
Cancer-Associated Fibroblasts Influence the Biological Properties of Malignant Tumours via Paracrine Secretion and Exosome Production

 ,
,  , , ,
, , , 
Abstract
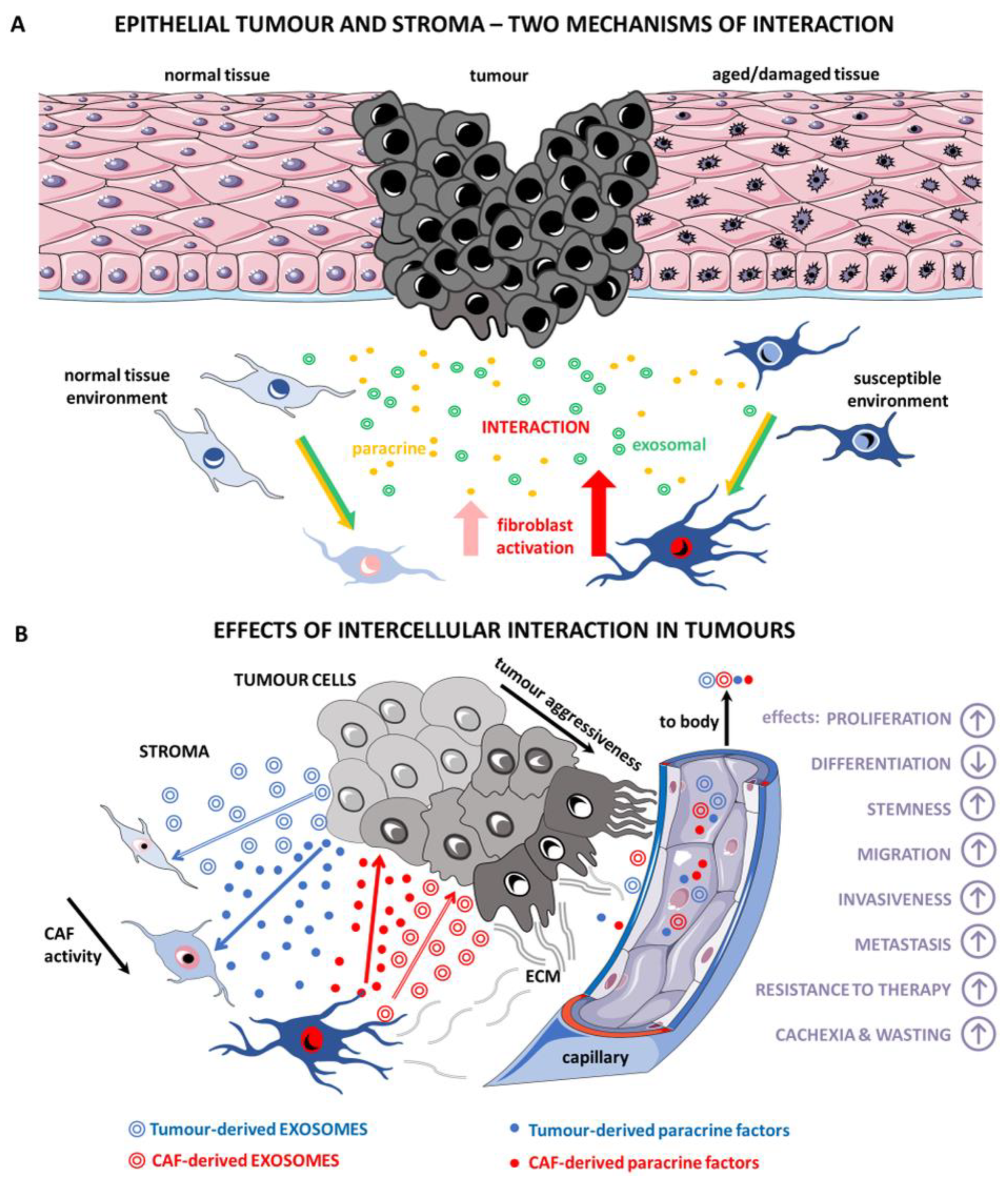
:1. Introduction
2. Fibroblasts Participate in Organ Morphogenesis
3. Fibroblasts as a Fundamental Bioactive Part of the Majority of Tumour Types
3.1. Origin of CAFs
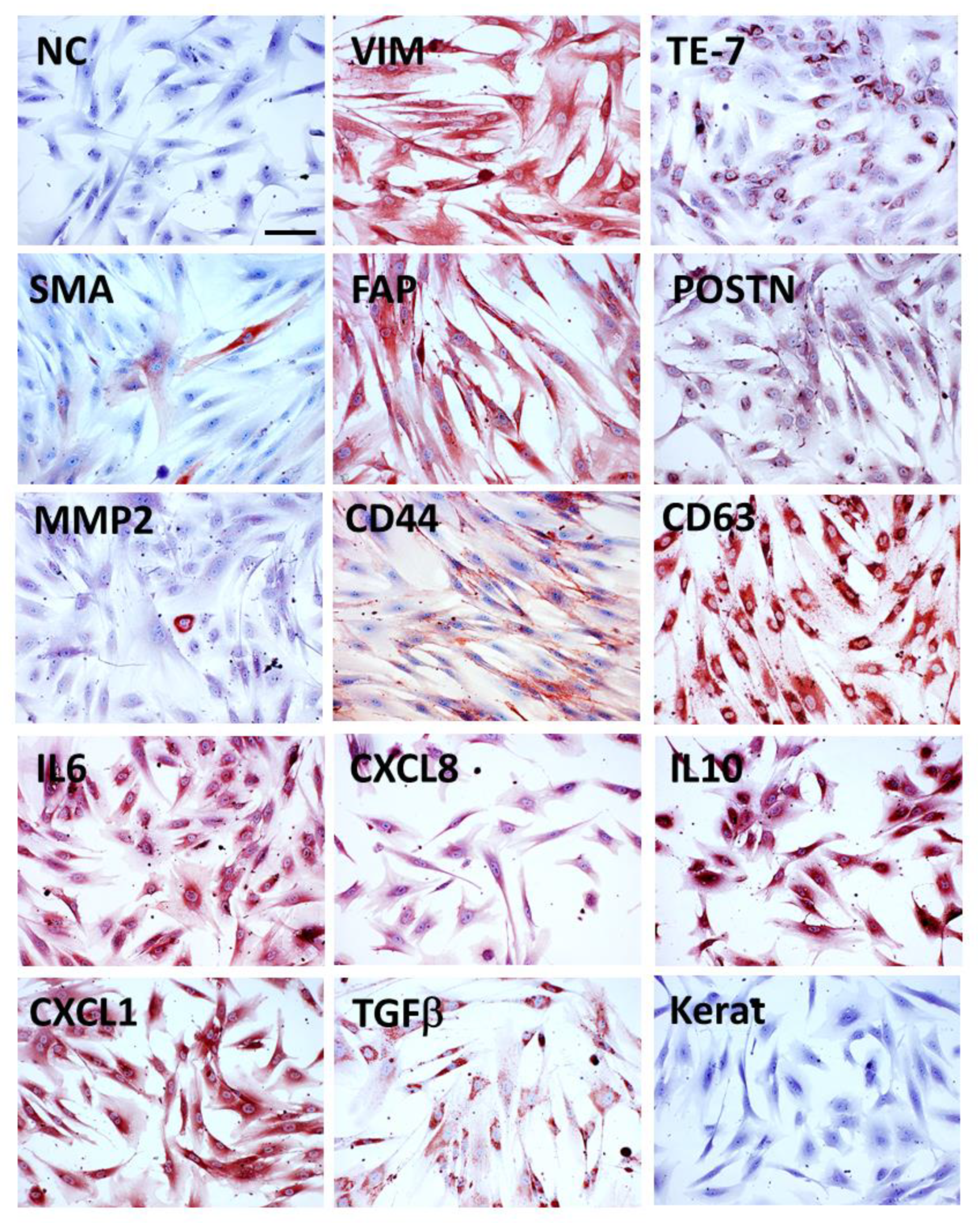
3.2. Markers of CAFs
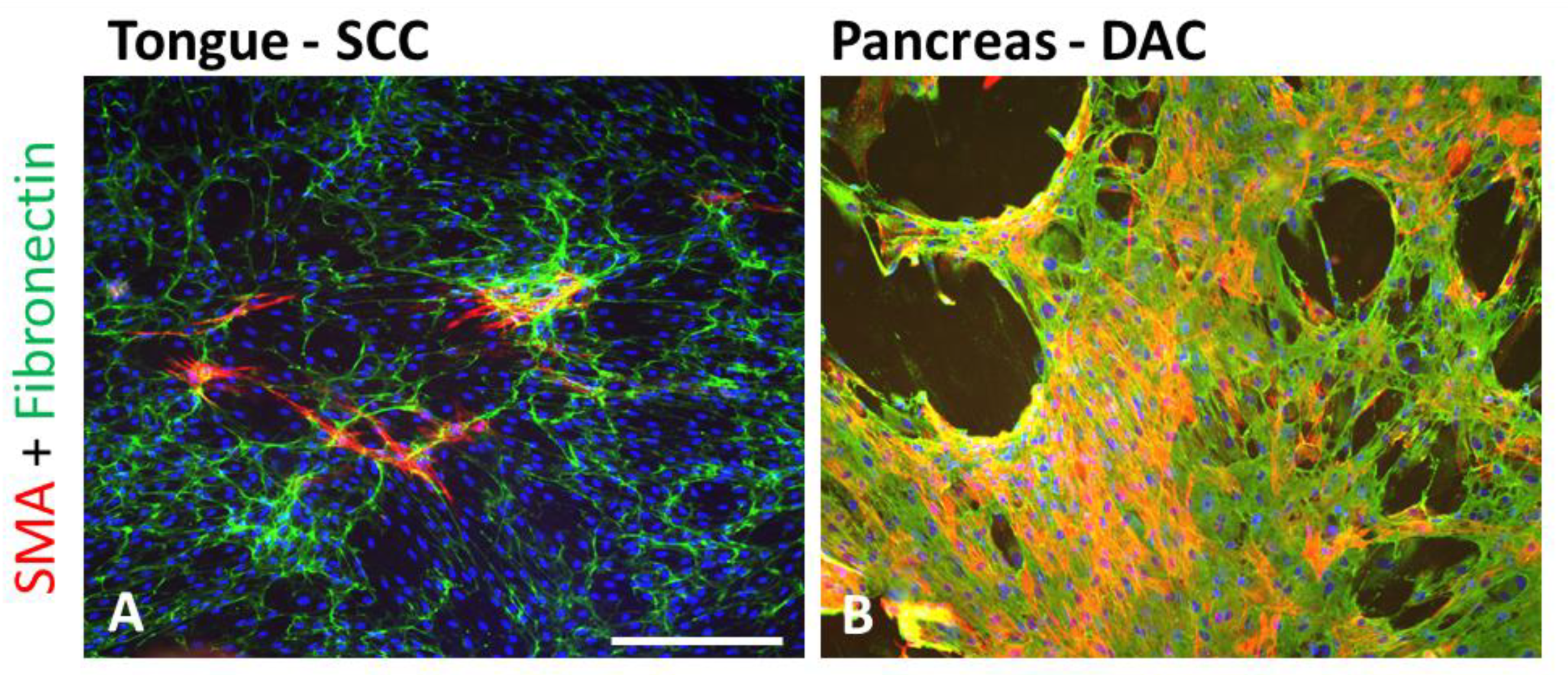
3.3. Heterogeneity of CAFs
3.4. Role of CAFs in Cancer Biology
3.4.1. Production and Remodelling of ECM
3.4.2. Production of Bioactive Factors with Emphasis on IL-6
4. Tumour Cell-Derived Exosomes in Cancer Biology
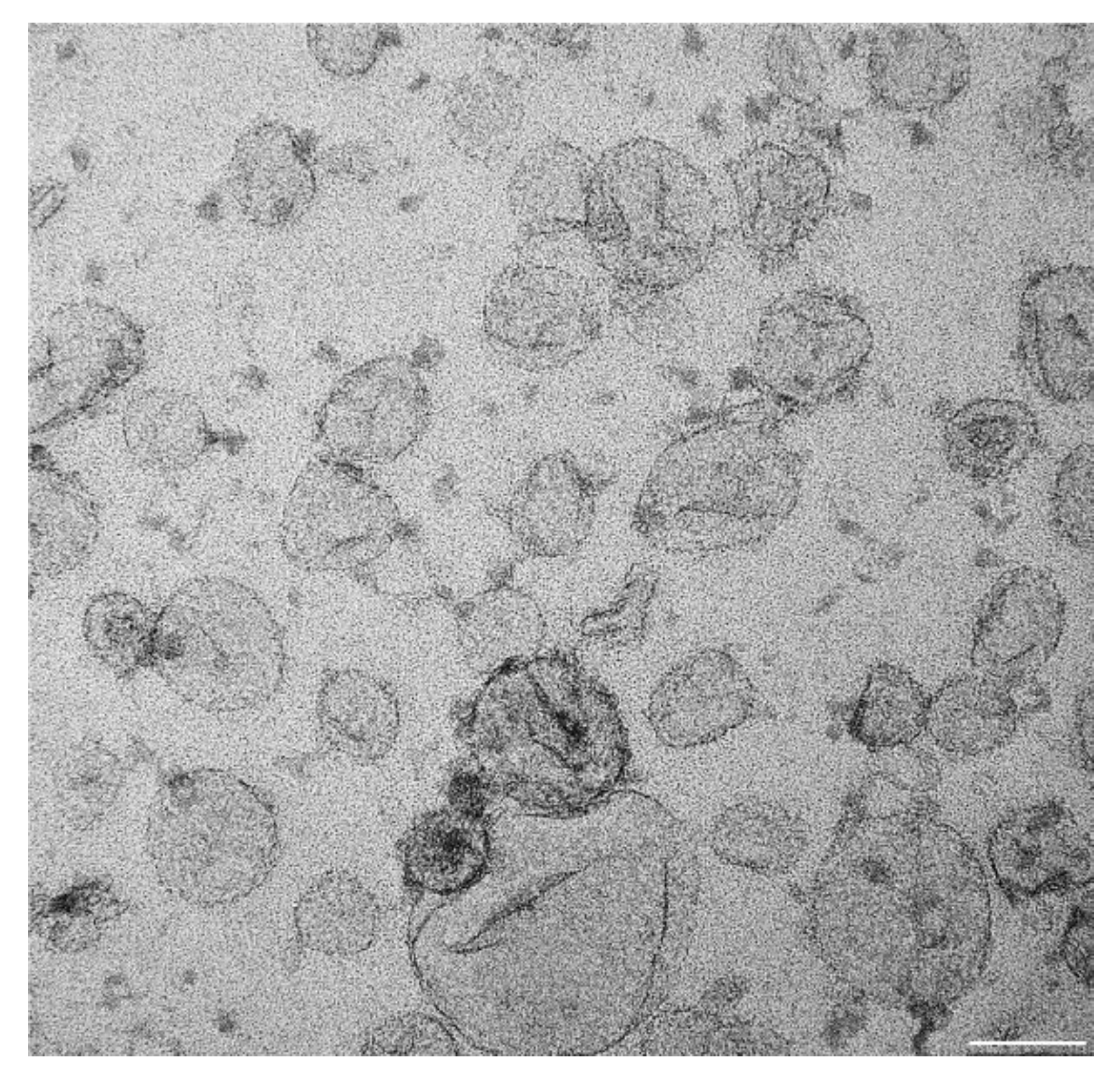
4.1. Definition of Exosome
4.2. Effect of Cancer Cell-Derived Exosomes (CaExos) on the Function of CAFs
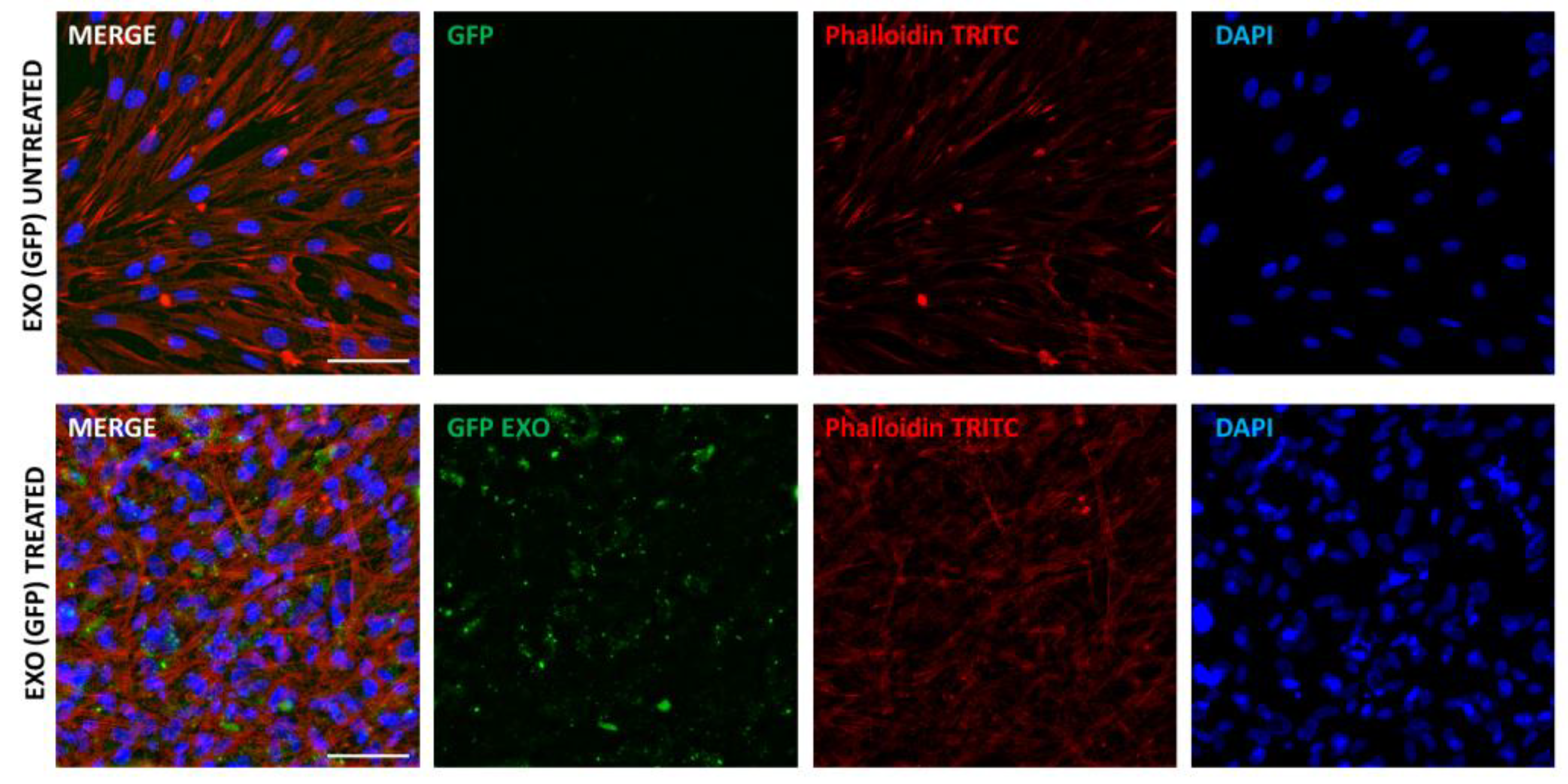
4.3. CAFs Produce Exosomes That Influence the Function of Cancer Cells
4.4. Systemic Effects of Exosomes—Role in Cancer Wasting and Cachexia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASC-1 | Neutral amino acid transporter SLC7A10 |
| CAFs | Cancer-associated fibroblasts |
| CaExos | Cancer cell-derived exosomes |
| CCL | Chemokine (C-C motif) ligand |
| CXCL | Chemokine (C-X-C motif) ligand |
| DDR2 | Discoidin domain receptor 2 |
| ECM | Extracellular matrix |
| EGF | Epidermal growth factor |
| EMT | Epithelial–mesenchymal transition |
| FAP | Fibroblast activation protein |
| FGF | Fibroblast growth factor |
| FSP1 | Fibroblast-specific protein 1 |
| HDGF | Hepatoma-derived growth factor |
| HGF | Hepatic growth factor |
| IGF | Insulin-like growth factor |
| IL | Interleukin |
| KERAT | Keratin |
| miR | microRNA |
| MMP | Matrix metalloproteinase |
| NG2 | Neural/glial antigen 2 |
| PDGF-BB | Platelet-derived growth factor BB |
| PDGFRα | Platelet-derived growth factor receptor α |
| PD-L1 | Programmed cell death ligand 1 |
| RAB3B | Ras-related protein Rab-3B |
| SMA | α-Smooth muscle actin |
| TGF | Transforming growth factor |
| TNFα | Tumour necrosis factor α |
| WNT | Wingless and Int signalization |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-Increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Nakasone, E.S.; Werb, Z. Tumors as Organs: Complex Tissues That Interface with the Entire Organism. Dev. Cell 2010, 18, 884–901. [Google Scholar] [CrossRef] [Green Version]
- Chia, S.B.; Degregori, J. Cancer Cells Build a Bad Neighbourhood in the Gut. Nature 2021, 594, 340–341. [Google Scholar] [CrossRef]
- Campisi, J. Senescent Cells, Tumor Suppression, and Organismal Aging: Good Citizens, Bad Neighbors. Cell 2005, 120, 513–522. [Google Scholar] [CrossRef]
- Krtolica, A.; Parrinello, S.; Lockett, S.; Desprez, P.Y.; Campisi, J. Senescent Fibroblasts Promote Epithelial Cell Growth and Tumorigenesis: A Link between Cancer and Aging. Proc. Natl. Acad. Sci. USA 2001, 98, 12072–12077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadiel, A.; Eichenbaum, K.D.; Xia, Y. Cell Interactome: Good Neighbors or Bad Neighbors. Biosci. Hypotheses 2008, 1, 255. [Google Scholar] [CrossRef] [Green Version]
- Mintz, B.; Illmensee, K. Normal Genetically Mosaic Mice Produced from Malignant Teratocarcinoma Cells. Proc. Natl. Acad. Sci. USA 1975, 72, 3585–3589. [Google Scholar] [CrossRef] [Green Version]
- Lacina, L.; Plzak, J.; Kodet, O.; Szabo, P.; Chovanec, M.; Dvorankova, B.; Smetana, K., Jr. Cancer Microenvironment: What Can We Learn from the Stem Cell Niche. Int. J. Mol. Sci. 2015, 16, 24094–24110. [Google Scholar] [CrossRef] [PubMed]
- Kodet, O.; Kučera, J.; Strnadová, K.; Dvořánková, B.; Štork, J.; Lacina, L.; Smetana, K. Cutaneous Melanoma Dissemination Is Dependent on the Malignant Cell Properties and Factors of Intercellular Crosstalk in the Cancer Microenvironment (Review). Int. J. Oncol. 2020, 57, 619–630. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef] [PubMed]
- Firestein, S.G.; Budd, R.C.; Gabriel, S.E.; McInnes, I.B.; O’Dell, J.R. Firestein: Kelley’s Textbook of Rheumatology, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Mikkola, M.L. Genetic Basis of Skin Appendage Development. Semin. Cell Dev. Biol. 2007, 18, 225–236. [Google Scholar] [CrossRef]
- Avagliano, A.; Fiume, G.; Ruocco, M.R.; Martucci, N.; Vecchio, E.; Insabato, L.; Russo, D.; Accurso, A.; Masone, S.; Montagnani, S.; et al. Influence of Fibroblasts on Mammary Gland Development, Breast Cancer Microenvironment Remodeling, and Cancer Cell Dissemination. Cancers 2020, 12, 1697. [Google Scholar] [CrossRef]
- Macias, H.; Hinck, L. Mammary Gland Development. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef] [Green Version]
- Schittny, J.C. Development of the Lung. Cell Tissue Res. 2017, 367, 427–444. [Google Scholar] [CrossRef] [Green Version]
- Duncan, S.A. Mechanisms Controlling Early Development of the Liver. Mech. Dev. 2003, 120, 19–33. [Google Scholar] [CrossRef]
- Jørgensen, M.C.; Ahnfelt-Rønne, J.; Hald, J.; Madsen, O.D.; Serup, P.; Hecksher-Sørensen, J. An Illustrated Review of Early Pancreas Development in the Mouse. Endocr. Rev. 2007, 28, 685–705. [Google Scholar] [CrossRef] [Green Version]
- Gittes, G.K. Developmental Biology of the Pancreas: A Comprehensive Review. Dev. Biol. 2009, 326, 4–35. [Google Scholar] [CrossRef] [Green Version]
- Driskell, R.R. Distinct Fibroblast Lineages Determine Dermal Architecture in Skin Development and Repair. Nature 2013, 504, 277–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driskell, R.R.; Watt, F.M. Understanding Fibroblast Heterogeneity in the Skin. Trends Cell Biol. 2015, 25, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Clavel, C.; Rendl, M.; Watt, F.M. Hair Follicle Dermal Papilla Cells at a Glance. J. Cell Sci. 2011, 124, 1179–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haydont, V.; Neiveyans, V.; Perez, P.; Busson, É.; Lataillade, J.J.; Asselineau, D.; Fortunel, N.O. Fibroblasts from the Human Skin Dermo-Hypodermal Junction Are Distinct from Dermal Papillary and Reticular Fibroblasts and from Mesenchymal Stem Cells and Exhibit a Specific Molecular Profile Related to Extracellular Matrix Organization and Modeling. Cells 2020, 9, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippeos, C.; Telerman, S.B.; Oulès, B.; Pisco, A.O.; Shaw, T.J.; Elgueta, R.; Lombardi, G.; Driskell, R.R.; Soldin, M.; Lynch, M.D.; et al. Spatial and Single-Cell Transcriptional Profiling Identifies Functionally Distinct Human Dermal Fibroblast Subpopulations. J. Investig. Dermatol. 2018, 138, 811–825. [Google Scholar] [CrossRef] [Green Version]
- Novotný, J.; Strnadová, K.; Dvořánková, B.; Kocourková, Š.; Jakša, R.; Dundr, P.; Pačes, V.; Smetana, K.; Kolář, M.; Lacina, L. Single-Cell RNA Sequencing Unravels Heterogeneity of the Stromal Niche in Cutaneous Melanoma Heterogeneous Spheroids. Cancers 2020, 12, 3324. [Google Scholar] [CrossRef] [PubMed]
- Kareva, I. What Can Ecology Teach Us about Cancer? Transl. Oncol. 2011, 4, 266–270. [Google Scholar] [CrossRef] [Green Version]
- Hirata, E.; Girotti, M.R.; Viros, A.; Hooper, S.; Spencer-Dene, B.; Matsuda, M.; Larkin, J.; Marais, R.; Sahai, E. Intravital Imaging Reveals How BRAF Inhibition Generates Drug-Tolerant Microenvironments with High Integrin Β1/FAK Signaling. Cancer Cell 2015, 27, 574–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beacham, E.C.D.A. Stromagenesis: The Changing Face of Fibroblastic Microenvironments during Tumor Progression. Semin. Cancer Biol. 2005, 15, 329–341. [Google Scholar] [CrossRef]
- Rodrigues, P.; Vanharanta, S. Circulating Tumor Cells: Come Together, Right Now, over Metastasis. Cancer Discov. 2019, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Paget, S. THE DISTRIBUTION OF SECONDARY GROWTHS IN CANCER OF THE BREAST. Lancet 1889, 133, 571–573. [Google Scholar] [CrossRef] [Green Version]
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of Deaths from Cancer Caused by Metastases? Cancer Med. 2019, 8, 5574. [Google Scholar] [CrossRef] [Green Version]
- Thompson, A.M.; Steel, C.M.; Chetty, U.; Carter, D.C. Evidence for the Multistep Theory of Carcinogenesis in Human Breast Cancer. Breast 1992, 1, 29–34. [Google Scholar] [CrossRef]
- Lacina, L.; Čoma, M.; Dvořánková, B.; Kodet, O.; Melegová, N.; Gál, P.; Smetana, K. Evolution of Cancer Progression in the Context of Darwinism. Anticancer. Res. 2019, 39, 1–16. [Google Scholar] [CrossRef]
- Lacina, L.; Kodet, O.; Dvořánková, B.; Szabo, P.; Smetana, K. Ecology of Melanoma Cell. Histol. Histopathol. 2018, 33, 247–254. [Google Scholar] [CrossRef]
- Josson, S.; Matsuoka, Y.; Chung, L.W.K.; Zhau, H.E.; Wang, R. Tumor-Stroma Co-Evolution in Prostate Cancer Progression and Metastasis. Semin. Cell Dev. Biol. 2010, 21, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujon, A.M.; Aktipis, A.; Alix-Panabières, C.; Amend, S.R.; Boddy, A.M.; Brown, J.S.; Capp, J.P.; DeGregori, J.; Ewald, P.; Gatenby, R.; et al. Identifying Key Questions in the Ecology and Evolution of Cancer. Evol. Appl. 2021, 14, 877–892. [Google Scholar] [CrossRef]
- Papkou, A.; Gokhale, C.S.; Traulsen, A.; Schulenburg, H. Host–Parasite Coevolution: Why Changing Population Size Matters. Zoology 2016, 119, 330–338. [Google Scholar] [CrossRef]
- Becht, E.; Giraldo, N.A.; Lacroix, L.; Buttard, B.; Elarouci, N.; Petitprez, F.; Selves, J.; Laurent-Puig, P.; Sautès-Fridman, C.; Fridman, W.H.; et al. Estimating the Population Abundance of Tissue-Infiltrating Immune and Stromal Cell Populations Using Gene Expression. Genome Biol. 2016, 17, 218. [Google Scholar] [CrossRef] [PubMed]
- Novák, Š.; Kolář, M.; Szabó, A.; Vernerová, Z.; Lacina, L.; Strnad, H.; Šáchová, J.; Hradilová, M.; Havránek, J.; Španko, M.; et al. Desmoplastic Crosstalk in Pancreatic Ductal Adenocarcinoma Is Reflected by Different Responses of Panc-1, MIAPaCa-2, PaTu-8902, and CAPAN-2 Cell Lines to Cancer-Associated/Normal Fibroblasts. Cancer Genom. Proteom. 2021, 18, 221–243. [Google Scholar] [CrossRef] [PubMed]
- Norton, J.; Foster, D.; Chinta, M.; Titan, A.; Longaker, M. Pancreatic Cancer Associated Fibroblasts (CAF): Under-Explored Target for Pancreatic Cancer Treatment. Cancers 2020, 12, 1347. [Google Scholar] [CrossRef]
- Balaziova, E.; Vymola, P.; Hrabal, P.; Mateu, R.; Zubal, M.; Tomas, R.; Netuka, D.; Kramar, F.; Zemanova, Z.; Svobodova, K.; et al. Fibroblast Activation Protein Expressing Mesenchymal Cells Promote Glioblastoma Angiogenesis. Cancers 2021, 13, 3304. [Google Scholar] [CrossRef]
- Pachva, M.C.; Lai, H.; Jia, A.; Rouleau, M.; Sorensen, P.H. Extracellular Vesicles in Reprogramming of the Ewing Sarcoma Tumor Microenvironment. Front. Cell Dev. Biol. 2021, 9, 726205. [Google Scholar] [CrossRef]
- Song, Y.J.; Xu, Y.; Deng, C.; Zhu, X.; Fu, J.; Chen, H.; Lu, J.; Xu, H.; Song, G.; Tang, Q.; et al. Gene Expression Classifier Reveals Prognostic Osteosarcoma Microenvironment Molecular Subtypes. Front. Immunol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Raffaghello, L.; Vacca, A.; Pistoia, V.; Ribatti, D. Cancer Associated Fibroblasts in Hematological Malignancies. Oncotarget 2015, 6, 2589–2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blentic, A.; Tandon, P.; Payton, S.; Walshe, J.; Carney, T.; Kelsh, R.N.; Mason, I.; Graham, A. The Emergence of Ectomesenchyme. Dev. Dyn. 2008, 237, 592–601. [Google Scholar] [CrossRef] [Green Version]
- LeBleu, V.S.; Neilson, E.G. Origin and Functional Heterogeneity of Fibroblasts. FASEB J. 2020, 34, 3519–3536. [Google Scholar] [CrossRef] [Green Version]
- Živicová, V.; Lacina, L.; Mateu, R.; Smetana, K.; Kavková, R.; Krejcí, E.D.; Grim, M.; Kvasilová, A.; Borský, J.; Strnad, H.; et al. Analysis of Dermal Fibroblasts Isolated from Neonatal and Child Cleft Lip and Adult Skin: Developmental Implications on Reconstructive Surgery. Int. J. Mol. Med. 2017, 40, 1323–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Yang, J.; Liu, J.; Wang, Y.; Mu, J.; Zeng, Q.; Deng, S.; Zhou, H. Signaling Pathways in Cancer-Associated Fibroblasts and Targeted Therapy for Cancer. Signal Transduct. Target. Ther. 2021, 6, 218. [Google Scholar] [CrossRef] [PubMed]
- Bukkuri, A.; Adler, F.R. Viewing Cancer through the Lens of Corruption: Using Behavioral Ecology to Understand Cancer. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Lee, Y.T.; Tan, Y.J.; Falasca, M.; Oon, C.E. Cancer-Associated Fibroblasts: Epigenetic Regulation and Therapeutic Intervention in Breast Cancer. Cancers 2020, 12, 2949. [Google Scholar] [CrossRef] [PubMed]
- Lacina, L.; Brábek, J.; Král, V.; Kodet, O.; Smetana, K. Interleukin-6: A Molecule with Complex Biological Impact in Cancer. Histol. Histopathol. 2019, 34, 125–136. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Petersen, O.W.; Nielsen, H.L.; Gudjonsson, T.; Villadsen, R.; Rank, F.; Niebuhr, E.; Bissell, M.J.; Rønnov-Jessen, L. Epithelial to Mesenchymal Transition in Human Breast Cancer Can Provide a Nonmalignant Stroma. Am. J. Pathol. 2003, 162, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Kopantzev, E.P.; Vayshlya, N.A.; Kopantseva, M.R.; Egorov, V.I.; Pikunov, M.; Zinovyeva, M.V.; Vinogradova, T.V.; Zborovskaya, I.B.; Sverdlov, E.D. Cellular and Molecular Phenotypes of Proliferating Stromal Cells from Human Carcinomas. Br. J. Cancer 2010, 102, 1533–1540. [Google Scholar] [CrossRef]
- Polyak, K.; Haviv, I.; Campbell, I.G. Co-Evolution of Tumor Cells and Their Microenvironment. Trends Genet. 2009, 25, 30–38. [Google Scholar] [CrossRef]
- Haviv, I.; Polyak, K.; Qiu, W.; Hu, M.; Campbell, I. Origin of Carcinoma Associated Fibroblasts. Cell Cycle (Georget. Tex.) 2009, 8, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Kodet, O.; Dvořánková, B.; Bendlová, B.; Sýkorová, V.; Krajsová, I.; Štork, J.; Kučera, J.; Szabo, P.; Strnad, H.; Kolář, M.; et al. Microenvironment-Driven Resistance to B-Raf Inhibition in a Melanoma Patient Is Accompanied by Broad Changes of Gene Methylation and Expression in Distal Fibroblasts. Int. J. Mol. Med. 2018, 41, 2687–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dvořánková, B.; Smetana, K.; Říhová, B.; Kučera, J.; Mateu, R.; Szabo, P. Cancer-Associated Fibroblasts Are Not Formed from Cancer Cells by Epithelial-to-Mesenchymal Transition in Nu/Nu Mice. Histochem. Cell Biol. 2015, 143, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Gunaydin, G.; Kesikli, S.A.; Guc, D. Cancer Associated Fibroblasts Have Phenotypic and Functional Characteristics Similar to the Fibrocytes That Represent a Novel MDSC Subset. Oncoimmunology 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, L.; Muñoz-Durango, N.; Riedel, C.A.; Echeverría, C.; Kalergis, A.M.; Cabello-Verrugio, C.; Simon, F. Endothelial-to-Mesenchymal Transition: Cytokine-Mediated Pathways That Determine Endothelial Fibrosis under Inflammatory Conditions. Cytokine Growth Factor Rev. 2017, 33, 41–54. [Google Scholar] [CrossRef]
- Ganguly, D.; Chandra, R.; Karalis, J.; Teke, M.; Aguilera, T.; Maddipati, R.; Wachsmann, M.B.; Ghersi, D.; Siravegna, G.; Zeh, H.J.; et al. Cancer-Associated Fibroblasts: Versatile Players in the Tumor Microenvironment. Cancers 2020, 12, 2652. [Google Scholar] [CrossRef]
- Iyoshi, S.; Yoshihara, M.; Nakamura, K.; Sugiyama, M.; Koya, Y.; Kitami, K.; Uno, K.; Mogi, K.; Tano, S.; Tomita, H.; et al. Pro-Tumoral Behavior of Omental Adipocyte-Derived Fibroblasts in Tumor Microenvironment at the Metastatic Site of Ovarian Cancer. Int. J. Cancer 2021, 149, 1961–1972. [Google Scholar] [CrossRef]
- Louault, K.; Li, R.R.; De Clerck, Y.A. Cancer-Associated Fibroblasts: Understanding Their Heterogeneity. Cancers 2020, 12, 3108. [Google Scholar] [CrossRef]
- Manoukian, P.; Bijlsma, M.; van Laarhoven, H. The Cellular Origins of Cancer-Associated Fibroblasts and Their Opposing Contributions to Pancreatic Cancer Growth. Front. Cell Dev. Biol. 2021, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Fang, Q.; Liu, P.; Ma, D.; Cao, S.; Zhang, L.; Chen, Q.; Hu, T.; Wang, J. Mesenchymal Stem Cells With Cancer-Associated Fibroblast-Like Phenotype Stimulate SDF-1/CXCR4 Axis to Enhance the Growth and Invasion of B-Cell Acute Lymphoblastic Leukemia Cells Through Cell-to-Cell Communication. Front. Cell Dev. Biol. 2021, 9, 9. [Google Scholar] [CrossRef]
- Dvořánková, B.; Szabo, P.; Lacina, L.; Gal, P.; Uhrova, J.; Zima, T.; Kaltner, H.; André, S.; Gabius, H.J.; Sykova, E.; et al. Human Galectins Induce Conversion of Dermal Fibroblasts into Myofibroblasts and Production of Extracellular Matrix: Potential Application in Tissue Engineering and Wound Repair. Cells Tissues Organs 2011, 194, 469–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Xia, Q.; Muhammad, T.; Liu, L.; Meng, X.; Bars-Cortina, D.; Khan, A.A.; Huang, Y.; Dong, L. Glioblastoma Therapy: Rationale for a Mesenchymal Stem Cell-Based Vehicle to Carry Recombinant Viruses. Stem Cell Rev. Rep. 2021, 1–21. [Google Scholar] [CrossRef]
- Moreno, R. Mesenchymal Stem Cells and Oncolytic Viruses: Joining Forces against Cancer. J. ImmunoTherapy Cancer 2021, 9, e001684. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.; Woods, E.L.; Dally, J.; Kong, D.; Steadman, R.; Moseley, R.; Midgley, A.C. Myofibroblasts: Function, Formation, and Scope of Molecular Therapies for Skin Fibrosis. Biomolecules 2021, 11, 1095. [Google Scholar] [CrossRef] [PubMed]
- Mifková, A.; Kodet, O.; Szabo, P.; Kučera, J.; Dvořánková, B.; André, S.; Koripelly, G.; Gabius, H.J.; Lehn, J.M.; Smetana, K. Synthetic Polyamine BPA-C8 Inhibits TGF-Β1-Mediated Conversion of Human Dermal Fibroblast to Myofibroblasts and Establishment of Galectin-1-Rich Extracellular Matrix in Vitro. ChemBioChem 2014, 15, 1465–1470. [Google Scholar] [CrossRef]
- Han, C.; Liu, T.; Yin, R. Biomarkers for Cancer-Associated Fibroblasts. Biomark. Res. 2020, 8, 64. [Google Scholar] [CrossRef]
- Boyd, L.N.C.; Andini, K.D.; Peters, G.J.; Kazemier, G.; Giovannetti, E. Heterogeneity and Plasticity of Cancer-Associated Fibroblasts in the Pancreatic Tumor Microenvironment. Semin. Cancer Biol. 2021, S1044-579X(21)00056-0. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Zhang, Q.; Liu, B.; Cheng, Y.; Zhang, Y.; Sun, Y.; Liu, J. Exosomal MiR-590-3p Derived from Cancer-Associated Fibroblasts Confers Radioresistance in Colorectal Cancer. Mol. Ther.-Nucleic Acids 2021, 24, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.S.; Kanugula, S.S.; Sudhir, S.; Pereira, M.P.; Jain, S.; Aghi, M.K. The Role of Cancer-Associated Fibroblasts in Tumor Progression. Cancers 2021, 13, 1399. [Google Scholar] [CrossRef] [PubMed]
- Busek, P.; Mateu, R.; Zubal, M.; Kotackova, L.; Sedo, A. Targeting Fibroblast Activation Protein in Cancer-Prospects and Caveats. Front. Biosci.-Landmark 2018, 23, 1933–1968. [Google Scholar]
- Šimková, A.; Bušek, P.; Šedo, A.; Konvalinka, J. Molecular Recognition of Fibroblast Activation Protein for Diagnostic and Therapeutic Applications. Biochim. Biophys. Acta-Proteins Proteom. 2020, 1868, 140409. [Google Scholar] [CrossRef] [PubMed]
- Irvine, A.F.; Waise, S.; Green, E.W.; Stuart, B.; Thomas, G.J. Characterising Cancer-Associated Fibroblast Heterogeneity in Non-Small Cell Lung Cancer: A Systematic Review and Meta-Analysis. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Miyashita, N.; Saito, A. Organ Specificity and Heterogeneity of Cancer-Associated Fibroblasts in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 10973. [Google Scholar] [CrossRef]
- Simon, T.; Salhia, B. Cancer Associated Fibroblast Subpopulations with Diverse and Dynamic Roles in the Tumor Microenvironment. Mol. Cancer Res. 2021. [Google Scholar] [CrossRef]
- Watt, D.M.; Morton, J.P. Heterogeneity in Pancreatic Cancer Fibroblasts—TGFβ as a Master Regulator? Cancers 2021, 13, 4984. [Google Scholar] [CrossRef]
- Pradhan, R.N.; Krishnamurty, A.T.; Fletcher, A.L.; Turley, S.J.; Müller, S. A Bird’s Eye View of Fibroblast Heterogeneity: A Pan-Disease, Pan-Cancer Perspective. Immunol. Rev. 2021, 302, 299–320. [Google Scholar] [CrossRef]
- Dvořánková, B.; Szabo, P.; Lacina, L.; Kodet, O.; Matouškové, E.; Smetana, K.; Matoušková, E.; Smetana, K. Fibroblasts Prepared from Different Types of Malignant Tumors Stimulate Expression of Luminal Marker Keratin 8 in the EM-G3 Breast Cancer Cell Line. Histochem. Cell Biol. 2012, 137, 679–685. [Google Scholar] [CrossRef]
- Geng, X.; Chen, H.; Zhao, L.; Hu, J.; Yang, W.; Li, G.; Cheng, C.; Zhao, Z.; Zhang, T.; Li, L.; et al. Cancer-Associated Fibroblast (CAF) Heterogeneity and Targeting Therapy of CAFs in Pancreatic Cancer. Front. Cell Dev. Biol. 2021, 9, 655152. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F. Tumors: Wounds That Do Not Heal. Cancer Immunol. Res. 2009, 315, 1650–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smetana, K.; Szabo, P.; Gál, P.; André, S.; Gabius, H.J.; Kodet, O.; Dvořánková, B. Emerging Role of Tissue Lectins as Microenvironmental Effectors in Tumors and Wounds. Histol. Histopathol. 2015, 30, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Szabó, P.; Kolář, M.; Dvořánková, B.; Lacina, L.; Štork, J.; Vlček, Č.; Strnad, H.; Tvrdek, M.; Smetana, K., Jr. Mouse 3T3 Fibroblasts under the Influence of Fibroblasts Isolated from Stroma of Human Basal Cell Carcinoma Acquire Properties of Multipotent Stem Cells. Biol. Cell 2011, 103, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Krejčí, E.; Dvořánková, B.; Szabo, P.; Naňka, O.; Strnad, H.; Kodet, O.; Lacina, L.; Kolář, M.; Smetana, K. Fibroblasts as Drivers of Healing and Cancer Progression: From In Vitro Experiments to Clinics; CRC Press: Boca Raton, FL, USA, 2016; ISBN 9781498704656. [Google Scholar]
- Lacina, L.; Smetana, K., Jr.; Dvořánková, B.; Pytlík, R.; Kideryová, L.; Kučerová, L.; Plzáková, Z.; Štork, J.; Gabius, H.-J.; André, S. Stromal Fibroblasts from Basal Cell Carcinoma Affect Phenotype of Normal Keratinocytes. Br. J. Dermatol. 2007, 156, 819–829. [Google Scholar] [CrossRef]
- Lacina, L.; Dvořánkova, B.; Smetana, K., Jr.; Chovanec, M.; Plzǎk, J.; Tachezy, R.; Kideryovǎ, L.; Kučerová, L.; Čada, Z.; Bouček, J.; et al. Marker Profiling of Normal Keratinocytes Identifies the Stroma from Squamous Cell Carcinoma of the Oral Cavity as a Modulatory Microenvironment in Co-Culture. Int. J. Radiat. Biol. 2007, 83, 837–848. [Google Scholar] [CrossRef]
- Ishii, T.; Suzuki, A.; Kuwata, T.; Hisamitsu, S.; Hashimoto, H.; Ohara, Y.; Yanagihara, K.; Mitsunaga, S.; Yoshino, T.; Kinoshita, T.; et al. Drug-Exposed Cancer-Associated Fibroblasts Facilitate Gastric Cancer Cell Progression Following Chemotherapy. Gastric Cancer 2021, 24, 810–822. [Google Scholar] [CrossRef] [PubMed]
- Kučera, J.; Strnadová, K.; Dvořánková, B.; Lacina, L.; Krajsová, I.; Štork, J.; Kovářová, H.; Skalníková, H.K.; Vodička, P.; Motlík, J.; et al. Serum Proteomic Analysis of Melanoma Patients with Immunohistochemical Profiling of Primary Melanomas and Cultured Cells: Pilot Study. Oncol. Rep. 2019, 42, 1793–1804. [Google Scholar] [CrossRef]
- Loumaye, A.; Thissen, J.P. Biomarkers of Cancer Cachexia. Clin. Biochem. 2017, 50, 1281–1288. [Google Scholar] [CrossRef]
- Cehreli, R.; Yavuzsen, T.; Ates, H.; Akman, T.; Ellidokuz, H.; Oztop, I. Can Inflammatory and Nutritional Serum Markers Predict Chemotherapy Outcomes and Survival in Advanced Stage Nonsmall Cell Lung Cancer Patients? BioMed Res. Int. 2019, 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zivicova, V.; Gal, P.; Mifkova, A.; Novak, S.; Kaltner, H.; Kolar, M.; Strnad, H.; Sachova, J.; Hradilova, M.; Chovanec, M.; et al. Detection of Distinct Changes in Gene-Expression Profiles in Specimens of Tumors and Transition Zones of Tenascin-Positive/-Negative Head and Neck Squamous Cell Carcinoma. Anticancer. Res. 2018, 38, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Libring, S.; Shinde, A.; Chanda, M.K.; Nuru, M.; George, H.; Saleh, A.M.; Abdullah, A.; Kinzer-Ursem, T.L.; Calve, S.; Wendt, M.K.; et al. The Dynamic Relationship of Breast Cancer Cells and Fibroblasts in Fibronectin Accumulation at Primary and Metastatic Tumor Sites. Cancers 2020, 12, 1270. [Google Scholar] [CrossRef]
- Sapudom, J.; Müller, C.D.; Nguyen, K.T.; Martin, S.; Anderegg, U.; Pompe, T. Matrix Remodeling and Hyaluronan Production by Myofibroblasts and Cancer-Associated Fibroblasts in 3d Collagen Matrices. Gels 2020, 6, 33. [Google Scholar] [CrossRef]
- Hashimoto, M.; Uesugi, N.; Osakabe, M.; Yanagawa, N.; Otsuka, K.; Kajiwara, Y.; Ueno, H.; Sasaki, A.; Sugai, T. Expression Patterns of Microenvironmental Factors and Tenascin-C at the Invasive Front of Stage II and III Colorectal Cancer: Novel Tumor Prognostic Markers. Front. Oncol. 2021, 11, 690816. [Google Scholar] [CrossRef] [PubMed]
- Kay, E.J.; Koulouras, G.; Zanivan, S. Regulation of Extracellular Matrix Production in Activated Fibroblasts: Roles of Amino Acid Metabolism in Collagen Synthesis. Front. Oncol. 2021, 11, 719922. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, V.; Chao, L.; Woodley, D.T.; Murrell, D.; Kim, M.; O’Toole, E.A.; Ly, A.; Cogan, J.; Mosallaei, D.; Wysong, A.; et al. Over-Expression of Stromal Periostin Correlates with Poor Prognosis of Cutaneous Squamous Cell Carcinomas. Exp. Dermatol. 2021, 30, 698–704. [Google Scholar] [CrossRef]
- Schwörer, S.; Pavlova, N.N.; Cimino, F.V.; King, B.; Cai, X.; Sizemore, G.M.; Thompson, C.B. Fibroblast Pyruvate Carboxylase Is Required for Collagen Production in the Tumour Microenvironment. Nat. Metab. 2021, 3, 1484–1499. [Google Scholar] [CrossRef] [PubMed]
- Sueyama, T.; Kajiwara, Y.; Mochizuki, S.; Shimazaki, H.; Shinto, E.; Hase, K.; Ueno, H. Periostin as a Key Molecule Defining Desmoplastic Environment in Colorectal Cancer. Virchows Arch. 2021, 478, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Eiro, N.; Cid, S.; Fraile, M.; Cabrera, J.R.; Gonzalez, L.O.; Vizoso, F.J. Analysis of the Gene Expression Profile of Stromal Pro-Tumor Factors in Cancer-Associated Fibroblasts from Luminal Breast Carcinomas. Diagnostics 2020, 10, 865. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Akasu, R.; Miyazaki, A. Calpain-Associated Proteolytic Regulation of the Stromal Microenvironment in Cancer. Curr. Pharm. Des. 2021, 27, 3128–3138. [Google Scholar] [CrossRef]
- Hassona, Y.; Cirillo, N.; Heesom, K.; Parkinson, E.K.; Prime, S.S. Senescent Cancer-Associated Fibroblasts Secrete Active MMP-2 That Promotes Keratinocyte Dis-Cohesion and Invasion. Br. J. Cancer 2014, 111, 1230–1237. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Chaudhuri, O. Beyond Proteases: Basement Membrane Mechanics and Cancer Invasion. J. Cell Biol. 2019, 218, 2456–2469. [Google Scholar] [CrossRef] [Green Version]
- Conti, S.; Kato, T.; Park, D.; Sahai, E.; Trepat, X.; Labernadie, A. CAFs and Cancer Cells Co-Migration in 3D Spheroid Invasion Assay. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2020; Volume 2179. [Google Scholar] [CrossRef]
- Miyazaki, K.; Togo, S.; Okamoto, R.; Idiris, A.; Kumagai, H.; Miyagi, Y. Collective Cancer Cell Invasion in Contact with Fibroblasts through Integrin-A5β1/Fibronectin Interaction in Collagen Matrix. Cancer Sci. 2020, 111, 4381–4392. [Google Scholar] [CrossRef]
- Mishra, P.; Banerjee, D.; Ben-Baruch, A. Chemokines at the Crossroads of Tumor-Fibroblast Interactions That Promote Malignancy. J. Leukoc. Biol. 2011, 89, 31–39. [Google Scholar] [CrossRef]
- Jobe, N.P.; Živicová, V.; Mifková, A.; Rösel, D.; Dvořánková, B.; Kodet, O.; Strnad, H.; Kolář, M.; Šedo, A.; Smetana, K.; et al. Fibroblasts Potentiate Melanoma Cells in Vitro Invasiveness Induced by UV-Irradiated Keratinocytes. Histochem. Cell Biol. 2018, 149, 503–516. [Google Scholar] [CrossRef]
- Asokan, S.; Bandapalli, O.R. CXCL8 Signaling in the Tumor Microenvironment. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 1302. [Google Scholar] [CrossRef]
- Španko, M.; Strnadová, K.; Pavlíček, A.J.; Szabo, P.; Kodet, O.; Valach, J.; Dvořánková, B.; Smetana, K.; Lacina, L. Il-6 in the Ecosystem of Head and Neck Cancer: Possible Therapeutic Perspectives. Int. J. Mol. Sci. 2021, 22, 1027. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brábek, J.; Jakubek, M.; Vellieux, F.; Novotný, J.; Kolář, M.; Lacina, L.; Szabo, P.; Strnadová, K.; Rösel, D.; Dvořánková, B.; et al. Interleukin-6: Molecule in the Intersection of Cancer, Ageing and COVID-19. Int. J. Mol. Sci. 2020, 21, 7937. [Google Scholar] [CrossRef]
- Shimura, Y.; Kurosawa, H.; Tsuchiya, M.; Sawa, M.; Kaneko, H.; Liu, L.; Makino, Y.; Nojiri, H.; Iwase, Y.; Kaneko, K.; et al. Serum Interleukin 6 Levels Are Associated with Depressive State of the Patients with Knee Osteoarthritis Irrespective of Disease Severity. Clin. Rheumatol. 2017, 36, 2781–2787. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.P.; Desprez, P.Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strnadova, K.; Sandera, V.; Dvorankova, B.; Kodet, O.; Duskova, M.; Smetana, K.; Lacina, L. Skin Aging: The Dermal Perspective. Clin. Dermatol. 2019, 37. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Interleukin-6 Signalling in Health and Disease. F1000Research 2020, 9, 1013. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Il-6 Trans-Signaling via the Soluble IL-6 Receptor: Importance for the Proinflammatory Activities of IL-6. Int. J. Biol. Sci. 2012, 8, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, K.; Ito, T.; Ozaki, Y.; Amuro, H.; Tanaka, A.; Nishizawa, T.; Son, Y.; Inaba, M.; Nomura, S. Serum Interleukin 6 before and after Therapy with Tocilizumab Is a Principal Biomarker in Patients with Rheumatoid Arthritis. J. Rheumatol. 2013, 40, 1074–1081. [Google Scholar] [CrossRef]
- Plzák, J.; Bouček, J.; Bandúrová, V.; Kolář, M.; Hradilová, M.; Szabo, P.; Lacina, L.; Chovanec, M.; Smetana, K. The Head and Neck Squamous Cell Carcinoma Microenvironment as a Potential Target for Cancer Therapy. Cancers 2019, 11, 440. [Google Scholar] [CrossRef] [Green Version]
- Jobe, N.P.; Rösel, D.; Dvořánková, B.; Kodet, O.; Lacina, L.; Mateu, R.; Smetana, K.; Brábek, J. Simultaneous Blocking of IL-6 and IL-8 Is Sufficient to Fully Inhibit CAF-Induced Human Melanoma Cell Invasiveness. Histochem. Cell Biol. 2016, 146, 205–217. [Google Scholar] [CrossRef]
- Ham, I.H.; Lee, D.; Hur, H. Cancer-Associated Fibroblast-Induced Resistance to Chemotherapy and Radiotherapy in Gastrointestinal Cancers. Cancers 2021, 13, 1172. [Google Scholar] [CrossRef]
- Fu, H.; Yang, H.; Zhang, X.; Xu, W. The Emerging Roles of Exosomes in Tumor–Stroma Interaction. J. Cancer Res. Clin. Oncol. 2016, 142, 1897–1907. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Dudás, J.; Riechelmann, H.; Skvortsova, I.I. The Role of Exosomes in Cancer Metastasis. Semin. Cancer Biol. 2017, 44, 170–181. [Google Scholar] [CrossRef]
- Liu, J.; Ren, L.; Li, S.; Li, W.; Zheng, X.; Yang, Y.; Fu, W.; Yi, J.; Wang, J.; Du, G. The Biology, Function, and Applications of Exosomes in Cancer. Acta Pharm. Sin. B 2021, 11, 2783–2797. [Google Scholar] [CrossRef] [PubMed]
- Dror, S.; Sander, L.; Schwartz, H.; Sheinboim, D.; Barzilai, A.; Dishon, Y.; Apcher, S.; Golan, T.; Greenberger, S.; Barshack, I.; et al. Melanoma MiRNA Trafficking Controls Tumour Primary Niche Formation. Nat. Cell Biol. 2016, 18, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Weidle, U.H.; Birzele, F.; Kollmorgen, G.; Rüger, R. The Multiple Roles of Exosomes in Metastasis. Cancer Genom. Proteom. 2017, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Dean, D.C.; Hornicek, F.J.; Shi, H.; Duan, Z. Exosomes Promote Pre-Metastatic Niche Formation in Ovarian Cancer. Mol. Cancer 2019, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef]
- Strnadová, K.; Pfeiferová, L.; Přikryl, P.; Dvořánková, B.; Vlčák, E.; Frýdlová, J.; Vokurka, M.; Novotný, J.; Šáchová, J.; Hradilová, M.; et al. Exosomes Produced by Melanoma Cells Significantly Influence the Biological Properties of Normal and Cancer-Associated Fibroblasts. Histochem. Cell Biol. 2021, 1–20. [Google Scholar] [CrossRef]
- Lee, J.S.; Yoo, J.E.; Kim, H.; Rhee, H.; Koh, M.J.; Nahm, J.H.; Choi, J.S.; Lee, K.H.; Park, Y.N. Tumor Stroma with Senescence-Associated Secretory Phenotype in Steatohepatitic Hepatocellular Carcinoma. PLoS ONE 2017, 12, e0171922. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, T.; Koiwa, M.; Yonemura, A.; Miyake, K.; Kariya, R.; Kubota, S.; Yokomizo-Nakano, T.; Yasuda-Yoshihara, N.; Uchihara, T.; Itoyama, R.; et al. Inflammation-Driven Senescence-Associated Secretory Phenotype in Cancer-Associated Fibroblasts Enhances Peritoneal Dissemination. Cell Rep. 2021, 34, 108779. [Google Scholar] [CrossRef]
- Kabir, T.D.; Leigh, R.J.; Tasena, H.; Mellone, M.; Coletta, R.D.; Parkinson, E.K.; Prime, S.S.; Thomas, G.J.; Paterson, I.C.; Zhou, D.; et al. A MiR-335/COX-2/PTEN Axis Regulates the Secretory Phenotype of Senescent Cancer-Associated Fibroblasts. Aging 2016, 8, 1608–1635. [Google Scholar] [CrossRef] [Green Version]
- Gener Lahav, T.; Adler, O.; Zait, Y.; Shani, O.; Amer, M.; Doron, H.; Abramovitz, L.; Yofe, I.; Cohen, N.; Erez, N. Melanoma-Derived Extracellular Vesicles Instigate Proinflammatory Signaling in the Metastatic Microenvironment. Int. J. Cancer 2019, 145, 2521–2534. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, T.L. Exosome and Mesenchymal Stem Cell Cross-Talk in the Tumor Microenvironment. Semin. Immunol. 2018, 35, 69–79. [Google Scholar] [CrossRef]
- Zhou, X.; Yan, T.; Huang, C.; Xu, Z.; Wang, L.; Jiang, E.; Wang, H.; Chen, Y.; Liu, K.; Shao, Z.; et al. Melanoma Cell-Secreted Exosomal MiR-155-5p Induce Proangiogenic Switch of Cancer-Associated Fibroblasts via SOCS1/JAK2/STAT3 Signaling Pathway. J. Exp. Clin. Cancer Res. 2018, 37, 242. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Hu, J. Melanoma-Derived Exosomes Induce Reprogramming Fibroblasts into Cancer-Associated Fibroblasts via Gm26809 Delivery. Cell Cycle 2019, 18, 3085–3094. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Geng, Y. Exosomes Derived from Chronic Lymphocytic Leukaemia Cells Transfer MiR-146a to Induce the Transition of Mesenchymal Stromal Cells into Cancer-Associated Fibroblasts. J. Biochem. 2020, 168, 491–498. [Google Scholar] [CrossRef]
- Yang, S.S.; Ma, S.; Dou, H.; Liu, F.; Zhang, S.Y.; Jiang, C.; Xiao, M.; Huang, Y.X. Breast Cancer-Derived Exosomes Regulate Cell Invasion and Metastasis in Breast Cancer via MiR-146a to Activate Cancer Associated Fibroblasts in Tumor Microenvironment. Exp. Cell Res. 2020, 391, 111983. [Google Scholar] [CrossRef]
- Huang, Q.; Hsueh, C.Y.; Shen, Y.J.; Guo, Y.; Huang, J.M.; Zhang, Y.F.; Li, J.Y.; Gong, H.L.; Zhou, L. Small Extracellular Vesicle-Packaged TGFβ1 Promotes the Reprogramming of Normal Fibroblasts into Cancer-Associated Fibroblasts by Regulating Fibronectin in Head and Neck Squamous Cell Carcinoma. Cancer Lett. 2021, 517. [Google Scholar] [CrossRef]
- Shelton, M.; Anene, C.A.; Nsengimana, J.; Roberts, W.; Newton-Bishop, J.; Boyne, J.R. The Role of CAF Derived Exosomal MicroRNAs in the Tumour Microenvironment of Melanoma. Biochim. Biophys. Acta-Rev. Cancer 2021, 1875, 188456. [Google Scholar] [CrossRef]
- Shu, S.; Matsuzaki, J.; Want, M.Y.; Conway, A.; Benjamin-Davalos, S.; Allen, C.L.; Koroleva, M.; Battaglia, S.; Odunsi, A.; Minderman, H.; et al. An Immunosuppressive Effect of Melanoma-Derived Exosomes on NY-ESO-1 Antigen-Specific Human CD8+ T Cells Is Dependent on IL-10 and Independent of BRAFV600E Mutation in Melanoma Cell Lines. Immunol. Investig. 2020, 49, 744–757. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Y.; Chang, X.; Ba, X.; Hu, N.; Liu, Q.; Fang, L.; Wang, Z. Melanoma-Derived Exosomes Endow Fibroblasts with an Invasive Potential via Mir-21 Target Signaling Pathway. Cancer Manag. Res. 2020, 12, 12965–12974. [Google Scholar] [CrossRef]
- Yeon, J.H.; Jeong, H.E.; Seo, H.; Cho, S.; Kim, K.; Na, D.; Chung, S.; Park, J.; Choi, N.; Kang, J.Y. Cancer-Derived Exosomes Trigger Endothelial to Mesenchymal Transition Followed by the Induction of Cancer-Associated Fibroblasts. Acta Biomater. 2018, 76, 146–153. [Google Scholar] [CrossRef]
- Deep, G.; Panigrahia, G.K. Hypoxia-Induced Signaling Promotes Prostate Cancer Progression: Exosomes Role as Messenger of Hypoxic Response in Tumor Microenvironmen. Crit. Rev. Oncog. 2015, 20, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Boussadia, Z.; Lamberti, J.; Mattei, F.; Pizzi, E.; Puglisi, R.; Zanetti, C.; Pasquini, L.; Fratini, F.; Fantozzi, L.; Felicetti, F.; et al. Acidic Microenvironment Plays a Key Role in Human Melanoma Progression through a Sustained Exosome Mediated Transfer of Clinically Relevant Metastatic Molecules. J. Exp. Clin. Cancer Res. 2018, 37, 245. [Google Scholar] [CrossRef]
- Xi, L.; Peng, M.; Liu, S.; Liu, Y.; Wan, X.; Hou, Y.; Qin, Y.; Yang, L.; Chen, S.; Zeng, H.; et al. Hypoxia-Stimulated ATM Activation Regulates Autophagy-Associated Exosome Release from Cancer-Associated Fibroblasts to Promote Cancer Cell Invasion. J. Extracell. Vesicles 2021, 10, e12146. [Google Scholar] [CrossRef]
- Melnik, B.C. MiR-21: An Environmental Driver of Malignant Melanoma? J. Transl. Med. 2015, 13, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dou, D.; Ren, X.; Han, M.; Xu, X.; Ge, X.; Gu, Y.; Wang, X. Cancer-Associated Fibroblasts-Derived Exosomes Suppress Immune Cell Function in Breast Cancer via the MiR-92/PD-L1 Pathway. Front. Immunol. 2020, 11, 2026. [Google Scholar] [CrossRef]
- Guo, L.; Li, B.; Yang, J.; Shen, J.; Ji, J.; Miao, M. Fibroblast-Derived Exosomal MicroRNA-369 Potentiates Migration and Invasion of Lung Squamous Cell Carcinoma Cells via NF1-Mediated MAPK Signaling Pathway. Int. J. Mol. Med. 2020, 46, 595–608. [Google Scholar] [CrossRef]
- Shan, G.; Gu, J.; Zhou, D.; Li, L.; Cheng, W.; Wang, Y.; Tang, T.; Wang, X. Cancer-Associated Fibroblast-Secreted Exosomal MiR-423-5p Promotes Chemotherapy Resistance in Prostate Cancer by Targeting GREM2 through the TGF-β Signaling Pathway. Exp. Mol. Med. 2020, 52, 1809–1822. [Google Scholar] [CrossRef]
- Chen, P.Y.; Wei, W.F.; Wu, H.Z.; Fan, L.S.; Wang, W. Cancer-Associated Fibroblast Heterogeneity: A Factor That Cannot Be Ignored in Immune Microenvironment Remodeling. Front. Immunol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, K.; Lu, X.; Wang, Y.; Chen, J. Cancer-Associated Fibroblasts-Derived Exosomes Promote Lung Cancer Progression by OIP5-AS1/ MiR-142-5p/ PD-L1 Axis. Mol. Immunol. 2021, 140, 47–58. [Google Scholar] [CrossRef]
- Kunou, S.; Shimada, K.; Takai, M.; Sakamoto, A.; Aoki, T.; Hikita, T.; Kagaya, Y.; Iwamoto, E.; Sanada, M.; Shimada, S.; et al. Exosomes Secreted from Cancer-Associated Fibroblasts Elicit Anti-Pyrimidine Drug Resistance through Modulation of Its Transporter in Malignant Lymphoma. Oncogene 2021, 40, 3989–4003. [Google Scholar] [CrossRef]
- Luo, G.; Zhang, Y.; Wu, Z.; Zhang, L.; Liang, C.; Chen, X. Exosomal LINC00355 Derived from Cancer-Associated Fibroblasts Promotes Bladder Cancer Cell Resistance to Cisplatin by Regulating MiR-34b-5p/ABCB1 Axis. Acta Biochim. Biophys. Sin. 2021, 53, 558–566. [Google Scholar] [CrossRef]
- Yin, H.; Yu, S.; Xie, Y.; Dai, X.; Dong, M.; Sheng, C.; Hu, J. Cancer-Associated Fibroblasts-Derived Exosomes Upregulate MicroRNA-135b-5p to Promote Colorectal Cancer Cell Growth and Angiogenesis by Inhibiting Thioredoxin-Interacting Protein. Cell. Signal. 2021, 84, 110029. [Google Scholar] [CrossRef]
- Yugawa, K.; Yoshizumi, T.; Mano, Y.; Itoh, S.; Harada, N.; Ikegami, T.; Kohashi, K.; Oda, Y.; Mori, M. Cancer-Associated Fibroblasts Promote Hepatocellular Carcinoma Progression through Downregulation of Exosomal MiR-150-3p. Eur. J. Surg. Oncol. 2021, 47, 384–393. [Google Scholar] [CrossRef]
- Zhang, H.W.; Shi, Y.; Liu, J.-B.; Wang, H.M.; Wang, P.Y.; Wu, Z.J.; Li, L.; Gu, L.P.; Cao, P.S.; Wang, G.R.; et al. Cancer-Associated Fibroblast-Derived Exosomal MicroRNA-24-3p Enhances Colon Cancer Cell Resistance to MTX by down-Regulating CDX2/HEPH Axis. J. Cell. Mol. Med. 2021, 25, 3699–3713. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, P.; Li, H.X. CAFs-Derived Exosomal MiRNA-130a Confers Cisplatin Resistance of NSCLC Cells through PUM2-Dependent Packaging. Int. J. Nanomed. 2021, 16, 561–577. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Du, J.; Min, Z.; Ma, L.; Zhang, W.; Zhu, W.; Liu, Y. Carcinoma-Associated Fibroblasts Derived Exosomes Modulate Breast Cancer Cell Stemness through Exonic CircHIF1A by MiR-580-5p in Hypoxic Stress. Cell Death Discov. 2021, 7, 1–15. [Google Scholar] [CrossRef]
- White, J.P. IL-6, Cancer and Cachexia: Metabolic Dysfunction Creates the Perfect Storm. Transl. Cancer Res. 2017, 6, S280–S285. [Google Scholar] [CrossRef]
- Yamagata, A.S.; Freire, P.P. Are Cachexia-Associated Tumors TransmitTERS of ER Stress. Biochem. Soc. Trans. 2021, 49, 1841–1853. [Google Scholar] [CrossRef]
- Kottorou, A.; Dimitrakopoulos, F.I.; Tsezou, A. Non-Coding RNAs in Cancer-Associated Cachexia: Clinical Implications and Future Perspectives. Transl. Oncol. 2021, 14, 101101. [Google Scholar] [CrossRef]
- Miao, C.; Zhang, W.; Feng, L.; Gu, X.; Shen, Q.; Lu, S.; Fan, M.; Li, Y.; Guo, X.; Ma, Y.; et al. Cancer-Derived Exosome MiRNAs Induce Skeletal Muscle Wasting by Bcl-2-Mediated Apoptosis in Colon Cancer Cachexia. Mol. Ther.-Nucleic Acids 2021, 24, 923–938. [Google Scholar] [CrossRef] [PubMed]
- Di, W.; Zhang, W.; Zhu, B.; Li, X.; Tang, Q.; Zhou, Y. Colorectal Cancer Prompted Adipose Tissue Browning and Cancer Cachexia through Transferring Exosomal MiR-146b-5p. J. Cell. Physiol. 2021, 236, 5399–5410. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, T.; Terracina, K.P.; Raza, A.; Matsubara, H.; Takabe, K. Cancer Cachexia, Mechanism and Treatment. World J. Gastrointest. Oncol. 2015, 7, 17–29. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Factors Inducing CAF Formation from Precursors | Note |
|---|---|---|
| Resident fibroblasts | IL-1β, TGF-β, FGF, PDGF | |
| Stellate cells | SDF1, TGF-β, PDGF | |
| Mesenchymal stem cells | SDF1, TGF-β, HDGF, FGF | |
| Adipocytes | TGF-β | |
| Mesothelial cells | TGF-β | |
| Fibrocytes | SDF1, TGF-β, FGF, IL-12, IFNγ | Novel fibrocyte subset circulating in peripheral blood |
| Pericytes | SDF1, TGF-β, PDGF | |
| Epithelial cells | TGF-β | Epithelial–mesenchymal transition is necessary |
| Endothelial cells | IL-1β, TGF-β, PDGF, TNFα | Endothelial–mesenchymal transition is necessary |
| Origin | Marker |
|---|---|
| Resident local fibroblasts | SMA, type I collagen, CCL2, RAB3B, tenascin C, periostin, podoplanin, S100A4, CD74 |
| Mesenchymal stem cells | DDR2, FSP-1, CXCL12, vimentin, SMA, calponin-1, PDGFRα, periostin, CD90, podoplanin |
| Adipocytes | FSP-1, SMA, FAP, ASC-1 |
| Endothelial cells | CD31, FSP-1, SMA, TGF-β2 |
| Pericytes | PDGF-BB, NG2, FSP-1, SMA |
| Type of Cancer | Molecule |
|---|---|
| Melanoma | miR-21, miR-155-5p, miR-210, miR-211, miR-222, long noncoding RNA Gm26809 |
| Chronic lymphocytic leukaemia | miR-146a |
| Breast cancer | miR-146a |
| Head and neck squamous cell carcinoma | TGF-β1 |
| Type of Cancer | Molecule | Effect |
|---|---|---|
| Breast | miR-92 miR-500a-5p miR-580-5p | Deregulation of PD-L1 Cancer cell proliferation and migration Cancer cell stemness |
| Gastrointestinal tract | miR-24-3p, miR-92a-3p, miR-135b-5p, miR-522, miR-590-3p | Cancer cell proliferation, angiogenesis, resistance to chemotherapy |
| Liver | miR-150-3p | Hepatoma cell migration |
| Lung cancer | miR-130a, miR-142-5p, miR-369 | Deregulation of PD-L1, cell migration, resistance to cisplatin |
| Malignant lymphoma | miR-4717-5p | Resistance to anti-pyrimidine drugs |
| Urinary bladder cancer | Long intergenic non-protein coding LINC00355 | Resistance to cisplatin |
| Prostate | miR-423-5p | Resistance to chemotherapy |
| Melanoma | miR-21, miR-155-5p | TGF-β production, tumour progression, angiogenesis support |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vokurka, M.; Lacina, L.; Brábek, J.; Kolář, M.; Ng, Y.Z.; Smetana, K., Jr. Cancer-Associated Fibroblasts Influence the Biological Properties of Malignant Tumours via Paracrine Secretion and Exosome Production. Int. J. Mol. Sci. 2022, 23, 964. https://doi.org/10.3390/ijms23020964
Vokurka M, Lacina L, Brábek J, Kolář M, Ng YZ, Smetana K Jr. Cancer-Associated Fibroblasts Influence the Biological Properties of Malignant Tumours via Paracrine Secretion and Exosome Production. International Journal of Molecular Sciences. 2022; 23(2):964. https://doi.org/10.3390/ijms23020964
Chicago/Turabian StyleVokurka, Martin, Lukáš Lacina, Jan Brábek, Michal Kolář, Yi Zhen Ng, and Karel Smetana, Jr. 2022. "Cancer-Associated Fibroblasts Influence the Biological Properties of Malignant Tumours via Paracrine Secretion and Exosome Production" International Journal of Molecular Sciences 23, no. 2: 964. https://doi.org/10.3390/ijms23020964