
Size and Fluorescence Properties of Algal Photosynthetic Antenna Proteins Estimated by Microscopy

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
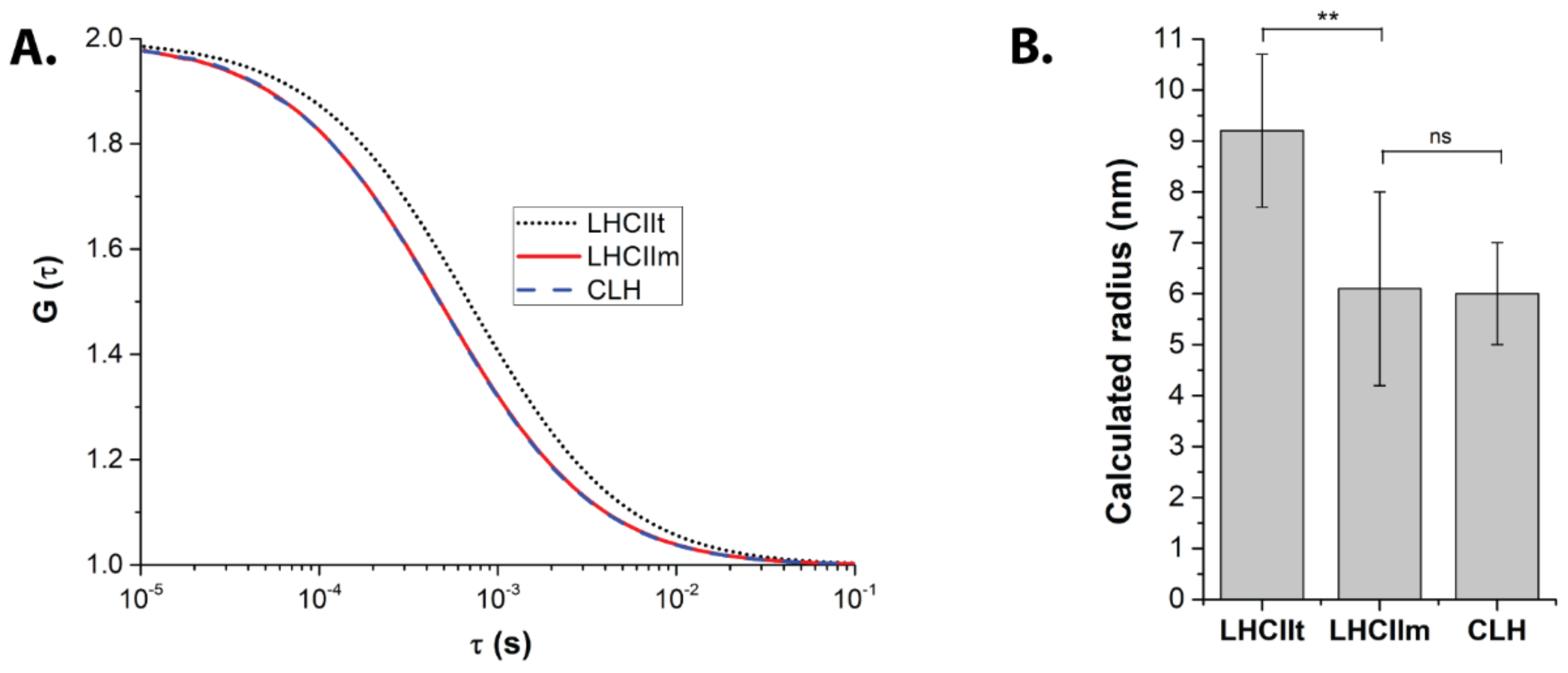
2.1. FCS Points to a Monomeric State of CLH
2.2. CLH and Monomeric LHC Proteins Co-Migrate on Sucrose Gradients
2.3. Fluorescence Yields of Antenna Proteins Varied with Their Oligomeric States
3. Discussion
4. Materials and Methods
4.1. Antenna Protein Purification and Preparation
4.2. Fluorescence Correlation Spectroscopy
4.3. Significance Level Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falkowski, P.G.; Owens, T.G. Light-Shade Adaptation: Two Strategies In Marine Phytoplankton. Plant. Physiol. 1980, 66, 592–595. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Andersson, B. The Dynamic Photosynthetic Membrane and Regulation of Solar-Energy Conversion. Trends Biochem. Sci. 1988, 13, 351–355. [Google Scholar] [CrossRef]
- Bonente, G.; Pippa, S.; Castellano, S.; Bassi, R.; Ballottari, M. Acclimation of Chlamydomonas reinhardtii to different growth irradiances. J. Biol. Chem. 2012, 287, 5833–5847. [Google Scholar] [CrossRef] [Green Version]
- Kouril, R.; Dekker, J.P.; Boekema, E.J. Supramolecular organization of photosystem II in green plants. Biochim. Biophys. Acta 2012, 1817, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotabova, E.; Jaresova, J.; Kana, R.; Sobotka, R.; Bina, D.; Prasil, O. Novel type of red-shifted chlorophyll a antenna complex from Chromera velia. I. Physiological relevance and functional connection to photosystems. Biochim. Biophys. Acta 2014, 1837, 734–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belgio, E.; Trskova, E.; Kotabova, E.; Ewe, D.; Prasil, O.; Kana, R. High light acclimation of Chromera velia points to photoprotective NPQ. Photosynth. Res. 2018, 135, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V. Light harvesting control in plants. FEBS Lett. 2018, 592, 3030–3039. [Google Scholar] [CrossRef]
- Pinnola, A. The rise and fall of Light-Harvesting Complex Stress-Related proteins as photoprotection agents during evolution. J. Exp. Bot. 2019, 70, 5527–5535. [Google Scholar] [CrossRef]
- Su, X.; Ma, J.; Wei, X.; Cao, P.; Zhu, D.; Chang, W.; Liu, Z.; Zhang, X.; Li, M. Structure and assembly mechanism of plant C2S2M2-type PSII-LHCII supercomplex. Science 2017, 357, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Nagao, R.; Kato, K.; Suzuki, T.; Ifuku, K.; Uchiyama, I.; Kashino, Y.; Dohmae, N.; Akimoto, S.; Shen, J.R.; Miyazaki, N.; et al. Structural basis for energy harvesting and dissipation in a diatom PSII-FCPII supercomplex. Nat. Plants 2019, 5, 890–901. [Google Scholar] [CrossRef]
- Pi, X.; Zhao, S.; Wang, W.; Liu, D.; Xu, C.; Han, G.; Kuang, T.; Sui, S.F.; Shen, J.R. The pigment-protein network of a diatom photosystem II-light-harvesting antenna supercomplex. Science 2019, 365. [Google Scholar] [CrossRef]
- Arshad, R.; Calvaruso, C.; Boekema, E.J.; Buchel, C.; Kouril, R. Revealing the architecture of the photosynthetic apparatus in the diatom Thalassiosira pseudonana. Plant. Physiol. 2021. [Google Scholar] [CrossRef]
- Wentworth, M.; Ruban, A.V.; Horton, P. The functional significance of the monomeric and trimeric states of the photosystem II light harvesting complexes. Biochemistry 2004, 43, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Janik, E.; Bednarska, J.; Zubik, M.; Sowinski, K.; Luchowski, R.; Grudzinski, W.; Gruszecki, W.I. Is It Beneficial for the Major Photosynthetic Antenna Complex of Plants To Form Trimers? J. Phys. Chem. B 2015, 119, 8501–8508. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Young, A.J.; Horton, P. Dynamic properties of the minor chlorophyll a/b binding proteins of photosystem II, an in vitro model for photoprotective energy dissipation in the photosynthetic membrane of green plants. Biochemistry 1996, 35, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Garab, G.; Cseh, Z.; Kovacs, L.; Rajagopal, S.; Varkonyi, Z.; Wentworth, M.; Mustardy, L.; Der, A.; Ruban, A.V.; Papp, E.; et al. Light-induced trimer to monomer transition in the main light-harvesting antenna complex of plants: Thermo-optic mechanism. Biochemistry 2002, 41, 15121–15129. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Bednarska, J.; Sowinski, K.; Luchowski, R.; Zubik, M.; Grudzinski, W.; Gruszecki, W.I. Light-induced formation of dimeric LHCII. Photosynth. Res. 2017, 132, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhoff, H.; Haase, W.; Wegner, S.; Danielsson, R.; Ackermann, R.; Albertsson, P.A. Low-light-induced formation of semicrystalline photosystem II arrays in higher plant chloroplasts. Biochemistry 2007, 46, 11169–11176. [Google Scholar] [CrossRef]
- Caffrey, M. Membrane protein crystallization. J. Struct. Biol. 2003, 142, 108–132. [Google Scholar] [CrossRef]
- Buchel, C. Fucoxanthin-chlorophyll proteins in diatoms: 18 and 19 kDa subunits assemble into different oligomeric states. Biochemistry 2003, 42, 13027–13034. [Google Scholar] [CrossRef]
- Bina, D.; Gardian, Z.; Herbstova, M.; Kotabova, E.; Konik, P.; Litvin, R.; Prasil, O.; Tichy, J.; Vacha, F. Novel type of red-shifted chlorophyll a antenna complex from Chromera velia: II. Biochemistry and spectroscopy. Biochim. Biophys. Acta 2014, 1837, 802–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Su, X.; Cao, P.; Liu, X.; Chang, W.; Li, M.; Zhang, X.; Liu, Z. Structure of spinach photosystem II-LHCII supercomplex at 3.2 A resolution. Nature 2016, 534, 69–74. [Google Scholar] [CrossRef]
- Van Bezouwen, L.S.; Caffarri, S.; Kale, R.S.; Kouril, R.; Thunnissen, A.W.H.; Oostergetel, G.T.; Boekema, E.J. Subunit and chlorophyll organization of the plant photosystem II supercomplex. Nat. Plants 2017, 3, 17080. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.; Tian, L.; Dai, H.E.; Qin, X.; Cheng, L.; Kuang, T.; Sui, S.F.; Shen, J.R. Unique organization of photosystem I-light-harvesting supercomplex revealed by cryo-EM from a red alga. Proc. Natl. Acad. Sci. USA 2018, 115, 4423–4428. [Google Scholar] [CrossRef] [Green Version]
- Burton-Smith, R.N.; Watanabe, A.; Tokutsu, R.; Song, C.; Murata, K.; Minagawa, J. Structural determination of the large photosystem II-light-harvesting complex II supercomplex of Chlamydomonas reinhardtii using nonionic amphipol. J. Biol. Chem. 2019, 294, 15003–15013. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Pi, X.; Wang, W.; Han, G.; Zhu, L.; Liu, M.; Cheng, L.; Shen, J.R.; Kuang, T.; Sui, S.F. Structure of a green algal photosystem I in complex with a large number of light-harvesting complex I subunits. Nat. Plants 2019, 5, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Huang, Z.; Chang, S.; Wang, W.; Wang, J.; Kuang, T.; Han, G.; Shen, J.R.; Zhang, X. Structure of a C2S2M2N2-type PSII-LHCII supercomplex from the green alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 21246–21255. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Watanabe, A.; Li, A.; Kim, E.; Song, C.; Murata, K.; Song, D.; Minagawa, J.; Liu, Z. Structural insight into light harvesting for photosystem II in green algae. Nat. Plants 2019, 5, 1320–1330. [Google Scholar] [CrossRef]
- Suga, M.; Ozawa, S.I.; Yoshida-Motomura, K.; Akita, F.; Miyazaki, N.; Takahashi, Y. Structure of the green algal photosystem I supercomplex with a decameric light-harvesting complex I. Nat. Plants 2019, 5, 626–636. [Google Scholar] [CrossRef]
- Wang, W.; Yu, L.J.; Xu, C.; Tomizaki, T.; Zhao, S.; Umena, Y.; Chen, X.; Qin, X.; Xin, Y.; Suga, M.; et al. Structural basis for blue-green light harvesting and energy dissipation in diatoms. Science 2019, 363. [Google Scholar] [CrossRef]
- Gelzinis, A.; Augulis, R.; Buchel, C.; Robert, B.; Valkunas, L. Confronting FCP structure with ultrafast spectroscopy data: Evidence for structural variations. Phys. Chem. Chem. Phys. PCCP 2021, 23, 806–821. [Google Scholar] [CrossRef] [PubMed]
- Tichy, J.; Gardian, Z.; Bina, D.; Konik, P.; Litvin, R.; Herbstova, M.; Pain, A.; Vacha, F. Light harvesting complexes of Chromera velia, photosynthetic relative of apicomplexan parasites. Biochim. Biophys. Acta 2013, 1827, 723–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llansola-Portoles, M.J.; Uragami, C.; Pascal, A.A.; Bina, D.; Litvin, R.; Robert, B. Pigment structure in the FCP-like light-harvesting complex from Chromera velia. Biochim. Biophys. Acta 2016, 1857, 1759–1765. [Google Scholar] [CrossRef] [PubMed]
- Crepin, A.; Cunill-Semanat, E.; Kuthanova Trskova, E.; Belgio, E.; Kana, R. Antenna Protein Clustering In Vitro Unveiled by Fluorescence Correlation Spectroscopy. Int. J. Mol. Sci. 2021, 22, 2969. [Google Scholar] [CrossRef]
- Kuthanova Trskova, E.; Belgio, E.; Yeates, A.M.; Sobotka, R.; Ruban, A.V.; Kana, R. Antenna proton sensitivity determines photosynthetic light harvesting strategy. J. Exp. Bot. 2018, 69, 4483–4493. [Google Scholar] [CrossRef] [PubMed]
- Caffarri, S.; Kouril, R.; Kereiche, S.; Boekema, E.J.; Croce, R. Functional architecture of higher plant photosystem II supercomplexes. EMBO J. 2009, 28, 3052–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crepin, A.; Santabarbara, S.; Caffarri, S. Biochemical and Spectroscopic Characterization of Highly Stable Photosystem II Supercomplexes from Arabidopsis. J. Biol. Chem. 2016, 291, 19157–19171. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.P.; Wientjes, E. The relevance of dynamic thylakoid organisation to photosynthetic regulation. Biochim. Biophys. Acta. Bioenerg. 2020, 1861, 148039. [Google Scholar] [CrossRef]
- Elson, E.L. Fluorescence correlation spectroscopy: Past, present, future. Biophys. J. 2011, 101, 2855–2870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, L.; Prado, M.; Arteni, A.A.; Elmlund, D.A.; Blankenship, R.E.; Adir, N. Structural studies show energy transfer within stabilized phycobilisomes independent of the mode of rod-core assembly. Biochim. Biophys. Acta 2014, 1837, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Iwai, M.; Pack, C.G.; Takenaka, Y.; Sako, Y.; Nakano, A. Photosystem II antenna phosphorylation-dependent protein diffusion determined by fluorescence correlation spectroscopy. Sci. Rep. 2013, 3, 2833. [Google Scholar] [CrossRef] [Green Version]
- Kana, R.; Steinbach, G.; Sobotka, R.; Vamosi, G.; Komenda, J. Fast Diffusion of the Unassembled PetC1-GFP Protein in the Cyanobacterial Thylakoid Membrane. Life 2020, 11, 15. [Google Scholar] [CrossRef]
- Lepetit, B.; Volke, D.; Gilbert, M.; Wilhelm, C.; Goss, R. Evidence for the existence of one antenna-associated, lipid-dissolved and two protein-bound pools of diadinoxanthin cycle pigments in diatoms. Plant. Physiol. 2010, 154, 1905–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarff, C.A.; Fuller, M.J.G.; Thompson, R.F.; Iadaza, M.G. Variations on Negative Stain Electron Microscopy Methods: Tools for Tackling Challenging Systems. J. Vis. Exp. 2018, 6, e57199. [Google Scholar] [CrossRef] [Green Version]
- Van Oort, B.; van Hoek, A.; Ruban, A.V.; van Amerongen, H. Aggregation of light-harvesting complex II leads to formation of efficient excitation energy traps in monomeric and trimeric complexes. FEBS Lett. 2007, 581, 3528–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregg, W.W.; Rousseaux, C.S. Directional and Spectral Irradiance in Ocean Models: Effects on Simulated Global Phytoplankton, Nutrients, and Primary Production. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Neilson, J.A.; Durnford, D.G. Structural and functional diversification of the light-harvesting complexes in photosynthetic eukaryotes. Photosynth Res. 2010, 106, 57–71. [Google Scholar] [CrossRef]
- Saccon, F.; Durchan, M.; Polivka, T.; Ruban, A.V. The robustness of the terminal emitter site in major LHCII complexes controls xanthophyll function during photoprotection. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2020, 19, 1308–1318. [Google Scholar] [CrossRef]
- Kana, R.; Kotabova, E.; Kopecna, J.; Trskova, E.; Belgio, E.; Sobotka, R.; Ruban, A.V. Violaxanthin inhibits nonphotochemical quenching in light-harvesting antenna of Chromera velia. FEBS Lett. 2016, 590, 1076–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levenberg, K. A method for the solution of certain non-linear problems in least squares. Quart. Appl. Math. 1944, 2, 164–168. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Sample | Diffusion Time (μs) | Particle Radius (nm) | Number of Protein Particles |
|---|---|---|---|
| LHCIIt | 745.6 ± 81.2 | 9.2 ± 1.5 | 25.1 ± 4.6 |
| LHCIIm | 490.9 ± 73.8 | 6.1 ± 1.9 | 38.3 ± 9.5 |
| CLH | 483.8 ± 81.7 | 6.0 ± 1.0 | 50.7 ± 11.7 |
| Sample | Total Photon Count Rate (kHz) | Count per Protein Particle (CPP) (Hz) | Nb Chl | Count per Chlorophyll (Hz) |
|---|---|---|---|---|
| LHCIIt | 7.44 ± 1.34 | 299.57 ± 42.66 | 42 | 7.13 ± 1.02 |
| LHCIIm | 7.53 ± 0.69 | 207.86 ± 56.56 | 14 | 14.85 ± 4.04 |
| CLH | 6.50 ± 1.40 | 135.43 ± 45.32 | 8 | 16.93 ± 5.66 |
| Complex | Approximate MW Apoprotein (Da) | Nb Chl | Nb Car | Approximate Total MW (Da) |
|---|---|---|---|---|
| CLH monomer | 18,500 | 8 | 4 | 28,100 |
| LHCII monomer | 25,000 | 14 | 4 | 40,000 |
| CLH dimer | 37,000 | 16 | 8 | 56,200 |
| CLH trimer | 55,500 | 24 | 12 | 84,300 |
| CLH tetramer | 74,000 | 32 | 16 | 112,400 |
| LHCII trimer | 75,000 | 42 | 12 | 120,000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crepin, A.; Belgio, E.; Šedivá, B.; Trsková, E.K.; Cunill-Semanat, E.; Kaňa, R. Size and Fluorescence Properties of Algal Photosynthetic Antenna Proteins Estimated by Microscopy. Int. J. Mol. Sci. 2022, 23, 778. https://doi.org/10.3390/ijms23020778
Crepin A, Belgio E, Šedivá B, Trsková EK, Cunill-Semanat E, Kaňa R. Size and Fluorescence Properties of Algal Photosynthetic Antenna Proteins Estimated by Microscopy. International Journal of Molecular Sciences. 2022; 23(2):778. https://doi.org/10.3390/ijms23020778
Chicago/Turabian StyleCrepin, Aurélie, Erica Belgio, Barbora Šedivá, Eliška Kuthanová Trsková, Edel Cunill-Semanat, and Radek Kaňa. 2022. "Size and Fluorescence Properties of Algal Photosynthetic Antenna Proteins Estimated by Microscopy" International Journal of Molecular Sciences 23, no. 2: 778. https://doi.org/10.3390/ijms23020778