
H4K5 Butyrylation Coexist with Acetylation during Human Spermiogenesis and Are Retained in the Mature Sperm Chromatin
 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
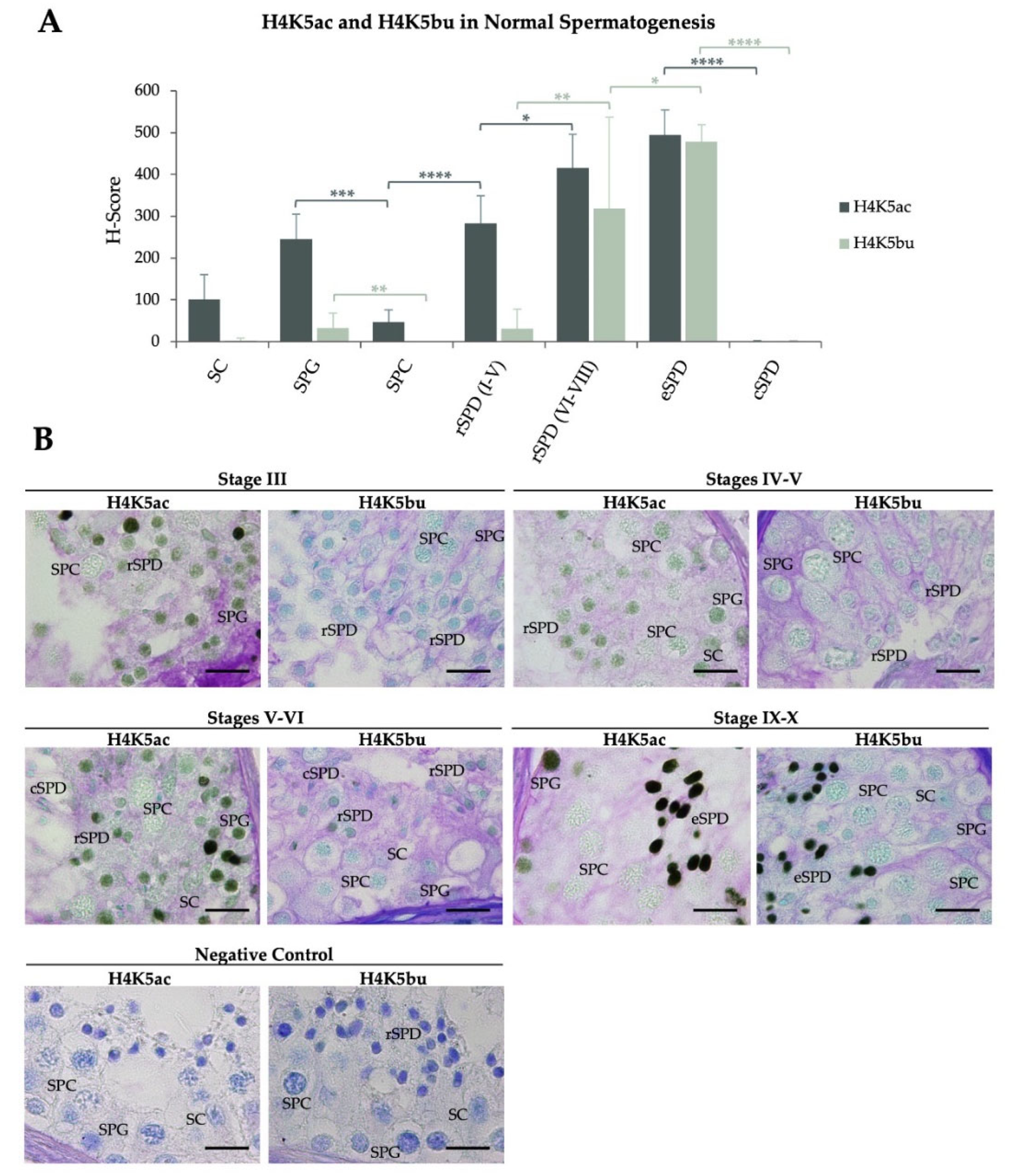
2.1. H4K5 Butyrylation Show Different Spermatogenic Pattern to Acetylation on the Same Residue in Patients with Normal Spermatogenesis
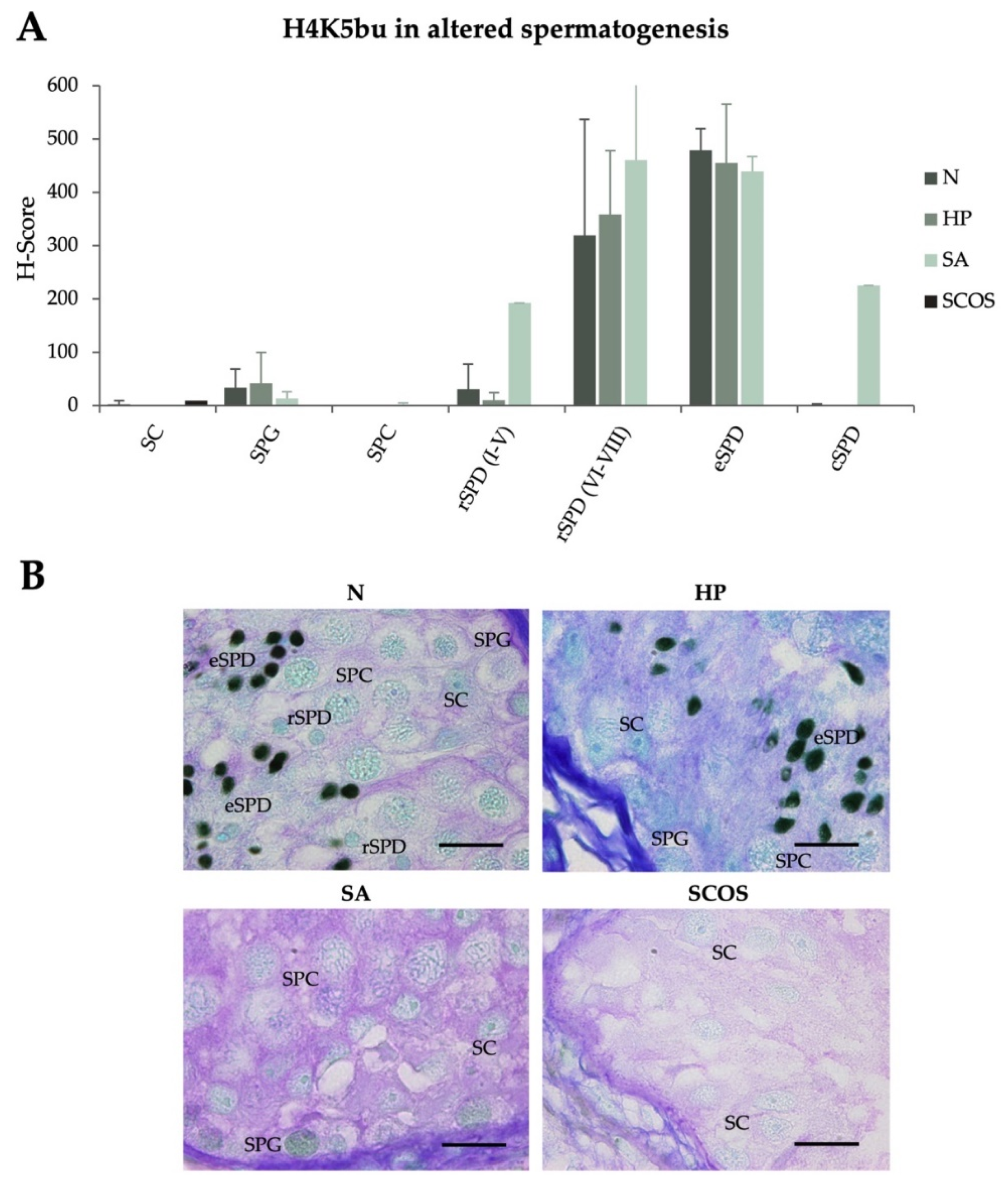
2.2. H4K5 Butyrylation Spermatogenic Levels Are Not Impaired under Different Testicular Defects
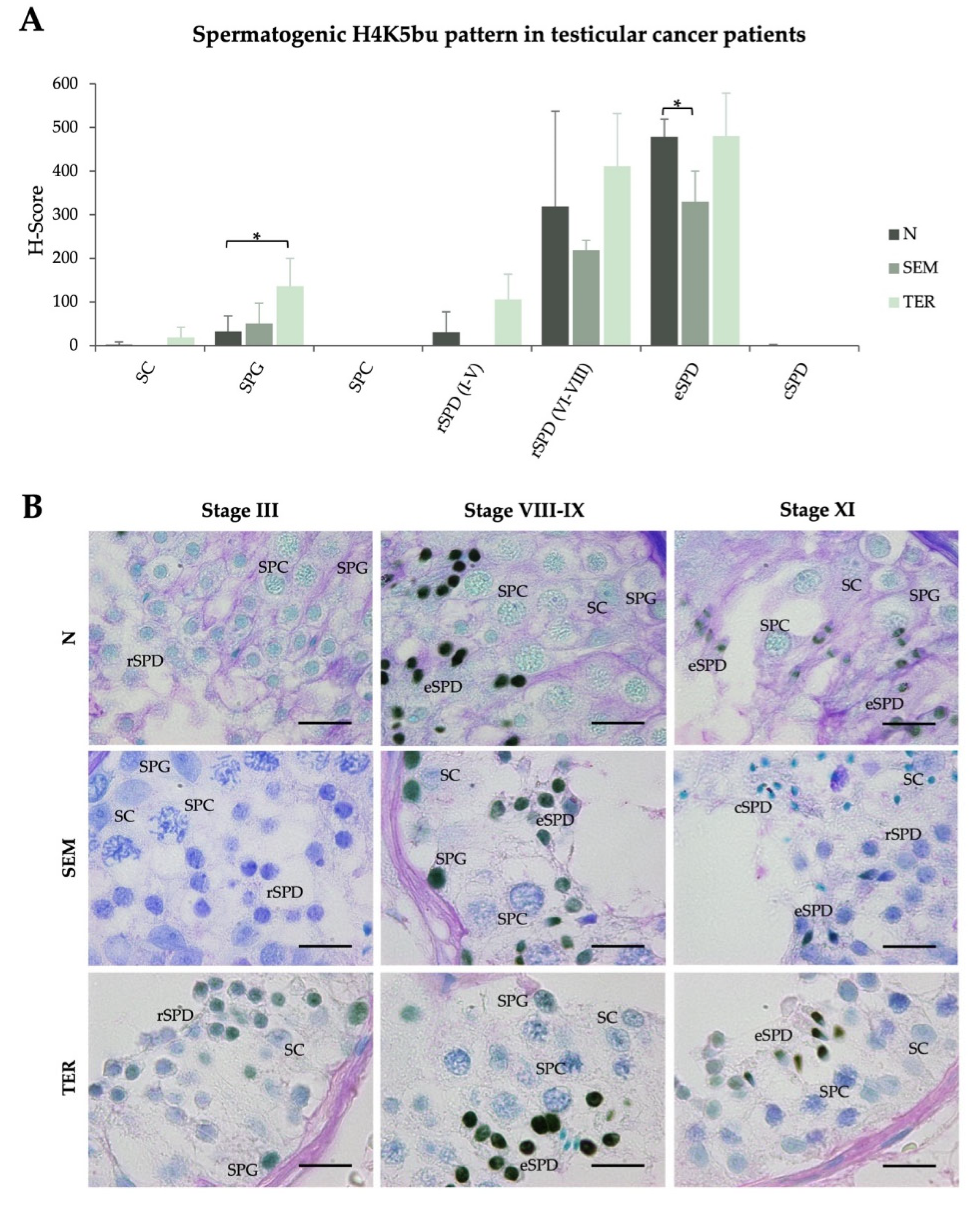
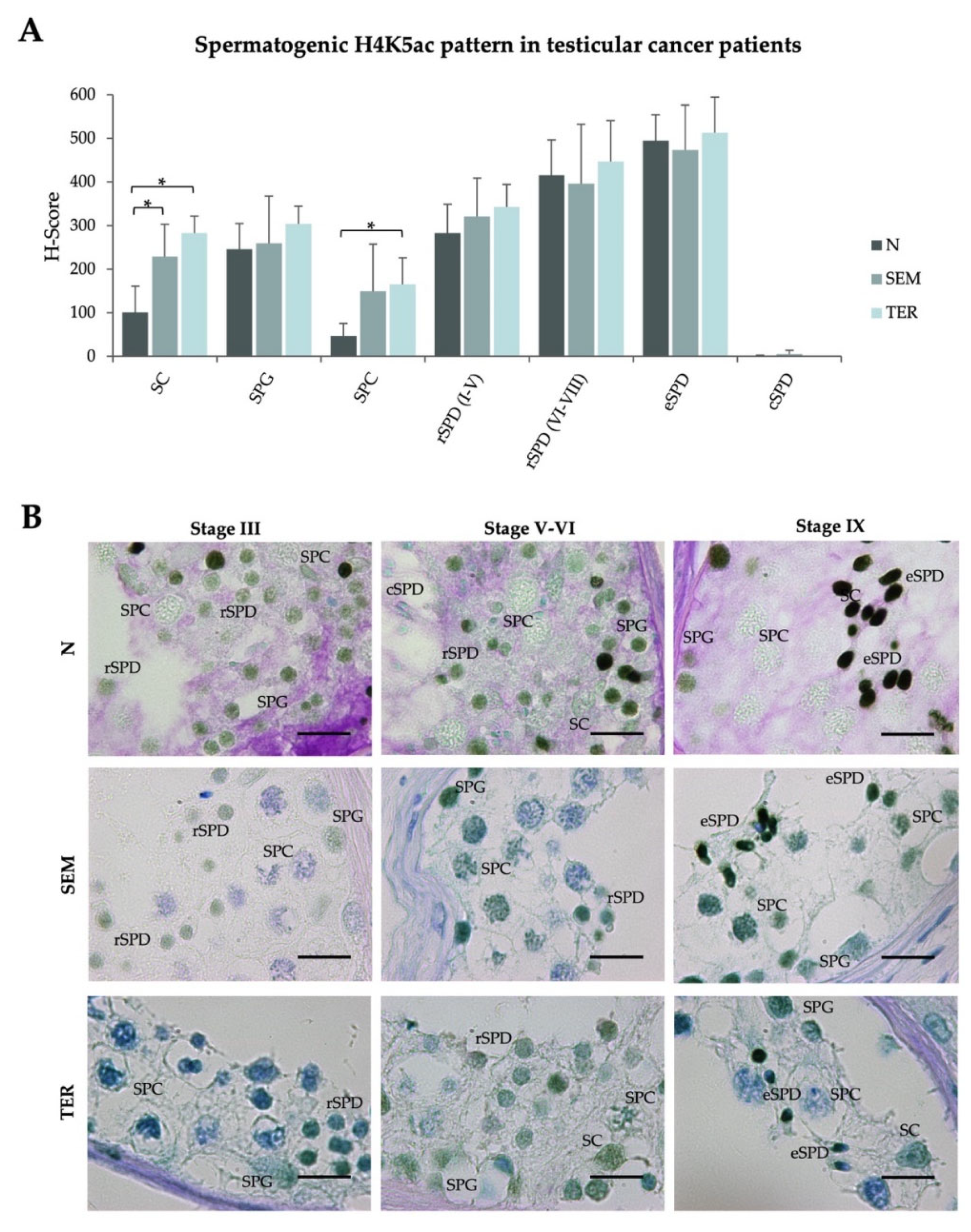
2.3. Testicular Cancers Do Not Affect the Spermatogenic Pattern of Butyrylation on H4K5, but Neither Acetylation on the Same Residue, in Healthy Seminiferous Tubules with Complete Spermatogenesis
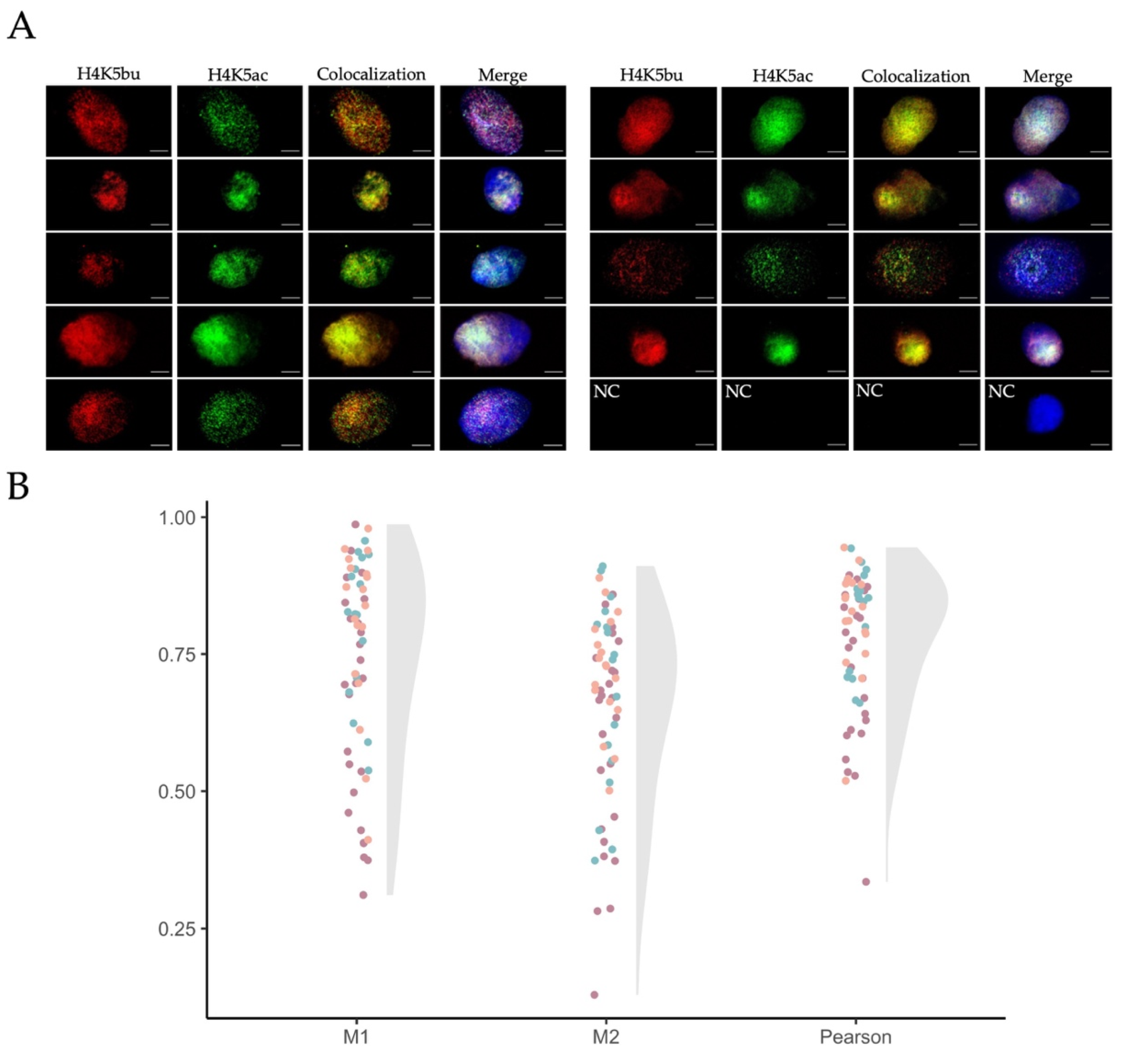
2.4. Butyrylation and Acetylation on H4K5 Are Retained in Ejaculated Human Sperm from Normozoospermic Patients
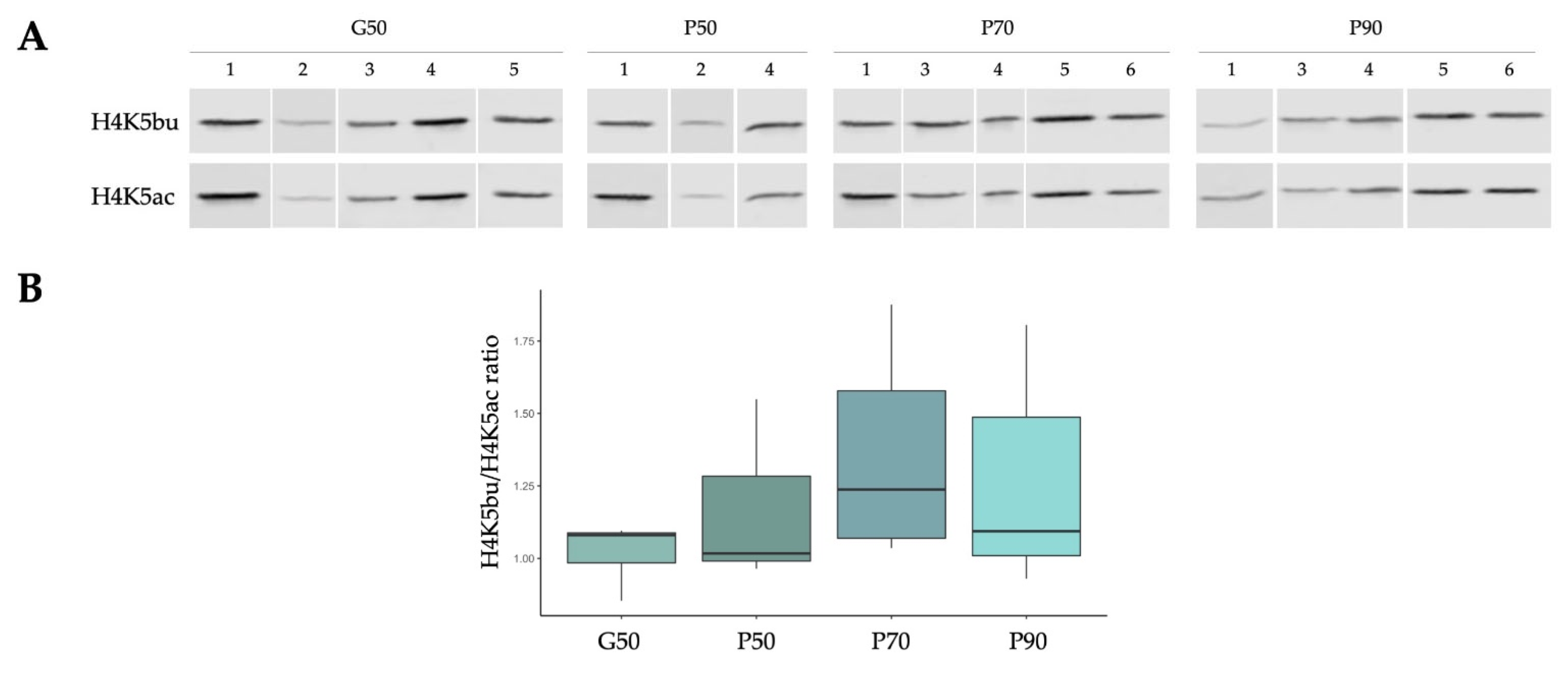
2.5. The Proportion of H4K5bu over H4K5ac Remains Constant within Different Sperm Populations Selected According to Sperm Quality
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.1.1. Testicular Biopsies
4.1.2. Semen samples
4.2. Processing and Paraffin-Embedding of the Testicular Material
4.3. Testicular Histopathological Evaluation
4.4. Immunohistochemistry
4.5. Spermatogenic Stage Classification and IHC Evaluation
4.6. Immunofluorescence on Mature Sperm Cells
4.7. Colocalization Analyses
4.8. Odyssey® Western Blot
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Mateo, S.; Castillo, J.; Estanyol, J.M.; Ballescà, J.L.; Oliva, R. Proteomic Characterization of the Human Sperm Nucleus. Proteomics 2011, 11, 2714–2726. [Google Scholar] [CrossRef] [PubMed]
- Hammoud, S.; Nix, D.A.; Hammoud, A.O.; Gibson, M.; Cairns, B.R.; Carrell, D. Genome-Wide Analysis Identifies Changes in Histone Retention and Epigenetic Modifications at Developmental and Imprinted Gene Loci in the Sperm of Infertile Men. Hum. Reprod. 2011, 26, 2558–2569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, R.; Luís Ballescà, J. Altered Histone Retention and Epigenetic Modifications in the Sperm of Infertile Men. Asian J. Androl. 2012, 14, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Azpiazu, R.; Amaral, A.; Castillo, J.; Estanyol, J.M.; Guimerà, M.; Ballescà, J.L.; Balasch, J.; Oliva, R. High-Throughput Sperm Differential Proteomics Suggests That Epigenetic Alterations Contribute to Failed Assisted Reproduction. Hum. Reprod. 2014, 29, 1225–1237. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.; Amaral, A.; Azpiazu, R.; Vavouri, T.; Estanyol, J.M.; Ballesca, J.L.; Oliva, R. Genomic and Proteomic Dissection and Characterization of the Human Sperm Chromatin. Mol. Hum. Reprod. 2014, 20, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.; Estanyol, J.; Ballescà, J.; Oliva, R. Human Sperm Chromatin Epigenetic Potential: Genomics, Proteomics, and Male Infertility. Asian J. Androl. 2015, 17, 601. [Google Scholar] [CrossRef]
- Barrachina, F.; Soler-Ventura, A.; Oliva, R.; Jodar, M. Sperm Nucleoproteins (Histones and Protamines). In A Clinician’s Guide to Sperm DNA and Chromatin Damage; Zini, A., Agarwal, A., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 31–51. [Google Scholar]
- Oliva, R.; Dixon, G.H. Vertebrate Protamine Genes and the Histone-to-Protamine Replacement Reaction. Prog. Nucleic Acid Res. Mol. Biol. 1991, 40, 25–94. [Google Scholar] [CrossRef]
- Christensen, M.E.; Dixon, G.H. Hyperacetylation of Histone H4 Correlates with the Terminal, Transcriptionally Inactive Stages of Spermatogenesis in Rainbow Trout. Dev. Biol. 1982, 93, 404–415. [Google Scholar] [CrossRef]
- Oliva, R.; Mezquita, C. Histone H4 Hyperacetylation and Rapid Turnover of Its Acetyl Groups in Transcriptionally Inactive Rooster Testis Spermatids. Nucleic Acids Res. 1982, 10, 8049–8059. [Google Scholar] [CrossRef] [Green Version]
- Grimes, S.R.; Henderson, N. Hyperacetylation of Histone H4 in Rat Testis Spermatids. Exp. Cell Res. 1984, 152, 91–97. [Google Scholar] [CrossRef]
- Oliva, R.; Bazett-Jones, D.; Mezquita, C.; Dixon, G.H. Factors Affecting Nucleosome Disassembly by Protamines in Vitro. Histone Hyperacetylation and Chromatin Structure, Time Dependence, and the Size of the Sperm Nuclear Proteins. J. Biol. Chem. 1987, 262, 17016–17025. [Google Scholar] [CrossRef]
- Hazzouri, M.; Pivot-Pajot, C.; Faure, A.-K.; Usson, Y.; Pelletier, R.; Sèle, B.; Khochbin, S.; Rousseaux, S. Regulated Hyperacetylation of Core Histones during Mouse Spermatogenesis: Involvement of Histone-Deacetylases. Eur. J. Cell Biol. 2000, 79, 950–960. [Google Scholar] [CrossRef]
- Oliva, R.; Castillo, J. Proteomics and the Genetics of Sperm Chromatin Condensation. Asian J. Androl. 2011, 13, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Oliva, R.; Bazett-Jones, D.P.; Locklear, L.; Dixon, G.H. Histone Hyperacetylation Can Induce Unfolding of the Nucleosome Core Particle. Nucleic Acids Res. 1990, 18, 2739–2747. [Google Scholar] [CrossRef] [Green Version]
- Singh, I.; Parte, P. Heterogeneity in the Epigenetic Landscape of Murine Testis-Specific Histone Variants TH2A and TH2B Sharing the Same Bi-Directional Promoter. Front. Cell. Dev. Biol. 2021, 9, 755751. [Google Scholar] [CrossRef]
- Pentakota, S.K.; Sandhya, S.; Sikarwar, A.P.; Chandra, N.; Satyanarayana Rao, M.R. Mapping Post-Translational Modifications of Mammalian Testicular Specific Histone Variant TH2B in Tetraploid and Haploid Germ Cells and Their Implications on the Dynamics of Nucleosome Structure. J. Proteome Res. 2014, 13, 5603–5617. [Google Scholar] [CrossRef]
- Ding, D.; Nguyen, T.T.; Pang, M.Y.H.; Ishibashi, T. Primate-Specific Histone Variants. Genome 2021, 64, 337–346. [Google Scholar] [CrossRef]
- Hoghoughi, N.; Barral, S.; Vargas, A.; Rousseaux, S.; Khochbin, S. Histone Variants: Essential Actors in Male Genome Programming. J. Biochem. 2018, 163, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Barral, S.; Morozumi, Y.; Tanaka, H.; Montellier, E.; Govin, J.; de Dieuleveult, M.; Charbonnier, G.; Couté, Y.; Puthier, D.; Buchou, T.; et al. Histone Variant H2A.L.2 Guides Transition Protein-Dependent Protamine Assembly in Male Germ Cells. Mol Cell 2017, 66, 89–101.e8. [Google Scholar] [CrossRef] [Green Version]
- Rezaei-Gazik, M.; Vargas, A.; Amiri-Yekta, A.; Vitte, A.-L.; Akbari, A.; Barral, S.; Esmaeili, V.; Chuffart, F.; Sadighi-Gilani, M.A.; Couté, Y.; et al. Direct Visualization of Pre-Protamine 2 Detects Protamine Assembly Failures and Predicts ICSI Success. Mol. Hum. Reprod 2022, 28, gaac004. [Google Scholar] [CrossRef]
- Dhar, S.; Thota, A.; Rao, M.R.S. Insights into Role of Bromodomain, Testis-Specific (Brdt) in Acetylated Histone H4-Dependent Chromatin Remodeling in Mammalian Spermiogenesis. J. Biol. Chem. 2012, 287, 6387–6405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudarzi, A.; Shiota, H.; Rousseaux, S.; Khochbin, S. Genome-Scale Acetylation-Dependent Histone Eviction during Spermatogenesis. J. Mol. Biol. 2014, 426, 3342–3349. [Google Scholar] [CrossRef] [PubMed]
- Her, Y.R.; Wang, L.; Chepelev, I.; Manterola, M.; Berkovits, B.; Cui, K.; Zhao, K.; Wolgemuth, D.J. Genome-Wide Chromatin Occupancy of BRDT and Gene Expression Analysis Suggest Transcriptional Partners and Specific Epigenetic Landscapes That Regulate Gene Expression during Spermatogenesis. Mol. Reprod. Dev. 2021, 88, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Manterola, M.; Brown, T.M.; Oh, M.Y.; Garyn, C.; Gonzalez, B.J.; Wolgemuth, D.J. BRDT Is an Essential Epigenetic Regulator for Proper Chromatin Organization, Silencing of Sex Chromosomes and Crossover Formation in Male Meiosis. PLoS Genet. 2018, 14, e1007209. [Google Scholar] [CrossRef] [Green Version]
- Morinière, J.; Rousseaux, S.; Steuerwald, U.; Soler-López, M.; Curtet, S.; Vitte, A.-L.; Govin, J.; Gaucher, J.; Sadoul, K.; Hart, D.J.; et al. Cooperative Binding of Two Acetylation Marks on a Histone Tail by a Single Bromodomain. Nature 2009, 461, 664–668. [Google Scholar] [CrossRef]
- Pivot-Pajot, C.; Caron, C.; Govin, J.; Vion, A.; Rousseaux, S.; Khochbin, S. Acetylation-Dependent Chromatin Reorganization by BRDT, a Testis-Specific Bromodomain-Containing Protein. Mol. Cell. Biol. 2003, 23, 5354–5365. [Google Scholar] [CrossRef] [Green Version]
- Shang, E.; Nickerson, H.D.; Wen, D.; Wang, X.; Wolgemuth, D.J. The First Bromodomain of Brdt, a Testis-Specific Member of the BET Sub-Family of Double-Bromodomain-Containing Proteins, Is Essential for Male Germ Cell Differentiation. Development 2007, 134, 3507–3515. [Google Scholar] [CrossRef] [Green Version]
- Sonnack, V.; Failing, K.; Bergmann, M.; Steger, K. Expression of Hyperacetylated Histone H4 during Normal and Impaired Human Spermatogenesis. Andrologia 2002, 34, 384–390. [Google Scholar] [CrossRef]
- Faure, A.K.; Pivot-Pajot, C.; Kerjean, A.; Hazzouri, M.; Pelletier, R.; Péoc’h, M.; Sèle, B.; Khochbin, S.; Rousseaux, S. Misregulation of Histone Acetylation in Sertoli Cell-only Syndrome and Testicular Cancer. Mol. Hum. Reprod. 2003, 9, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Kleiman, S.E.; Bar-Shira Maymon, B.; Hauser, R.; Botchan, A.; Paz, G.; Yavetz, H.; Yogev, L. Histone H4 Acetylation and AZFc Involvement in Germ Cells of Specimens of Impaired Spermatogenesis. Fertil. Steril. 2008, 89, 1728–1736. [Google Scholar] [CrossRef]
- Barrachina, F.; de la Iglesia, A.; Jodar, M.; Soler-Ventura, A.; Mallofré, C.; Rodriguez-Carunchio, L.; Goudarzi, A.; Corral, J.M.; Ballescà, J.L.; Castillo, J.; et al. Histone H4 Acetylation Is Dysregulated in Active Seminiferous Tubules Adjacent to Testicular Tumours. Hum. Reprod. 2022, 30, 1712–1726. [Google Scholar] [CrossRef]
- Crespo, M.; Damont, A.; Blanco, M.; Lastrucci, E.; Kennani, S.E.; Ialy-Radio, C.; El Khattabi, L.; Terrier, S.; Louwagie, M.; Kieffer-Jaquinod, S.; et al. Multi-Omic Analysis of Gametogenesis Reveals a Novel Signature at the Promoters and Distal Enhancers of Active Genes. Nucleic Acids Res. 2020, 48, 4115–4138. [Google Scholar] [CrossRef]
- Sabari, B.R.; Zhang, D.; Allis, C.D.; Zhao, Y. Metabolic Regulation of Gene Expression through Histone Acylations. Nat. Rev. Mol. Cell. Biol. 2017, 18, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.; Luo, H.; Lee, S.; Jin, F.; Yang, J.S.; Montellier, E.; Buchou, T.; Cheng, Z.; Rousseaux, S.; Rajagopal, N.; et al. Identification of 67 Histone Marks and Histone Lysine Crotonylation as a New Type of Histone Modification. Cell 2011, 146, 1016–1028. [Google Scholar] [CrossRef] [Green Version]
- Goudarzi, A.; Zhang, D.; Huang, H.; Barral, S.; Kwon, O.K.; Qi, S.; Tang, Z.; Buchou, T.; Vitte, A.-L.; He, T.; et al. Dynamic Competing Histone H4 K5K8 Acetylation and Butyrylation Are Hallmarks of Highly Active Gene Promoters. Mol. Cell 2016, 62, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Brykczynska, U.; Hisano, M.; Erkek, S.; Ramos, L.; Oakeley, E.J.; Roloff, T.C.; Beisel, C.; Schübeler, D.; Stadler, M.B.; Peters, A.H.F.M. Repressive and Active Histone Methylation Mark Distinct Promoters in Human and Mouse Spermatozoa. Nat. Struct. Mol. Biol. 2010, 17, 679–687. [Google Scholar] [CrossRef]
- Erkek, S.; Hisano, M.; Liang, C.-Y.; Gill, M.; Murr, R.; Dieker, J.; Schübeler, D.; van der Vlag, J.; Stadler, M.B.; Peters, A.H.F.M. Molecular Determinants of Nucleosome Retention at CpG-Rich Sequences in Mouse Spermatozoa. Nat. Struct. Mol. Biol. 2013, 20, 868–875. [Google Scholar] [CrossRef] [Green Version]
- Saida, M.; Iles, D.; Elnefati, A.; Brinkworth, M.; Miller, D. Key Gene Regulatory Sequences with Distinctive Ontological Signatures Associate with Differentially Endonuclease-Accessible Mouse Sperm Chromatin. Reproduction 2011, 142, 73–86. [Google Scholar] [CrossRef]
- Vavouri, T.; Lehner, B. Chromatin Organization in Sperm May Be the Major Functional Consequence of Base Composition Variation in the Human Genome. PLoS Genet. 2011, 7, e1002036. [Google Scholar] [CrossRef] [Green Version]
- Ihara, M.; Meyer-Ficca, M.L.; Leu, N.A.; Rao, S.; Li, F.; Gregory, B.D.; Zalenskaya, I.A.; Schultz, R.M.; Meyer, R.G. Paternal Poly (ADP-Ribose) Metabolism Modulates Retention of Inheritable Sperm Histones and Early Embryonic Gene Expression. PLoS Genet. 2014, 10, e1004317. [Google Scholar] [CrossRef]
- Castillo, J.; Jodar, M.; Oliva, R. The Contribution of Human Sperm Proteins to the Development and Epigenome of the Preimplantation Embryo. Hum. Reprod. Update 2018, 24, 535–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teperek, M.; Simeone, A.; Gaggioli, V.; Miyamoto, K.; Allen, G.E.; Erkek, S.; Kwon, T.; Marcotte, E.M.; Zegerman, P.; Bradshaw, C.R.; et al. Sperm Is Epigenetically Programmed to Regulate Gene Transcription in Embryos. Genome Res. 2016, 26, 1034–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneshiro, K.R.; Rechtsteiner, A.; Strome, S. Sperm-Inherited H3K27me3 Impacts Offspring Transcription and Development in C. Elegans. Nat. Commun. 2019, 10, 1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, P.D.; Rando, O.J. Chromatin as a Potential Carrier of Heritable Information. Curr. Opin. Cell Biol. 2010, 22, 284–290. [Google Scholar] [CrossRef] [Green Version]
- De Geyter, C.; Calhaz-Jorge, C.; Kupka, M.S.; Wyns, C.; Mocanu, E.; Motrenko, T.; Scaravelli, G.; Smeenk, J.; Vidakovic, S.; Goossens, V.; et al. ART in Europe, 2014: Results Generated from European Registries by ESHRE: The European IVF-Monitoring Consortium (EIM) for the European Society of Human Reproduction and Embryology (ESHRE). Hum. Reprod. 2018, 33, 1586–1601. [Google Scholar] [CrossRef]
- de Mateo, S.; Ramos, L.; van der Vlag, J.; de Boer, P.; Oliva, R. Improvement in Chromatin Maturity of Human Spermatozoa Selected through Density Gradient Centrifugation. Int. J. 2011, 34, 256–267. [Google Scholar] [CrossRef]
- Castillo, J.; Simon, L.; de Mateo, S.; Lewis, S.; Oliva, R. Protamine/DNA Ratios and DNA Damage in Native and Density Gradient Centrifuged Sperm From Infertile Patients. J. Androl. 2011, 32, 324–332. [Google Scholar] [CrossRef] [Green Version]
- Chiamchanya, C.; Kaewnoonual, N.; Visutakul, P.; Manochantr, S.; Chaiya, J. Comparative Study of the Effects of Three Semen Preparation Media on Semen Analysis, DNA Damage and Protamine Deficiency, and the Correlation between DNA Integrity and Sperm Parameters. Asian J. Androl. 2010, 12, 271–277. [Google Scholar] [CrossRef]
- Muratori, M.; Tarozzi, N.; Carpentiero, F.; Danti, S.; Perrone, F.M.; Cambi, M.; Casini, A.; Azzari, C.; Boni, L.; Maggi, M.; et al. Sperm Selection with Density Gradient Centrifugation and Swim up: Effect on DNA Fragmentation in Viable Spermatozoa. Sci. Rep. 2019, 9, 7492. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; License: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2021.
- Jodar, M.; Kalko, S.; Castillo, J.; Ballescà, J.L.; Oliva, R. Differential RNAs in the Sperm Cells of Asthenozoospermic Patients. Hum. Reprod. 2012, 27, 1431–1438. [Google Scholar] [CrossRef]
- Cerilli, L.A.; Kuang, W.; Roger, D. A Practical Approach to Testicular Biopsy Interpretation for Male Infertility. Arch. Pathol. Lab. Med. 2010, 134, 1197–1204. [Google Scholar] [CrossRef]
- Moch, H.; Cubilla, A.L.; Humphrey, P.A.; Reuter, V.E.; Ulbright, T.M. The 2016 WHO Classification of Tumours of the Urinary System and Male Genital Organs—Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2016, 70, 93–105. [Google Scholar] [CrossRef]
- Salonia, A.; Bettocchi, C.; Boeri, L.; Capogrosso, P.; Carvalho, J.; Cilesiz, N.C.; Cocci, A.; Corona, G.; Dimitropoulos, K.; Gül, M.; et al. European Association of Urology Guidelines on Sexual and Reproductive Health—2021 Update: Male Sexual Dysfunction. Eur. Urol. 2021, 80, 333–357. [Google Scholar] [CrossRef]
- Leblanc, L.; Lagrange, F.; Lecoanet, P.; Marçon, B.; Eschwege, P.; Hubert, J. Testicular Microlithiasis and Testicular Tumor: A Review of the Literature. Basic Clin. Androl. 2018, 28, 8. [Google Scholar] [CrossRef]
- Muciaccia, B.; Boitani, C.; Berloco, B.P.; Nudo, F.; Spadetta, G.; Stefanini, M.; de Rooij, D.G.; Vicini, E. Novel Stage Classification of Human Spermatogenesis Based on Acrosome Development1. Biol. Reprod. 2013, 89, 60. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Bolte, S.; Cordelières, F.P. A Guided Tour into Subcellular Colocalization Analysis in Light Microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2020. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H-Scores (Mean ± SD) | ||||||||
|---|---|---|---|---|---|---|---|---|
| SC | SPG | SPC | rSPD (I–V) | rSPD (VI–VIII) | eSPD | cSPD | ||
| H4K5bu | ||||||||
| Infertile patients with normal spermatogenesis | N (n = 6) | 2.5 ± 6.1 | 33 ± 35.4# | 0.0 ± 0.0 | 30.8 ± 46.9 | 319.1 ± 128.0# | 478.4 ± 40.7# | 0.9 ± 2.1# |
| Infertile patients with altered spermatogenesis | SCOS (n = 3) | 8.9 ± 15.4 | - | - | - | - | - | - |
| HP (n = 2) | 0.0 ± 0.0 | 41.3 ± 58.4 | 0.0 ± 0.0 | 10.0 ± 14.14 | 358.9 ± 118.7 | 455.6 ± 110.0 | 0.0 ± 0.0 | |
| SA (n = 3) | 0.0 ± 0.0 | 12.8 ± 13.2 | 1.7 ± 3.3 | 192.3 | 460.0 ± 242.5 | 439.4 ± 27.4 | 225.0 | |
| Testicular cancer patients | SEM (n = 4) | 0.0 ± 0.0 | 51.2 ± 46.3 | 0.0 ± 0.0 | 0.0 ± 0.0 | 219.0 ± 22.9 | 330.1 ± 69.9 * | 0.0 ± 0.0 |
| TER (n = 3) | 18.7 ± 23.8 | 135.9 ± 64.3 * | 0.0 ± 0.0 | 106.2 ± 57.3 | 411.1 ± 120.6 | 480.2 ± 98.7 | 0.0 ± 0.0 | |
| H4K5ac | ||||||||
| Infertile patients with normal spermatogenesis | N (n = 6) | 101.1 ± 59.5 | 245.3 ± 59.8 | 46.8 ± 29.1# | 282.7 ± 66.2# | 415.8 ± 80.8# | 494.9 ± 59.4 | 0.9 ± 2.1# |
| Testicular cancer patients | SEM (n = 4) | 228.5 ± 74.8 * | 259.2 ± 108.2 | 149.4 ± 107.9 | 320.7 ± 88.4 | 396.5 ± 135.9 | 473.4 ± 102.8 | 5.7 ± 7.86 |
| TER (n = 3) | 282.5 ± 39.2 * | 304.3 ± 40.2 | 165.5 ± 60.4 * | 342.9 ± 51.5 | 447.2 ± 93.7 | 513.1 ± 81.7 | 0.0 ± 0.0 | |
| Antibodies | Company | Reference |
|---|---|---|
| Primary antibodies | ||
| Rabbit monoclonal anti-H4K5bu | PTM BIO LLC, Chicago, IL, USA | #PTM-313 |
| Mouse monoclonal anti-H4K5ac | PTM BIO LLC, Chicago, IL, USA | #PTM-163 |
| Secondary antibodies (IHC) | ||
| Goat Anti-Rabbit IgG Antibody (H + L), Biotinylated | Vector Laboratories, Burlingame, CA, USA | BA-1000 |
| Goat Anti-Mouse IgG Antibody (H + L), Biotinylated | Vector Laboratories, Burlingame, CA, USA | BA-9200 |
| Secondary antibodies (IF) | ||
| Goat anti-mouse IgG Alexa Fluor 488 | Invitrogen, Waltham, MA, USA | A28175 |
| Goat anti-rabbit IgG Alexa Fluor 633 | Invitrogen, Waltham, MA, USA | A21070 |
| Secondary antibodies (Odyssey® Fluorescent WB) | ||
| Anti-Mouse IgG (IRDye-700) | LI-COR, Inc., Lincoln, NE, USA | P/N 925-68070 |
| Anti-Rabbit IgG (IRDye-800) | LI-COR, Inc., Lincoln, NE, USA | P/N 925-32211 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Iglesia, A.; Jauregi, P.; Jodar, M.; Barrachina, F.; Ded, L.; Mallofré, C.; Rodríguez-Carunchio, L.; Corral, J.M.; Ballescà, J.L.; Komrskova, K.; et al. H4K5 Butyrylation Coexist with Acetylation during Human Spermiogenesis and Are Retained in the Mature Sperm Chromatin. Int. J. Mol. Sci. 2022, 23, 12398. https://doi.org/10.3390/ijms232012398
de la Iglesia A, Jauregi P, Jodar M, Barrachina F, Ded L, Mallofré C, Rodríguez-Carunchio L, Corral JM, Ballescà JL, Komrskova K, et al. H4K5 Butyrylation Coexist with Acetylation during Human Spermiogenesis and Are Retained in the Mature Sperm Chromatin. International Journal of Molecular Sciences. 2022; 23(20):12398. https://doi.org/10.3390/ijms232012398
Chicago/Turabian Stylede la Iglesia, Alberto, Paula Jauregi, Meritxell Jodar, Ferran Barrachina, Lukas Ded, Carme Mallofré, Leonardo Rodríguez-Carunchio, Juan Manuel Corral, Josep Lluís Ballescà, Katerina Komrskova, and et al. 2022. "H4K5 Butyrylation Coexist with Acetylation during Human Spermiogenesis and Are Retained in the Mature Sperm Chromatin" International Journal of Molecular Sciences 23, no. 20: 12398. https://doi.org/10.3390/ijms232012398