonadotropin (PMSG) and 5 IU human chorionic gonadotropin (hCG, Intervet, Beaucouze, France) 48 h apart. Fourteen hours after hCG injection, animals were sacrificed by cervical dislocation. Cumuli oophori were collected by tearing the ampulla wall of the oviduct, placed in Ferticult medium (FertiPro, Belgium) supplemented with 3% bovine serum albumin (BSA) and maintained at 37 °C under 5% CO2 in air under mineral oil (Sigma, St. Louis, MO, USA). For zona-free in vitro fertilization (IVF) assay, oocytes were freed from the cumulus cells by 3–5 min incubation at 37 °C with hyaluronidase (Sigma) in M2 medium (Sigma). Oocytes were rinsed and kept in Ferticult medium at 37 °C under 5% CO2 atmosphere under mineral oil. Zona pellucida (ZP) was then dissolved with acidic Tyrode’s (AT) solution (pH 2.5, Sigma) under visual monitoring. The zona-free eggs were rapidly washed in medium and kept at 37 °C under 5% CO2 atmosphere for 2 to 3 h to recover their fertilizability.
Mouse spermatozoa were obtained from the cauda epididymis of control or KO conditional male mice (8 to 14 weeks old) and capacitated at 37 °C under 5% CO2 for 90 min in a 500 µL drop of Ferticult medium supplemented with 30 mg/mL BSA, under mineral oil.
Cumulus-intact or zona-free oocytes were inseminated for 3 h in a 50 µL drop of medium with capacitated spermatozoa at a final concentration of 1 × 106/mL or 1 × 105/mL respectively. Then, they were washed and directly mounted in Vectashield/DAPI for observation under UV light (Zeiss Axioskop 20 microscope). They were considered fertilized when the oocytes showed a fluorescent decondensed sperm head within their cytoplasm.
4.6. High Frequency Ultrasonography Implantation and Survival Assessment
Gestations were obtained by crossing oocyte Itgb1-conditional KO females (Zp3-Cre +/− Itgb1 flox/flox) with sperm Itgb1-conditional KO males (Stra8-Cre +/− Itgb1 flox/flox). Females were used to collect phenotypic data from the gestation. Females from the control group (Zp3- Cre −/− Itgb1 flox/flox) were crossed with control males (Stra8-Cre −/− Itgb1 flox/flox). Implantation and survival rates were obtained using high-frequency (40 MHz) ultrasonography (VEVO 2100 with MS-550D probe, Visulasonics, Toronto, Canada). Briefly, a chemical hair remover was used to eliminate abdominal hair. Ultrasonographic contact gel was used to ensure contact between the skin surface and the transducer. Body temperature, electrocardiographic and respiratory profiles were monitored using ultrasound device’s integrated heating pad and monitoring device (THM150, Indus Instruments, Webster, TX, USA). The implantation and the survival rates were determined early in the gestation, at E6.5, E7.5 and E10.5. At these stages, the small size of embryos permits a fluent count and resorbed embryos are also visible. During each examination, the number of implanted embryos in each uterine horn as well as their status (alive or dead) were assessed.
4.7. Statistical Analysis
Results are expressed as mean ± sem of at least three independent experiments. For statistical analysis, one-way ANOVA multiple-comparisons test or t-test were performed using GraphPad Prism version 7.00 for Windows (GraphPad Software, La Jolla California, CA, USA). Differences were considered statistically significant when p < 0.05.



 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}