
The Photosystem II Assembly Factor Ycf48 from the Cyanobacterium Synechocystis sp. PCC 6803 Is Lipidated Using an Atypical Lipobox Sequence

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
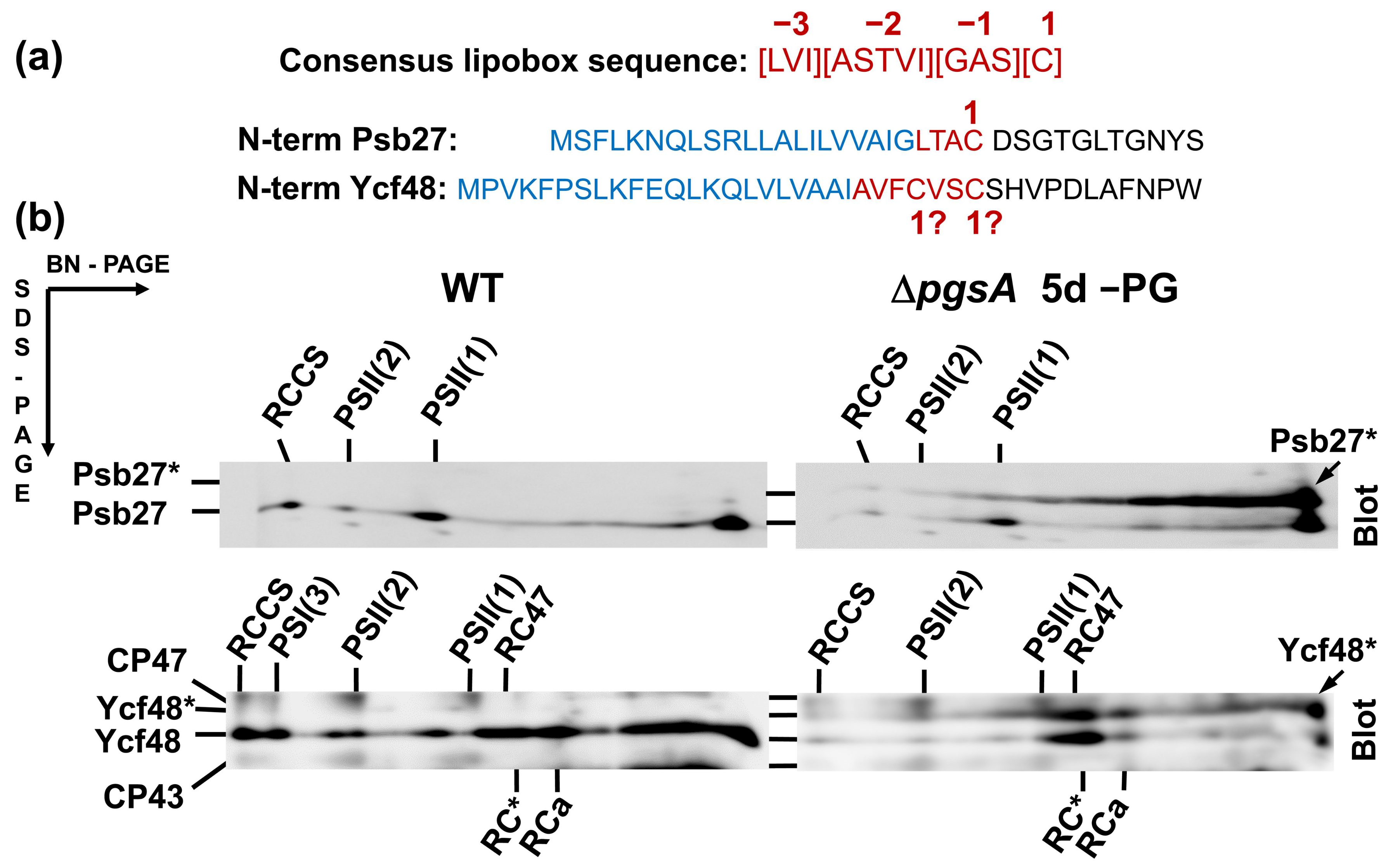
2.1. Analysis of Ycf48 in PG Deficient Cells
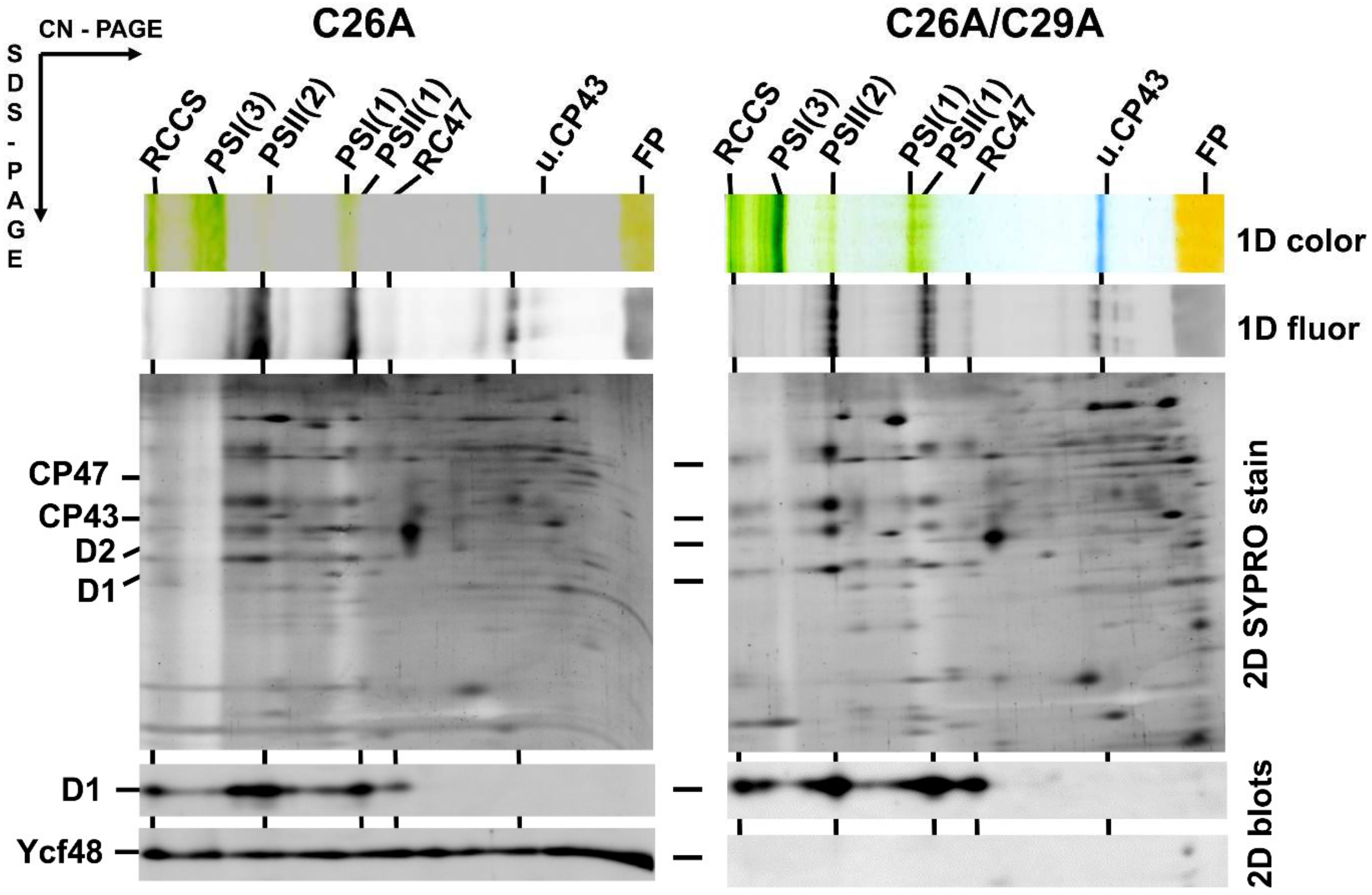
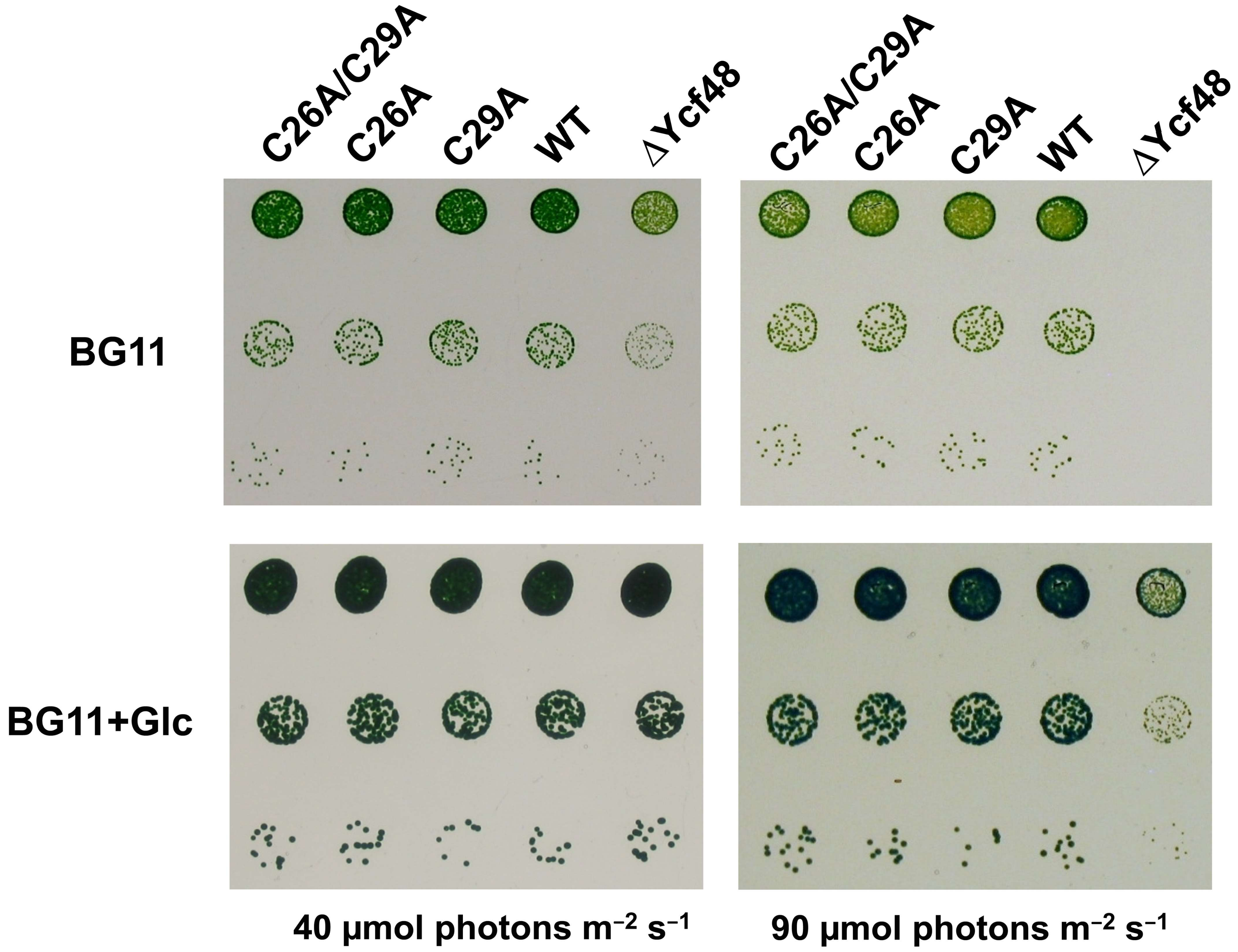
2.2. CVSC Motif in the Ycf48 Preprotein May Represent an Alternative Cyanobacterial Lipobox
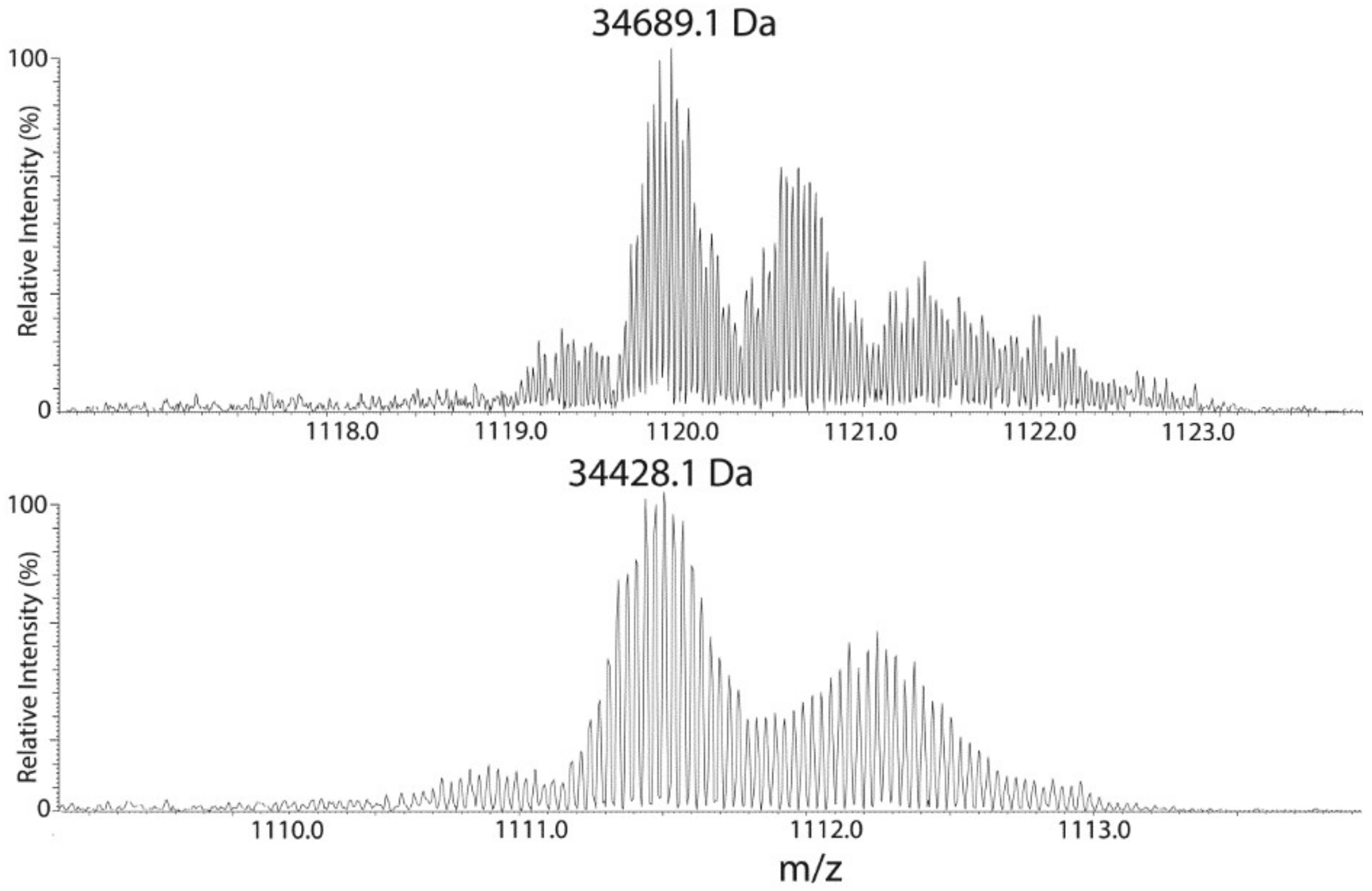
2.3. Mass Spectrometric Analysis Confirmed the Presence of a Lipidic Moiety at the N-Terminus of the Ycf48 Protein and Shortening of the C-Terminus
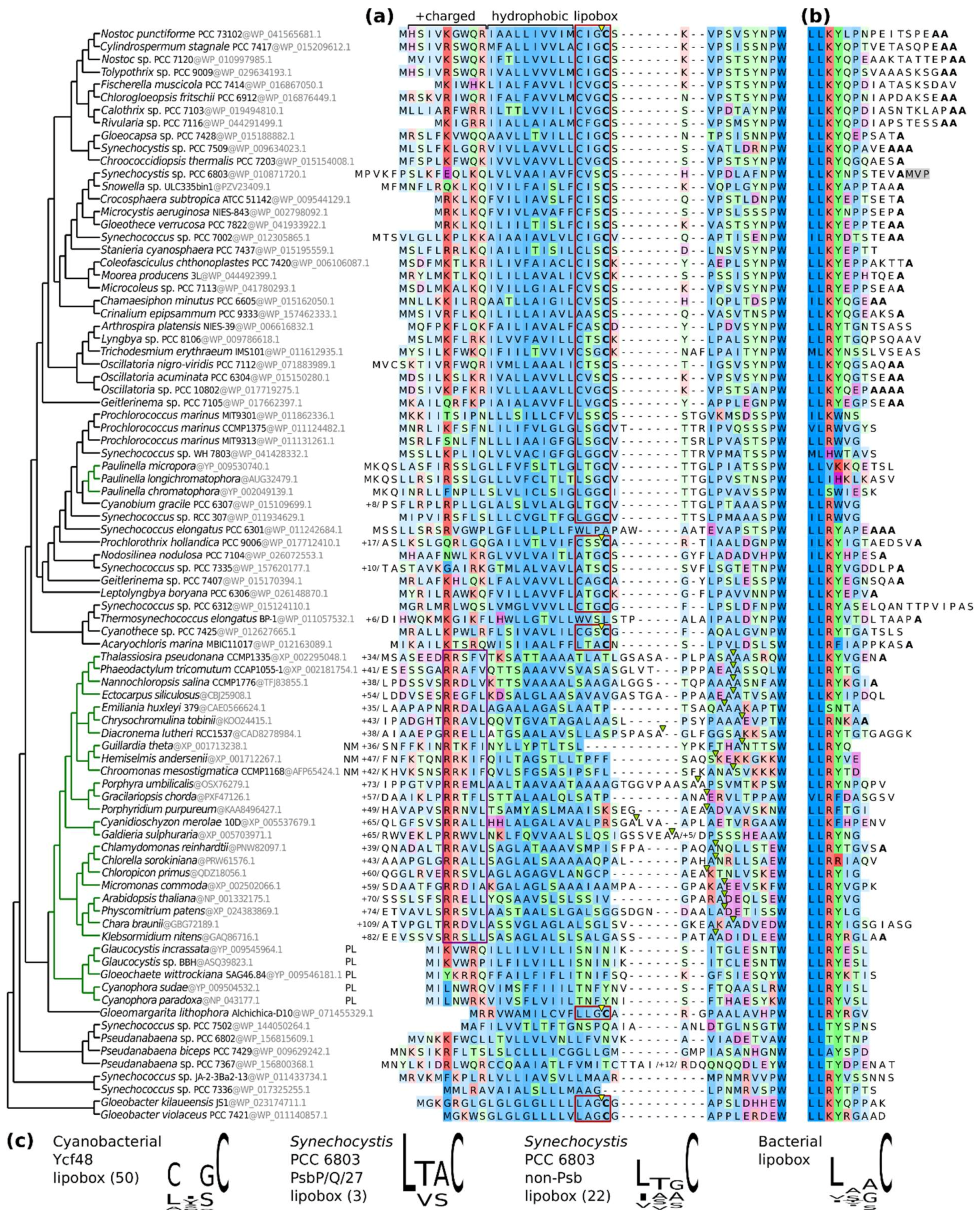
2.4. Comparing Lipoboxes and N- and C-Termini of Ycf48 among Cyanobacteria and Chloroplasts
3. Discussion
4. Materials and Methods
4.1. Synechocystis Strains, Their Construction and Growth Conditions
4.2. Determination of Chlorophyll Content
4.3. Preparation of Cellular Membranes and His-tag Purification
4.4. Protein Electrophoresis and Immunoblotting
4.5. Mass Spectrometry Analyses
4.6. In Silico Analysis of Ycf48 Sequences
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dühring, U.; Ossenbühl, F.; Wilde, A. Late assembly steps and dynamics of the cyanobacterial photosystem I. J. Biol. Chem. 2007, 282, 10915–10921. [Google Scholar] [CrossRef] [Green Version]
- Komenda, J.; Sobotka, R.; Nixon, P.J. Assembling and maintaining the photosystem II complex in chloroplasts and cyanobacteria. Cur. Opin. Plant. Biol. 2012, 15, 245–251. [Google Scholar] [CrossRef]
- Nickelsen, J.; Rengstl, B. Photosystem II assembly: From cyanobacteria to plants. Annu. Rev. Plant. Biol. 2013, 64, 609–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, S.; Liauw, P.; Nickelsen, J.; Nowaczyk, M. Analysis of photosystem II biogenesis in cyanobacteria. Biochim. Biophys. Acta 2016, 1857, 274–287. [Google Scholar] [CrossRef] [PubMed]
- Nowaczyk, M.M.; Hebeler, R.; Schlodder, E.; Meyer, H.E.; Warscheid, B.; Rogner, M. Psb27, a cyanobacterial lipoprotein, is involved in the repair cycle of photosystem II. Plant. Cell 2006, 18, 3121–3131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cormann, K.U.; Bartsch, M.; Rogner, M.; Nowaczyk, M.M. Localization of the CyanoP binding site on photosystem II by surface plasrnon resonance spectroscopy. Front. Plant. Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoppová, J.; Yu, J.F.; Konik, P.; Nixon, P.J.; Komenda, J. CyanoP is involved in the early steps of photosystem II assembly in the cyanobacterium Synechocystis sp PCC 6803. Plant. and Cell Physiology 2016, 57, 1921–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Summerfield, T.C.; Shand, J.A.; Bentley, F.K.; Eaton-Rye, J.J. PsbQ (Sll1638) in Synechocystis sp. PCC 6803 is required for Photosystem II activity in specific mutants and in nutrient-limiting conditions. Biochemistry 2005, 44, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Kashino, Y.; Inoue-Kashino, N.; Roose, J.L.; Pakrasi, H.B. Absence of the PsbQ protein results in destabilization of the PsbV protein and decreased oxygen evolution activity in cyanobacterial photosystem II. J. Biol. Chem. 2006, 281, 20834–20841. [Google Scholar] [CrossRef] [Green Version]
- Fagerlund, R.D.; Eaton-Rye, J.J. The lipoproteins of cyanobacterial photosystem II. J. Photochem. Photobiol. 2011, 104, 191–203. [Google Scholar] [CrossRef]
- Nakayama, H.; Kurokawa, K.; Lee, B.L. Lipoproteins in bacteria: Structures and biosynthetic pathways. FEBS J. 2012, 279, 4247–4268. [Google Scholar] [CrossRef] [PubMed]
- Buddelmeijer, N. The molecular mechanism of bacterial lipoprotein modification—How, when and why? FEMS Microbiol. Rev. 2015, 39, 246–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komenda, J.; Nickelsen, J.; Tichý, M.; Prášil, O.; Eichacker, L.A.; Nixon, P.J. The cyanobacterial homologue of HCF136/YCF48 is a component of an early photosystem II assembly complex and is important for both the efficient assembly and repair of photosystem II in Synechocystis sp PCC 6803. J. Biol. Chem. 2008, 283, 22390–22399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Knoppová, J.; Michoux, F.; Bialek, W.; Cota, E.; Shukla, M.K.; Strašková, A.; Pascual Aznar, G.; Sobotka, R.; Komenda, J.; et al. Ycf48 involved in the biogenesis of the oxygen-evolving photosystem II complex is a seven-bladed beta-propeller protein. Proc. Natl. Acad. Sci. U.S.A. 2018, 115, E7824–E7833. [Google Scholar] [CrossRef] [Green Version]
- Komenda, J.; Reisinger, V.; Muller, B.C.; Dobáková, M.; Granvogl, B.; Eichacker, L.A. Accumulation of the D2 protein is a key regulatory step for assembly of the photosystem II reaction center complex in Synechocystis PCC 6803. J. Biol. Chem. 2004, 279, 48620–48629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagio, M.; Gombos, Z.; Varkonyi, Z.; Masamoto, K.; Sato, N.; Tsuzuki, M.; Wada, H. Direct evidence for requirement of phosphatidylglycerol in photosystem II of photosynthesis. Plant. Physiol. 2000, 124, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Komenda, J.; Knoppová, J.; Kopečná, J.; Sobotka, R.; Halada, P.; Yu, J.F.; Nickelsen, J.; Boehm, M.; Nixon, P.J. The Psb27 assembly factor binds to the CP43 complex of Photosystem II in the cyanobacterium Synechocystis sp. PCC 6803. Plant. Physiol. 2012, 158, 476–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoppová, J.; Sobotka, R.; Tichý, M.; Yu, J.; Koník, P.; Halada, P.; Nixon, P.J.; Komenda, J. Discovery of a chlorophyll binding protein complex involved in the early steps of photosystem II assembly in Synechocystis. Plant. Cell 2014, 26, 1200–1212. [Google Scholar] [CrossRef] [Green Version]
- Frain, K.M.; Robinson, C.; van Dijl, J.M. Transport of folded proteins by the Tat system. Prot. J. 2019, 38, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Babu, M.M.; Priya, M.L.; Selvan, A.T.; Madera, M.; Gough, J.; Aravind, L.; Sankaran, K. A database of bacterial lipoproteins (DOLOP) with functional assignments to predicted lipoproteins. J. Bacteriol. 2006, 188, 2761–2773. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Xiao, Y.; Pi, X.; Zhao, L.; Zhu, Q.; Wang, W.; Kuang, T.; Han, G.; Sui, S.-F.; Shen, J.-R. Structural insights into a dimeric Psb27-photosystem II complex from a cyanobacterium Thermosynechococcus vulcanus. Proc. Natl. Acad. Sci. USA 2021, 118, e2018053118. [Google Scholar] [CrossRef]
- Mattoo, A.K.; Edelman, M. Intramembrane translocation and posttranslational palmitoylation of the chloroplast 32-kDa herbicide-binding protein. Proc. Natl. Acad. Sci. USA 1987, 84, 1497–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengstl, B.; Knoppová, J.; Komenda, J.; Nickelsen, J. Characterization of a Synechocystis double mutant lacking the photosystem II assembly factors YCF48 and Sll0933. Planta 2013, 237, 471–480. [Google Scholar] [CrossRef]
- Bučinská, L.; Kiss, E.; Konik, P.; Knoppová, J.; Komenda, J.; Sobotka, R. The ribosome-bound protein Pam68 promotes insertion of chlorophyll into the CP47 subunit of Photosystem II. Plant. Physiol. 2018, 176, 2931–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juneau, A.D.; Frankel, L.K.; Bricker, T.M.; Roose, J.L. N-Terminal Lipid Modification Is Required for the Stable Accumulation of CyanoQ in Synechocystis sp. PCC 6803. PLoS ONE 2016, 11, e0163646. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, H.; Weisz, D.A.; Vidavsky, I.; Gross, M.L.; Pakrasi, H.B. MS-based cross-linking analysis reveals the location of the PsbQ protein in cyanobacterial photosystem II. Proc. Natl. Acad. Sci. USA 2014, 111, 4638–4643. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.S.; Chisholm, D.A.; Dismukes, G.C.; Brudvig, G.W.; Diner, B.A. Spectroscopic evidence from site-directed mutants of Synechocystis PCC6803 in favor of a close interaction between histidine-189 and redox-active tyrosine-160 both of polypeptide D2 of the photosystem II reaction center. Biochemistry 1993, 32, 13742–13748. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yu, J.F.; Kobayashi, T.; Nakanishi, H.; Nixon, P.J.; Noguchi, T. Functional roles of D2-Lys317 and the interacting chloride ion in the water oxidation reaction of photosystem II as revealed by Fourier transform infrared analysis. Biochemistry 2013, 52, 4748–4757. [Google Scholar] [CrossRef] [Green Version]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Blue native electrophoresis for isolation of membrane protein complexes in enzymatically active form. Anal. Biochem. 1991, 199, 223–231. [Google Scholar] [CrossRef]
- Wittig, I.; Karas, M.; Schagger, H. High resolution clear native electrophoresis for in-gel functional assays and fluorescence studies of membrane protein complexes. Mol. Cell. Proteomics 2007, 6, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Dobáková, M.; Sobotka, R.; Tichý, M.; Komenda, J. Psb28 protein is involved in the biogenesis of the photosystem II inner antenna CP47 (PsbB) in the cyanobacterium Synechocystis sp. PCC 6803. Plant. Physiol. 2009, 149, 1076–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, E.; Knoppová, J.; Aznar, G.P.; Pilný, J.; Yu, J.; Halada, P.; Nixon, P.J.; Sobotka, R.; Komenda, J. A photosynthesis-specific rubredoxin-like protein is required for efficient association of the D1 and D2 proteins during the initial steps of photosystem II assembly. Plant. Cell 2019, 31, 2241–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitelegge, J.P.; Gundersen, C.B.; Faull, K.F. Electrospray-ionization mass spectrometry of intact intrinsic membrane proteins. Protein Sci. 1998, 7, 1423–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Marshall, A.G. A universal algorithm for fast and automated charge state deconvolution of electrospray mass-to-charge ratio spectra. J. Am. Soc. Mass Spectr. 1998, 9, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponce-Toledo, R.I.; Deschamps, P.; López-García, P.; Zivanovic, Y.; Benzerara, K.; Moreira, D. An early-branching freshwater cyanobacterium at the origin of plastids. Curr. Biol. 2017, 27, 386–391. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Retention Time (min) | Measured Mass (Average, Da) | Calculated Mass (Average, Da) | Assignment (From Init Met) | Delta (Da) |
|---|---|---|---|---|---|
| Intact Protein | |||||
| Intact protein | 51 | 34,689.1 | 33,874.6 | 29–339 | 814.5 |
| Intact protein | 43 | 34,428.1 | 33,874.6 | 29–339 | 553.5 |
| CNBr Fragments | |||||
| N-terminal fr | 59.9–60.2 | 14,117.0 | 13,303.6 | 29–151 | 813.3 |
| 58.4–58.6 | 22,188.5 | 21,356.6 + 16.0 | 29–224 | 815.8 | |
| C-terminal fr | 28.3 | 5525.3 | 5525.1 | 289–339 | 0.1 |
| 32.7 | 12,505.0 | 12,487.8 +16.0 | 225–339 | 1.1 | |
| Primers | Sequences (5′–3´) |
|---|---|
| Ycf48-KasI-F | ATGTCCGGTGTGTGGCGCC |
| Ycf48-HindIII-R | TACGGCCCCCTCCACCAAAGCTT |
| Ycf48C26A-3F | GCGGCGATCGCCGTTTTCGCGGTGAGCTGCAGCCATGTGCCGGA |
| Ycf48C26A-2R | TCCGGCACATGGCTGCAGCTCACCGCGAAAACGGCGATCGCCGC |
| Ycf48C29A-3F | TCGCCGTTTTCTGTGTGAGCGCGAGCCATGTGCCGGATTTGGCCT |
| Ycf48C29A-2R | AGGCCAAATCCGGCACATGGCTCGCGCTCACACAGAAAACGGCGA |
| Ycf48C26/29A-3F | GCGGCGATCGCCGTTTTCGCGGTGAGCGCGAGCCATGTGCCGGATTTGGCCT |
| Ycf48C26/29A-2R | AGGCCAAATCCGGCACATGGCTCGCGCTCACCGCGAAAACGGCGATCGCCGC |
| psbDC1F | AGTTGCGACAAAATAACCCAGCTCCAGCAA |
| psbD-His-2R | GCGCGTCCGACTGCAATAGTATGATGATGATGATGATGCATAAATGCAAATCCTCTTGCGTAGCT |
| psbD-His-3F | AGCTACGCAAGAGGATTTGCATTTATGCATCATCATCATCATCATACTATTGCAGTCGGACGCGC |
| psbDC4R | TTGCCAAAGTATTCTCCTGATTTAAATGATATTGAGCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knoppová, J.; Yu, J.; Janouškovec, J.; Halada, P.; Nixon, P.J.; Whitelegge, J.P.; Komenda, J. The Photosystem II Assembly Factor Ycf48 from the Cyanobacterium Synechocystis sp. PCC 6803 Is Lipidated Using an Atypical Lipobox Sequence. Int. J. Mol. Sci. 2021, 22, 3733. https://doi.org/10.3390/ijms22073733
Knoppová J, Yu J, Janouškovec J, Halada P, Nixon PJ, Whitelegge JP, Komenda J. The Photosystem II Assembly Factor Ycf48 from the Cyanobacterium Synechocystis sp. PCC 6803 Is Lipidated Using an Atypical Lipobox Sequence. International Journal of Molecular Sciences. 2021; 22(7):3733. https://doi.org/10.3390/ijms22073733
Chicago/Turabian StyleKnoppová, Jana, Jianfeng Yu, Jan Janouškovec, Petr Halada, Peter J. Nixon, Julian P. Whitelegge, and Josef Komenda. 2021. "The Photosystem II Assembly Factor Ycf48 from the Cyanobacterium Synechocystis sp. PCC 6803 Is Lipidated Using an Atypical Lipobox Sequence" International Journal of Molecular Sciences 22, no. 7: 3733. https://doi.org/10.3390/ijms22073733