
Behavioral Alterations and Decreased Number of Parvalbumin-Positive Interneurons in Wistar Rats after Maternal Immune Activation by Lipopolysaccharide: Sex Matters

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Brain Analysis in P28-Old Rats
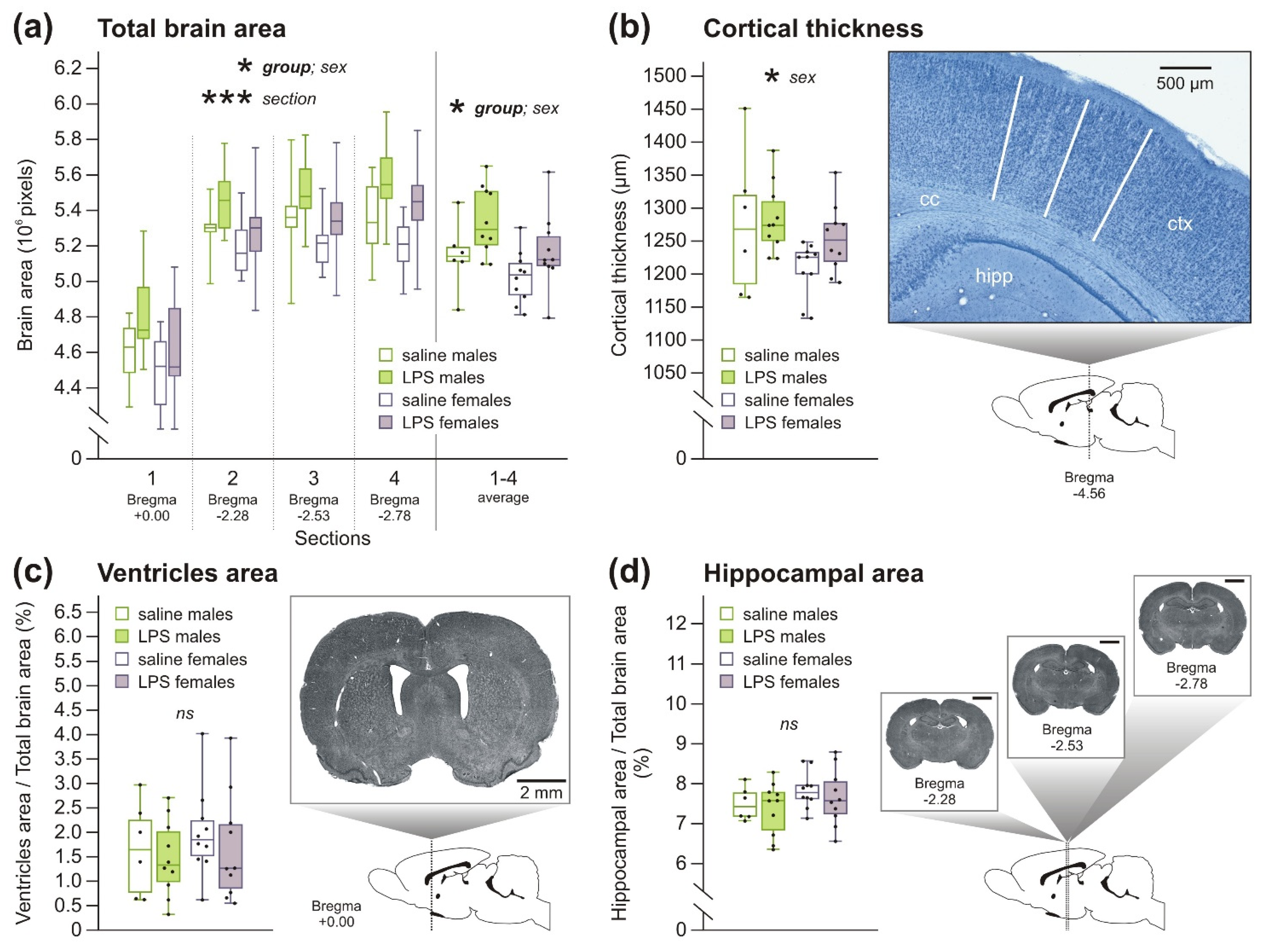
2.1.1. Macrocephaly without Changes in Other Brain Morphology Parameters in Rats after Prenatal Exposure to LPS
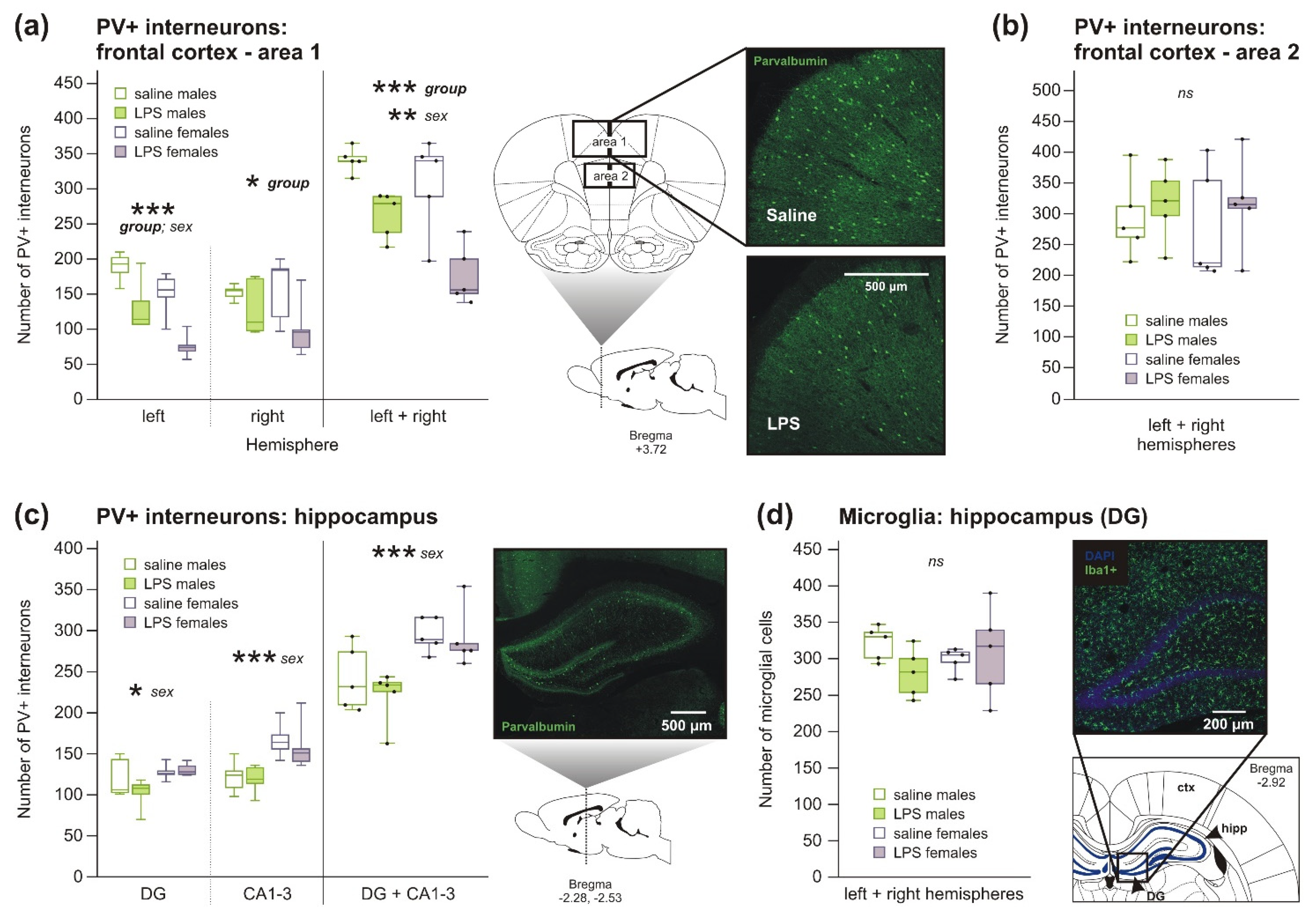
2.1.2. Decreased Number of PV+ Interneurons, but No Changes in Microglia Number, in Rats after Prenatal Exposure to LPS
2.2. Behavior Analysis in P45- and P90-Old Rats
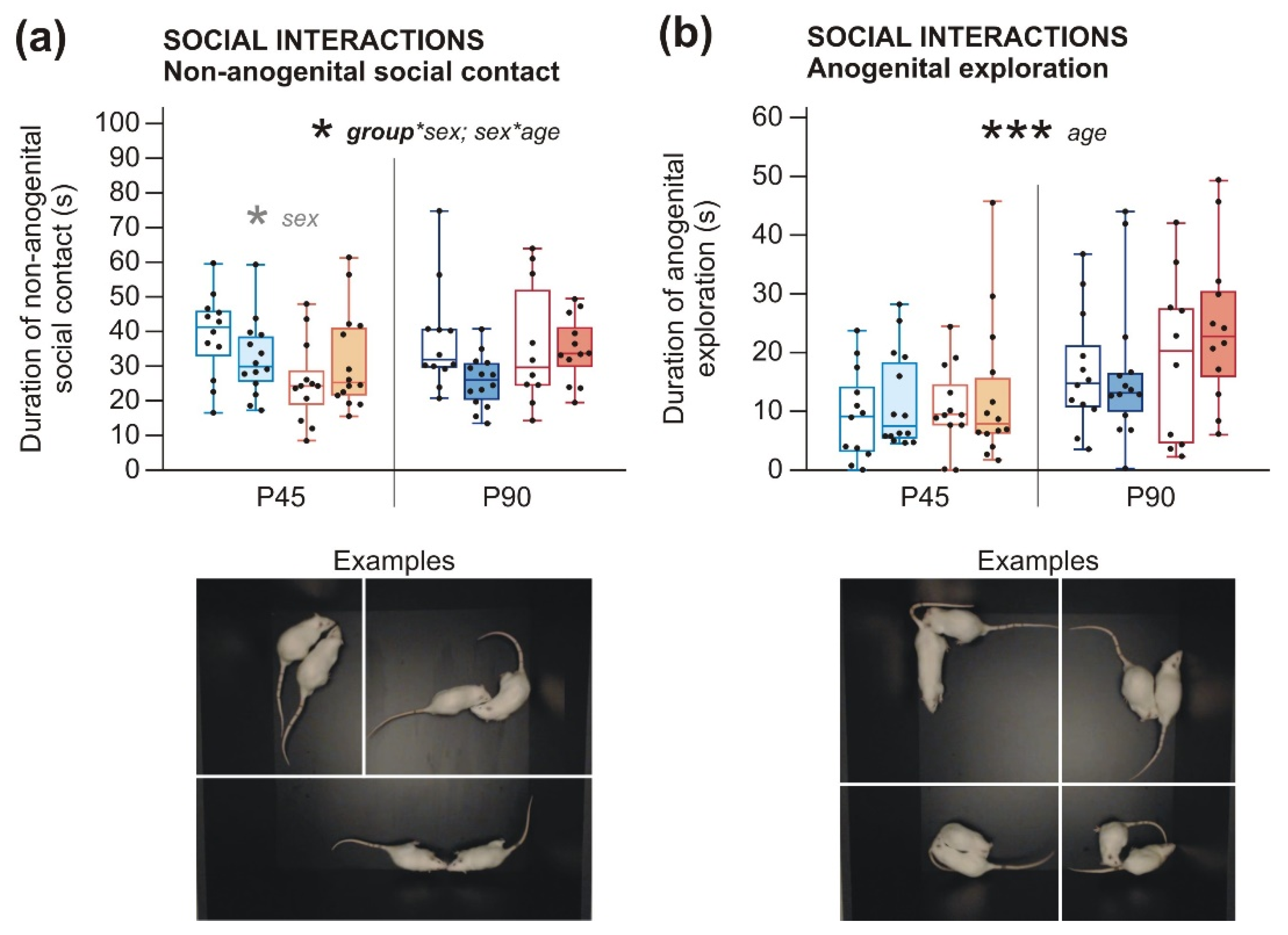
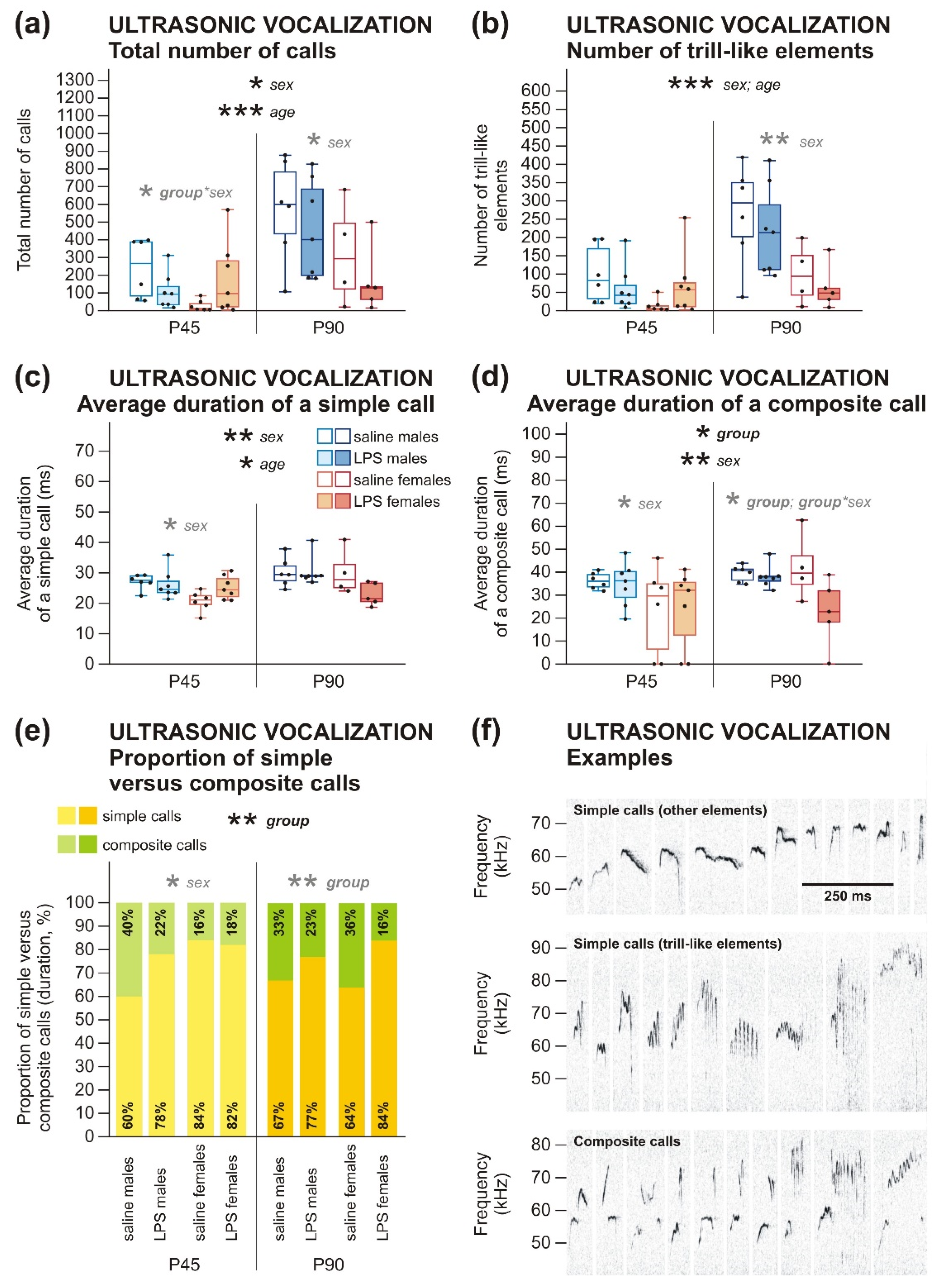
2.2.1. Social Behavior and Communication Deficits after Prenatal LPS Exposure Manifest Earlier in Males Than in Females
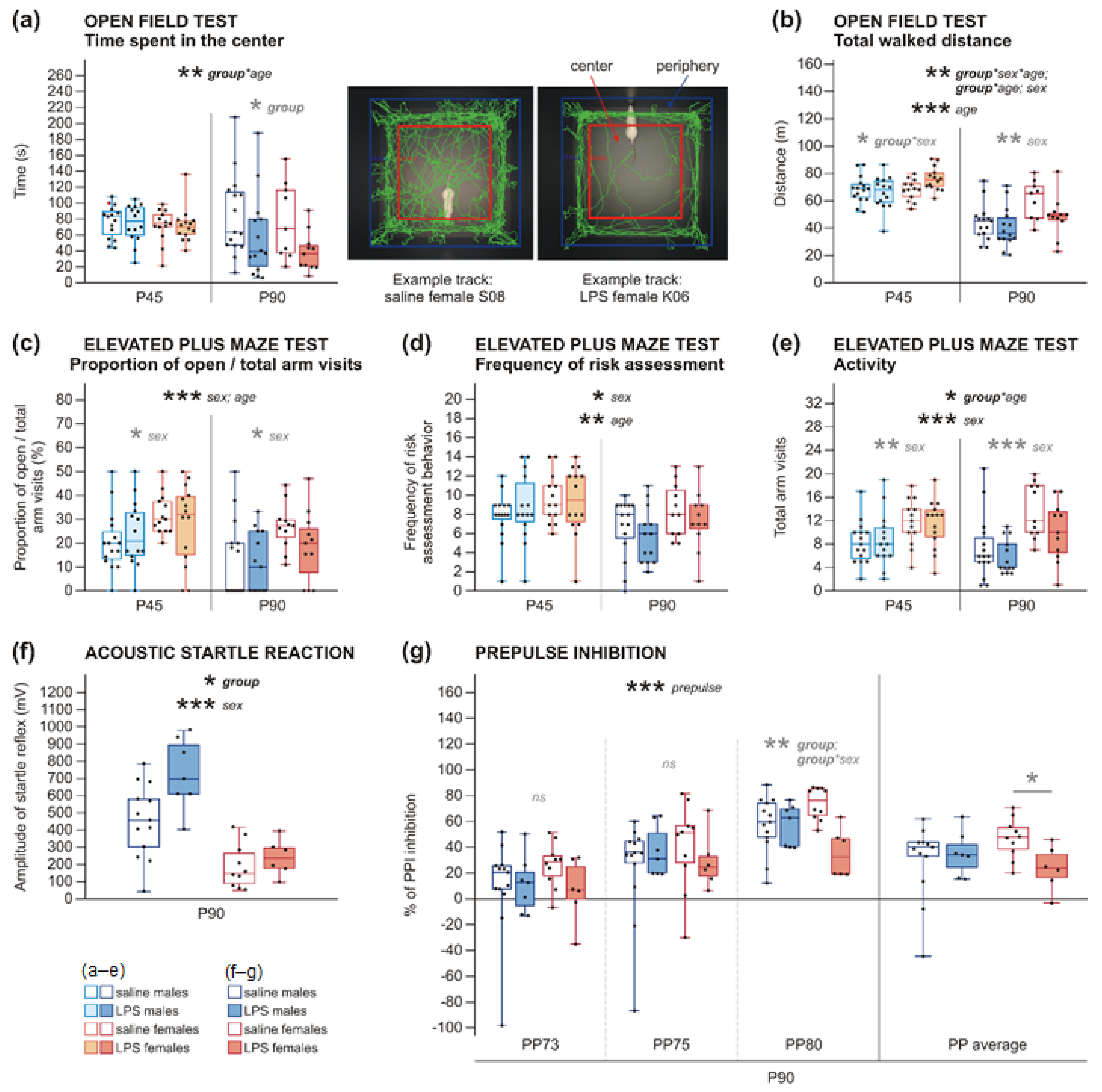
2.2.2. Increased Anxiety in Rats after Prenatal Exposure to LPS
2.2.3. Decreased Activity in Rats after Prenatal Exposure to LPS
2.2.4. Increased Startle Reaction in Adult Rats and PPI Deficit in Adult Females, but Not in Males, after Prenatal Exposure to LPS
3. Discussion
4. Materials and Methods
4.1. Animals
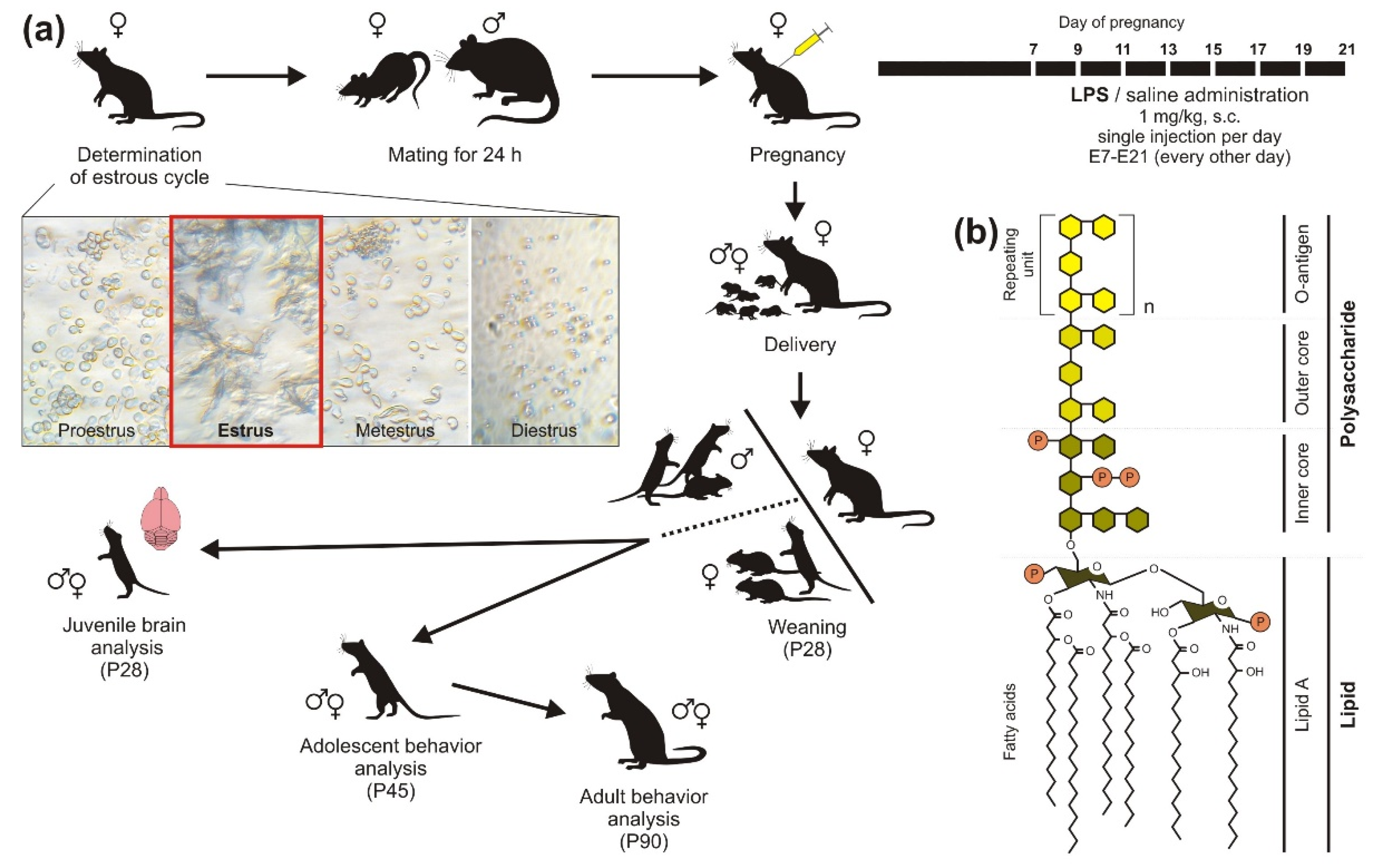
4.2. Model Preparation
4.3. Brain Analysis
4.3.1. Brain Tissue Harvesting
4.3.2. Toluidin Blue Staining and Brain Morphology Analysis
4.3.3. Immunohistochemistry and Analysis of PV+ Interneurons and Microglia
4.4. Behavioral Testing
4.4.1. Open Field Test (OF)
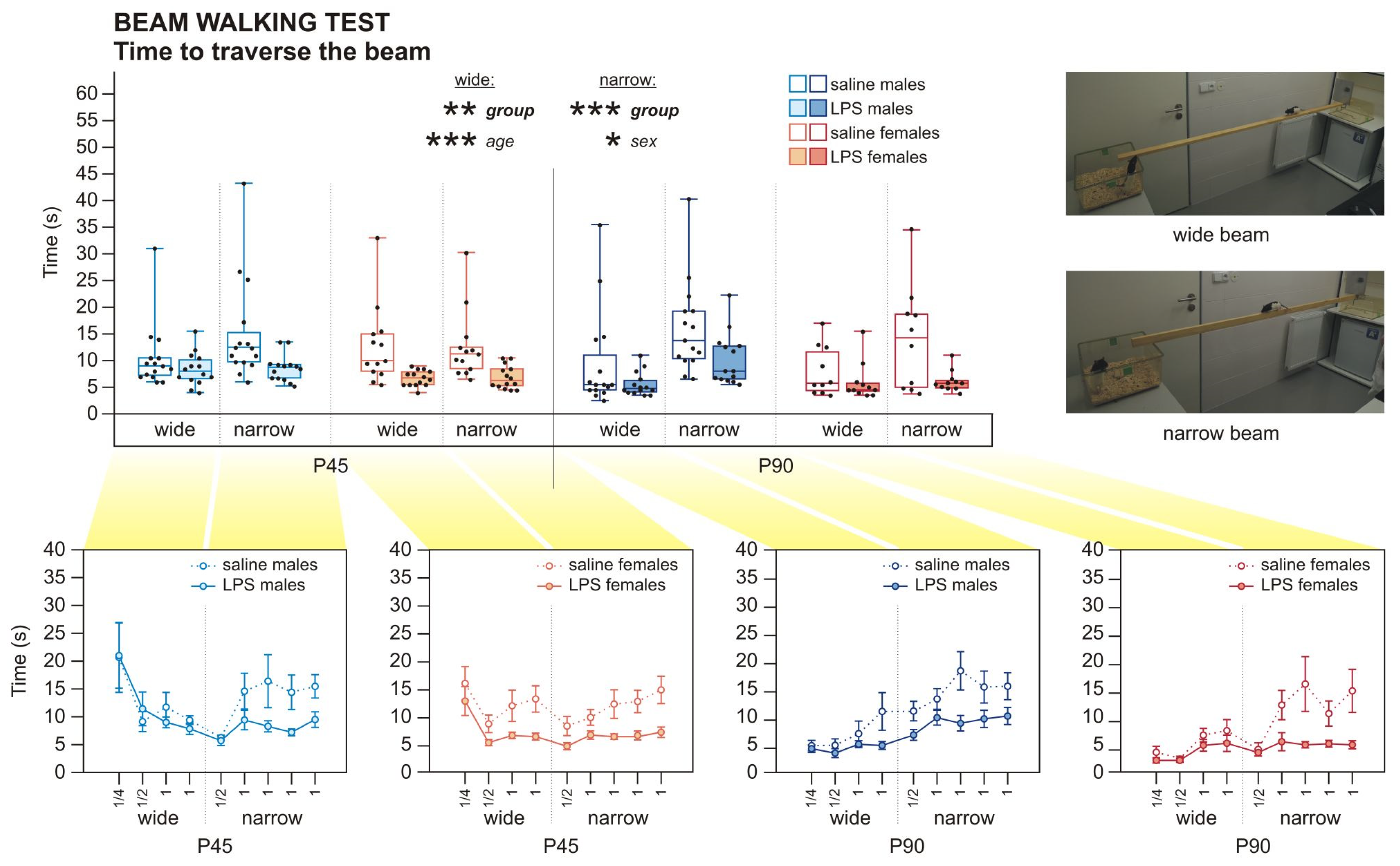
4.4.2. Beam Walking Test (BW)
4.4.3. Elevated Plus Maze Test (EPM)
4.4.4. Social Interactions
4.4.5. Ultrasonic Vocalization Recording (USV)
4.4.6. Acoustic Startle Reaction and Prepulse Inhibition (PPI)
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. (Pregnancy)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % of Females | |
|---|---|
| Total deliveries | 54% |
| Miscarriage/False pregnancy | 29% |
| Not pregnant | 17% |
| Deliveries of LPS dams (per total deliveries) | 54% |

Appendix B. (Statistical Tables)
| Source | df | Error df | F | p | ηp2 |
|---|---|---|---|---|---|
| Total Brain Area—Sections 1–4: 3wANOVA-RM | |||||
| Group | 1 | 32 | 7.020 | 0.012 * | 0.180 |
| Sex | 1 | 32 | 5.223 | 0.029 * | 0.140 |
| Group * sex | 1 | 32 | 0.154 | 0.698 | 0.005 |
| Sections | 2.106 | 67.376 | 329.753 | 0.000 *** | 0.912 |
| Sections * group | 2.106 | 67.376 | 1.163 | 0.321 | 0.035 |
| Sections * sex | 2.106 | 67.376 | 0.101 | 0.912 | 0.003 |
| Sections * group * sex | 2.106 | 67.376 | 0.547 | 0.590 | 0.017 |
| Cortical Thickness: 2wANOVA | |||||
| Group | 1 | 32 | 1.694 | 0.202 | 0.050 |
| Sex | 1 | 32 | 5.188 | 0.030 * | 0.140 |
| Group * sex | 1 | 32 | 0.729 | 0.400 | 0.022 |
| Ventricles Area/Total Brain Area: 2wANOVA | |||||
| Group | 1 | 32 | 0.542 | 0.467 | 0.017 |
| Sex | 1 | 32 | 0.821 | 0.372 | 0.025 |
| Group * sex | 1 | 32 | 0.077 | 0.783 | 0.002 |
| Dorsal Hippocampal Area/Total Brain Area: 2wANOVA | |||||
| Group | 1 | 32 | 0.590 | 0.448 | 0.018 |
| Sex | 1 | 32 | 2.362 | 0.134 | 0.069 |
| Group * sex | 1 | 32 | 0.013 | 0.909 | 0.000 |
| Source | df | Error df | F | p | ηp2 |
|---|---|---|---|---|---|
| PV+ Interneurons–Frontal Cortex, Area 1: 2wANOVA | |||||
| Group | 1 | 16 | 28.099 | 0.000 *** | 0.637 |
| Sex | 1 | 16 | 9.171 | 0.008 ** | 0.364 |
| Group * sex | 1 | 16 | 1.753 | 0.204 | 0.099 |
| PV+ Interneurons–Frontal Cortex, Area 1 (Left Hemisphere): 2wANOVA | |||||
| Group | 1 | 16 | 28.044 | 0.000 *** | 0.637 |
| Sex | 1 | 16 | 14.714 | 0.001 *** | 0.479 |
| Group * sex | 1 | 16 | 0.521 | 0.481 | 0.032 |
| PV+ Interneurons–Frontal Cortex, Area 1 (Right Hemisphere): 2wANOVA | |||||
| Group | 1 | 16 | 5.525 | 0.032 * | 0.257 |
| Sex | 1 | 16 | 0.553 | 0.468 | 0.033 |
| Group * sex | 1 | 16 | 1.064 | 0.318 | 0.062 |
| PV+ Interneurons–Frontal Cortex, Area 2: 2wANOVA | |||||
| Group | 1 | 16 | 0.807 | 0.382 | 0.048 |
| Sex | 1 | 16 | 0.056 | 0.815 | 0.004 |
| Group * sex | 1 | 16 | 0.034 | 0.857 | 0.002 |
| PV+ Interneurons–Dorsal Hippocampus: 2wANOVA | |||||
| Group | 1 | 16 | 0.799 | 0.385 | 0.048 |
| Sex | 1 | 16 | 16.649 | 0.001 *** | 0.510 |
| Group * sex | 1 | 16 | 0.326 | 0.576 | 0.020 |
| PV+ Interneurons–Dorsal Hippocampus (DG): 2wANOVA | |||||
| Group | 1 | 16 | 1.160 | 0.297 | 0.068 |
| Sex | 1 | 16 | 6.090 | 0.025 * | 0.276 |
| Group * sex | 1 | 16 | 2.209 | 0.157 | 0.121 |
| PV+ Interneurons–Dorsal Hippocampus (CA1-3): 2wANOVA | |||||
| Group | 1 | 16 | 0.278 | 0.605 | 0.017 |
| Sex | 1 | 16 | 17.319 | 0.001 *** | 0.520 |
| Group * sex | 1 | 16 | 0.055 | 0.818 | 0.003 |
| Microglia–Dorsal Hippocampus (DG): 2wANOVA | |||||
| Group | 1 | 16 | 0.843 | 0.372 | 0.050 |
| Sex | 1 | 16 | 0.021 | 0.886 | 0.001 |
| Group * sex | 1 | 16 | 2.154 | 0.162 | 0.119 |
| Source | df | Error df | F | p | ηp2 |
|---|---|---|---|---|---|
| Duration of Non-Anogenital Social Contact: 3wANOVA-RM | |||||
| Group | 1 | 44 | 1.212 | 0.277 | 0.027 |
| Sex | 1 | 44 | 0.279 | 0.600 | 0.006 |
| Group * sex | 1 | 44 | 4.880 | 0.032 * | 0.100 |
| Age | 1 | 44 | 0.135 | 0.715 | 0.003 |
| Age * group | 1 | 44 | 0.889 | 0.351 | 0.020 |
| Age * sex | 1 | 44 | 4.300 | 0.044 * | 0.089 |
| Age * group * sex | 1 | 44 | 0.054 | 0.817 | 0.001 |
| Duration of Anogenital Exploration: 3wANOVA-RM | |||||
| Group | 1 | 44 | 0.653 | 0.423 | 0.015 |
| Sex | 1 | 44 | 1.698 | 0.199 | 0.037 |
| Group * sex | 1 | 44 | 0.19 | 0.665 | 0.004 |
| Age | 1 | 44 | 13.822 | 0.001 *** | 0.239 |
| Age * group | 1 | 44 | 0.073 | 0.789 | 0.002 |
| Age * sex | 1 | 44 | 0.745 | 0.393 | 0.017 |
| Age * group * sex | 1 | 44 | 0.811 | 0.373 | 0.018 |
| Source | df | Error df | F | p | ηp2 |
|---|---|---|---|---|---|
| Total Number of Calls: 3wANOVA-RM | |||||
| Group | 1 | 18 | 1.573 | 0.226 | 0.080 |
| Sex | 1 | 18 | 7.321 | 0.014 * | 0.289 |
| Group * sex | 1 | 18 | 0.225 | 0.641 | 0.012 |
| Age | 1 | 18 | 21.815 | 0.000 *** | 0.548 |
| Age * group | 1 | 18 | 0.648 | 0.431 | 0.035 |
| Age * sex | 1 | 18 | 1.753 | 0.202 | 0.089 |
| Age * group * sex | 1 | 18 | 0.919 | 0.351 | 0.049 |
| Total Number of Calls at P45: 2wANOVA | |||||
| Group | 1 | 22 | 0.044 | 0.835 | 0.002 |
| Sex | 1 | 22 | 1.424 | 0.246 | 0.061 |
| Group * sex | 1 | 22 | 6.096 | 0.022 * | 0.217 |
| Total Number of Calls at P90: 2wANOVA | |||||
| Group | 1 | 18 | 1.317 | 0.266 | 0.068 |
| Sex | 1 | 18 | 5.183 | 0.035 * | 0.224 |
| Group * sex | 1 | 18 | 0.030 | 0.863 | 0.002 |
| Number of Trill-Like Elements: 3wANOVA-RM | |||||
| Group | 1 | 18 | 0.875 | 0.362 | 0.046 |
| Sex | 1 | 18 | 14.314 | 0.001 *** | 0.443 |
| Group * sex | 1 | 18 | 0.353 | 0.560 | 0.019 |
| Age | 1 | 18 | 16.385 | 0.001 *** | 0.477 |
| Age * group | 1 | 18 | 0.312 | 0.583 | 0.017 |
| Age * sex | 1 | 18 | 3.871 | 0.065 | 0.177 |
| Age * group * sex | 1 | 18 | 0.181 | 0.675 | 0.010 |
| Average Duration of a Simple Call: 3wANOVA-RM | |||||
| Group | 1 | 18 | 1.321 | 0.265 | 0.068 |
| Sex | 1 | 18 | 12.296 | 0.003 ** | 0.406 |
| Group * sex | 1 | 18 | 0.647 | 0.432 | 0.035 |
| Age | 1 | 18 | 6.919 | 0.017 * | 0.278 |
| Age * group | 1 | 18 | 1.990 | 0.175 | 0.100 |
| Age * sex | 1 | 18 | 0.083 | 0.776 | 0.005 |
| Age * group * sex | 1 | 18 | 3.145 | 0.093 | 0.149 |
| Average Duration of a Composite Call: 3wANOVA-RM | |||||
| Group | 1 | 18 | 4.852 | 0.041 * | 0.212 |
| Sex | 1 | 18 | 10.663 | 0.004 ** | 0.372 |
| Group * sex | 1 | 18 | 3.051 | 0.098 | 0.145 |
| Age | 1 | 18 | 4.201 | 0.055 | 0.189 |
| Age * group | 1 | 18 | 1.007 | 0.329 | 0.053 |
| Age * sex | 1 | 18 | 1.283 | 0.272 | 0.067 |
| Age * group * sex | 1 | 18 | 1.078 | 0.313 | 0.056 |
| Average Duration of a Composite Call at P45: 2wANOVA | |||||
| Group | 1 | 22 | 0.006 | 0.941 | 0.000 |
| Sex | 1 | 22 | 4.484 | 0.046 * | 0.169 |
| Group * sex | 1 | 22 | 0.045 | 0.833 | 0.002 |
| Average Duration of a Composite Call at P90: 2wANOVA | |||||
| Group | 1 | 18 | 6.098 | 0.024 * | 0.253 |
| Sex | 1 | 18 | 2.229 | 0.153 | 0.110 |
| Group * sex | 1 | 18 | 4.66 | 0.045 * | 0.206 |
| Proportion of Simple Versus Composite Calls: 3wANOVA-RM | |||||
| Group | 1 | 18 | 8.376 | 0.010 ** | 0.318 |
| Sex | 1 | 18 | 2.589 | 0.125 | 0.126 |
| Group * sex | 1 | 18 | 0.000 | 0.995 | 0.000 |
| Age | 1 | 18 | 0.529 | 0.476 | 0.029 |
| Age * group | 1 | 18 | 0.084 | 0.776 | 0.005 |
| Age * sex | 1 | 18 | 2.804 | 0.111 | 0.135 |
| Age * group * sex | 1 | 18 | 2.213 | 0.154 | 0.109 |
| Proportion of Simple Versus Composite Calls at P45: 2wANOVA | |||||
| Group | 1 | 22 | 1.602 | 0.219 | 0.068 |
| Sex | 1 | 22 | 4.786 | 0.040 * | 0.179 |
| Group * sex | 1 | 22 | 2.574 | 0.123 | 0.105 |
| Proportion of Simple Versus Composite Calls at P90: 2wANOVA | |||||
| Group | 1 | 18 | 11.52 | 0.003 ** | 0.390 |
| Sex | 1 | 18 | 0.188 | 0.669 | 0.010 |
| Group * sex | 1 | 18 | 1.402 | 0.252 | 0.072 |
| Source | df | Error df | F | p | ηp2 |
|---|---|---|---|---|---|
| Time to Traverse the Wide Beam: 3wANOVA-RM | |||||
| Group | 1 | 46 | 7.789 | 0.008 ** | 0.145 |
| Sex | 1 | 46 | 0.216 | 0.644 | 0.005 |
| Group * sex | 1 | 46 | 0.220 | 0.642 | 0.005 |
| Age | 1 | 46 | 11.717 | 0.001 *** | 0.203 |
| Age * group | 1 | 46 | 0.051 | 0.823 | 0.001 |
| Age * sex | 1 | 46 | 0.300 | 0.586 | 0.006 |
| Age * group * sex | 1 | 46 | 0.751 | 0.391 | 0.016 |
| Time to Traverse the Narrow Beam: 3wANOVA-RM | |||||
| Group | 1 | 46 | 24.401 | 0.000 *** | 0.347 |
| Sex | 1 | 46 | 6.202 | 0.016 * | 0.119 |
| Group * sex | 1 | 46 | 0.371 | 0.545 | 0.008 |
| Age | 1 | 46 | 0.003 | 0.953 | 0.000 |
| Age * group | 1 | 46 | 0.033 | 0.856 | 0.001 |
| Age * sex | 1 | 46 | 1.347 | 0.252 | 0.028 |
| Age * group * sex | 1 | 46 | 0.165 | 0.687 | 0.004 |
| Source | df | Error df | F | p | ηp2 | Simple Effects |
|---|---|---|---|---|---|---|
| Time Spent in the Center: 3wANOVA-RM | ||||||
| Group | 1 | 45 | 3.330 | 0.075 | 0.069 | |
| Sex | 1 | 45 | 1.143 | 0.291 | 0.025 | |
| Group * sex | 1 | 45 | 0.018 | 0.894 | 0.000 | |
| Age | 1 | 45 | 1.578 | 0.216 | 0.034 | |
| Age * group | 1 | 45 | 7.761 | 0.008 ** | 0.147 | 0.931 (group at P45) 0.016 * (group at P90) |
| Age * sex | 1 | 45 | 0.315 | 0.578 | 0.007 | |
| Age * group * sex | 1 | 45 | 0.726 | 0.399 | 0.016 | |
| Time Spent in the Center at P45: 2wANOVA | ||||||
| Group | 1 | 52 | 0.047 | 0.829 | 0.001 | |
| Sex | 1 | 52 | 0.479 | 0.492 | 0.009 | |
| Group * sex | 1 | 52 | 0.044 | 0.836 | 0.001 | |
| Time Spent in the Center at P90: 2wANOVA | ||||||
| Group | 1 | 45 | 6.016 | 0.018 * | 0.118 | |
| Sex | 1 | 45 | 0.886 | 0.352 | 0.019 | |
| Group * sex | 1 | 45 | 0.230 | 0.634 | 0.005 | |
| Total Walked Distance: 3wANOVA-RM | ||||||
| Group | 1 | 45 | 0.529 | 0.471 | 0.012 | |
| Sex | 1 | 45 | 8.251 | 0.006 ** | 0.155 | |
| Group * sex | 1 | 45 | 0.057 | 0.812 | 0.001 | |
| Age | 1 | 45 | 127.226 | 0.000 *** | 0.739 | |
| Age * group | 1 | 45 | 9.420 | 0.004 ** | 0.173 | 0.351 (group at P45) 0.151 (group at P90) |
| Age * sex | 1 | 45 | 3.569 | 0.065 | 0.073 | |
| Age * group * sex | 1 | 45 | 7.674 | 0.008 ** | 0.146 | |
| Total Walked Distance at P45: 2wANOVA | ||||||
| Group | 1 | 52 | 1.214 | 0.276 | 0.023 | |
| Sex | 1 | 52 | 3.374 | 0.072 | 0.061 | |
| Group * sex | 1 | 52 | 4.093 | 0.048 * | 0.073 | |
| Total Walked Distance at P90: 2wANOVA | ||||||
| Group | 1 | 45 | 3.752 | 0.059 | 0.077 | |
| Sex | 1 | 45 | 8.813 | 0.005 ** | 0.164 | |
| Group * sex | 1 | 45 | 1.193 | 0.280 | 0.026 | |
| Source | df | Error df | F | p | ηp2 | Simple Effects |
|---|---|---|---|---|---|---|
| Proportion of Open Arm/Total Arm Visits: 3wANOVA-RM | ||||||
| Group | 1 | 46 | 0.190 | 0.665 | 0.004 | |
| Sex | 1 | 46 | 13.877 | 0.001 *** | 0.232 | |
| Group * sex | 1 | 46 | 1.464 | 0.233 | 0.031 | |
| Age | 1 | 46 | 15.697 | 0.000 *** | 0.254 | |
| Age * group | 1 | 46 | 1.803 | 0.186 | 0.038 | |
| Age * sex | 1 | 46 | 0.058 | 0.811 | 0.001 | |
| Age * group * sex | 1 | 46 | 0.026 | 0.872 | 0.001 | |
| Frequency of Risk Assessment Behavior: 3wANOVA-RM | ||||||
| Group | 1 | 46 | 0.049 | 0.825 | 0.001 | |
| Sex | 1 | 46 | 4.330 | 0.043 * | 0.086 | |
| Group * sex | 1 | 46 | 0.120 | 0.730 | 0.003 | |
| Age | 1 | 46 | 11.098 | 0.002 ** | 0.194 | |
| Age * group | 1 | 46 | 2.444 | 0.125 | 0.050 | |
| Age * sex | 1 | 46 | 0.439 | 0.511 | 0.009 | |
| Age * group * sex | 1 | 46 | 0.052 | 0.820 | 0.001 | |
| Total Arm Visits: 3wANOVA-RM | ||||||
| Group | 1 | 46 | 0.980 | 0.327 | 0.021 | |
| Sex | 1 | 46 | 16.890 | 0.000 *** | 0.269 | |
| Group * sex | 1 | 46 | 0.501 | 0.483 | 0.011 | |
| Age | 1 | 46 | 2.408 | 0.128 | 0.050 | |
| Age * group | 1 | 46 | 4.696 | 0.035 * | 0.093 | 0.596 (group at P45) 0.147 (group at P90) |
| Age * sex | 1 | 46 | 2.211 | 0.144 | 0.046 | |
| Age * group * sex | 1 | 46 | 0.135 | 0.715 | 0.003 | |
| Total Arm Visits at P45: 2wANOVA | ||||||
| Group | 1 | 52 | 0.013 | 0.910 | 0.000 | |
| Sex | 1 | 52 | 7.927 | 0.007 ** | 0.132 | |
| Group * sex | 1 | 52 | 0.127 | 0.723 | 0.002 | |
| Total Arm Visits at P90: 2wANOVA | ||||||
| Group | 1 | 46 | 3.600 | 0.064 | 0.073 | |
| Sex | 1 | 46 | 15.740 | 0.000 *** | 0.255 | |
| Group * sex | 1 | 46 | 0.549 | 0.462 | 0.012 | |
| Source | df | Error df | F | p | ηp2 |
|---|---|---|---|---|---|
| Acoustic Startle Reaction: 2wANOVA | |||||
| Group | 1 | 32 | 6.873 | 0.013 * | 0.177 |
| Sex | 1 | 32 | 36.286 | 0.000 *** | 0.531 |
| Group * sex | 1 | 32 | 3.261 | 0.080 | 0.092 |
| PPI–All Prepulses: 3wANOVA-RM | |||||
| Group | 1 | 32 | 1.508 | 0.228 | 0.045 |
| Sex | 1 | 32 | 0.111 | 0.741 | 0.003 |
| Group * sex | 1 | 32 | 3.322 | 0.078 | 0.094 |
| Prepulse | 2 | 64 | 10.976 | 0.000 *** | 0.561 |
| Prepulse * group | 2 | 64 | 2.168 | 0.123 | 0.063 |
| Prepulse * sex | 2 | 64 | 0.429 | 0.653 | 0.013 |
| Prepulse * group * sex | 2 | 64 | 0.466 | 0.629 | 0.014 |
| PPI–Prepulse 73 dB: 2wANOVA | |||||
| Group | 1 | 32 | 0.868 | 0.358 | 0.026 |
| Sex | 1 | 32 | 0.339 | 0.564 | 0.010 |
| Group * sex | 1 | 32 | 1.195 | 0.283 | 0.036 |
| PPI–Prepulse 75 dB: 2wANOVA | |||||
| Group | 1 | 32 | 0.001 | 0.978 | 0.000 |
| Sex | 1 | 32 | 0.151 | 0.701 | 0.005 |
| Group * sex | 1 | 32 | 1.107 | 0.301 | 0.033 |
| PPI–Prepulse 80 dB: 2wANOVA | |||||
| Group | 1 | 32 | 9.369 | 0.004 ** | 0.226 |
| Sex | 1 | 32 | 0.151 | 0.700 | 0.005 |
| Group * sex | 1 | 32 | 9.154 | 0.005 ** | 0.222 |
| df | t | p | |||
| PPI–Averaged for All Prepulses, Females: t-test | |||||
| Group | 14 | 2.760 | 0.015 * | ||
| PPI–Prepulse 73 dB, Females: t-test | |||||
| Group | 14 | 1.877 | 0.081 | ||
| PPI–Prepulse 75 dB, Females: t-test | |||||
| Group | 14 | 0.745 | 0.469 | ||
| PPI–Prepulse 80 dB, Females: t-test | |||||
| Group | 14 | 4.920 | 0.000 *** | ||
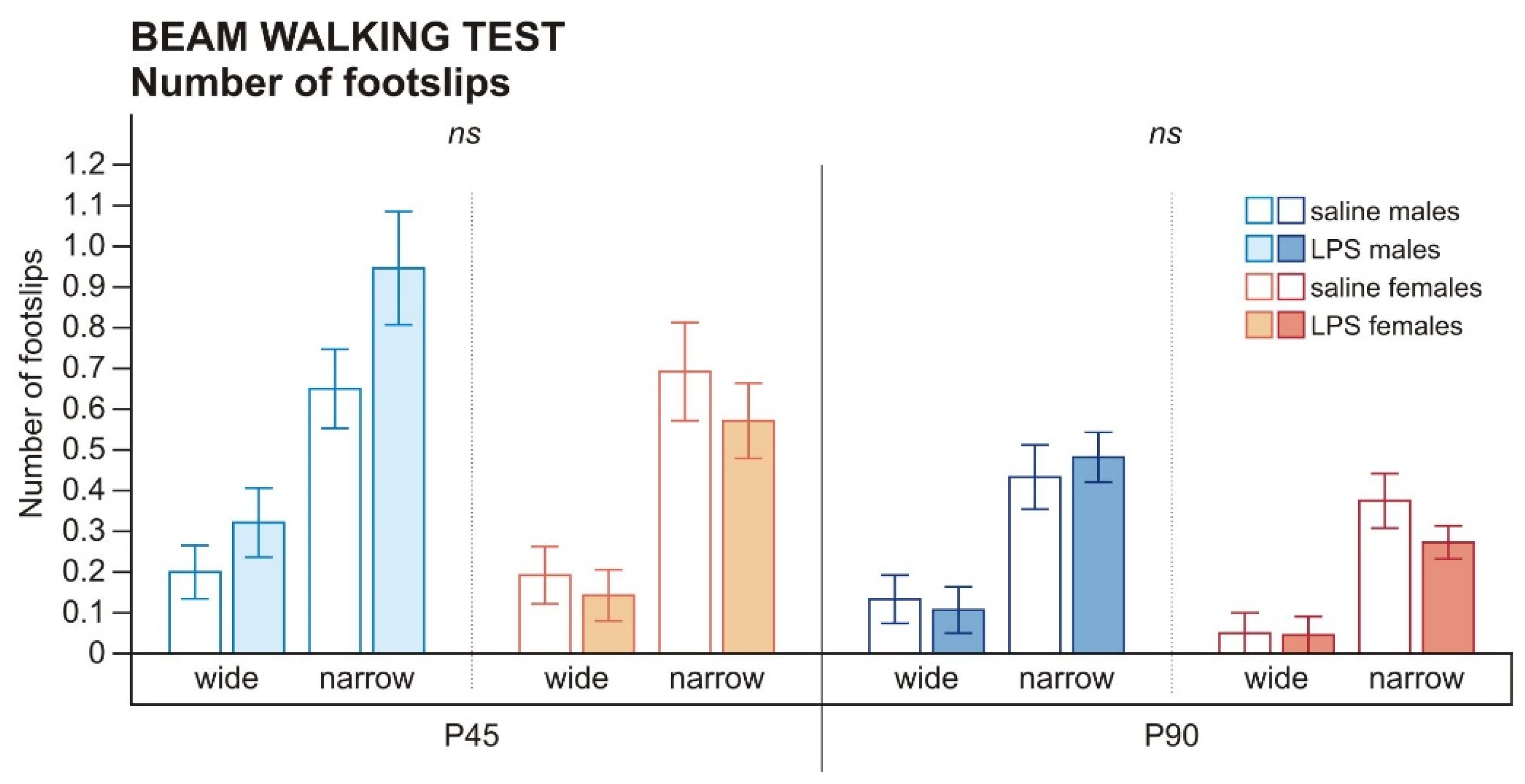
Appendix C. (Number of Footslips in the Beam Walking Test)
| Source | Omnibus Test |
|---|---|
| Number of Footslips on the Wide Beam: Negative Binomial Log Estimates Values | |
| Group (P45) | 0.899 |
| Group (P90) | 0.941 |
| Number of Footslips on the Narrow Beam: Negative Binomial Log Estimates Values | |
| Group (P45) | 0.307 |
| Group (P90) | 0.310 |

Appendix D. (Estrous Cycle Determination)
References
- Brown, A.S. Epidemiologic Studies of Exposure to Prenatal Infection and Risk of Schizophrenia and Autism. Dev. Neurobiol. 2012, 72, 1272–1276. [Google Scholar] [CrossRef] [Green Version]
- Buka, S.L.; Tsuang, M.T.; Torrey, E.F.; Klebanoff, M.A.; Wagner, R.L.; Yolken, R.H. Maternal Cytokine Levels during Pregnancy and Adult Psychosis. Brain. Behav. Immun. 2001, 15, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.; Malchow, B.; Hasan, A.; Falkai, P. The Impact of Environmental Factors in Severe Psychiatric Disorders. Front. Neurosci. 2014, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tordjman, S.; Drapier, D.; Bonnot, O.; Graignic, R.; Fortes, S.; Cohen, D.; Millet, B.; Laurent, C.; Roubertoux, P.L. Animal Models Relevant to Schizophrenia and Autism: Validity and Limitations. Behav. Genet. 2007, 37, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Volkmar, F.R.; Lord, C.; Bailey, A.; Schultz, R.T.; Klin, A. Autism and Pervasive Developmental Disorders. J. Child Psychol. Psychiatry 2004, 45, 135–170. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®®), 5th ed.; American Psychiatric Association: Arlington, VA, USA, 2013; ISBN 978-0-89042-557-2. [Google Scholar]
- Owen, M.J.; Sawa, A.; Mortensen, P.B. Schizophrenia. Lancet 2016, 388, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Kodish, I.; McClellan, J.M. Early onset schizophrenia. In Dulcan’s Textbook of Child and Adolescent Psychiatry, 2nd ed.; American Psychiatric Publishing, Inc.: Arlington, VA, USA, 2016; pp. 389–408. ISBN 978-1-58562-493-5. [Google Scholar]
- Parnas, J.; Bovet, P.; Zahavi, D. Schizophrenic Autism: Clinical Phenomenology and Pathogenetic Implications. World Psychiatry 2002, 1, 131–136. [Google Scholar]
- Pina-Camacho, L.; Parellada, M.; Kyriakopoulos, M. Autism Spectrum Disorder and Schizophrenia: Boundaries and Uncertainties. BJPsych Adv. 2016, 22, 316–324. [Google Scholar] [CrossRef]
- Rimol, L.M.; Hartberg, C.B.; Nesvåg, R.; Fennema-Notestine, C.; Hagler, D.J.; Pung, C.J.; Jennings, R.G.; Haukvik, U.K.; Lange, E.; Nakstad, P.H.; et al. Cortical Thickness and Subcortical Volumes in Schizophrenia and Bipolar Disorder. Biol. Psychiatry 2010, 68, 41–50. [Google Scholar] [CrossRef]
- Amaral, D.G.; Schumann, C.M.; Nordahl, C.W. Neuroanatomy of Autism. Trends Neurosci. 2008, 31, 137–145. [Google Scholar] [CrossRef]
- Courchesne, E.; Carper, R.; Akshoomoff, N. Evidence of Brain Overgrowth in the First Year of Life in Autism. JAMA 2003, 290, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Hadjikhani, N.; Joseph, R.M.; Snyder, J.; Tager-Flusberg, H. Anatomical Differences in the Mirror Neuron System and Social Cognition Network in Autism. Cereb. Cortex 2006, 16, 1276–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardan, A.Y.; Muddasani, S.; Vemulapalli, M.; Keshavan, M.S.; Minshew, N.J. An MRI Study of Increased Cortical Thickness in Autism. Am. J. Psychiatry 2006, 163, 1290–1292. [Google Scholar] [CrossRef] [Green Version]
- Schumann, C.M.; Hamstra, J.; Goodlin-Jones, B.L.; Lotspeich, L.J.; Kwon, H.; Buonocore, M.H.; Lammers, C.R.; Reiss, A.L.; Amaral, D.G. The Amygdala Is Enlarged in Children but Not Adolescents with Autism; the Hippocampus Is Enlarged at All Ages. J. Neurosci. 2004, 24, 6392–6401. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Burgos, G.; Cho, R.Y.; Lewis, D.A. Alterations in Cortical Network Oscillations and Parvalbumin Neurons in Schizophrenia. Biol. Psychiatry 2015, 77, 1031–1040. [Google Scholar] [CrossRef] [Green Version]
- Hashemi, E.; Ariza, J.; Rogers, H.; Noctor, S.C.; Martínez-Cerdeño, V. The Number of Parvalbumin-Expressing Interneurons Is Decreased in the Prefrontal Cortex in Autism. Cereb. Cortex 2017, 27, 1931–1943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, Y.A.; Kemper, T.L.; Bauman, M.L.; Blatt, G.J. Parvalbumin-, Calbindin-, and Calretinin-Immunoreactive Hippocampal Interneuron Density in Autism. Acta Neurol. Scand. 2010, 121, 99–108. [Google Scholar] [CrossRef]
- Stedehouder, J.; Kushner, S.A. Myelination of Parvalbumin Interneurons: A Parsimonious Locus of Pathophysiological Convergence in Schizophrenia. Mol. Psychiatry 2017, 22, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajcovic, B.; Fajnerova, I.; Horacek, J.; Kelemen, E.; Kubik, S.; Svoboda, J.; Stuchlik, A. Neural and Neuronal Discoordination in Schizophrenia: From Ensembles through Networks to Symptoms. Acta Physiol. 2019, 226, e13282. [Google Scholar] [CrossRef]
- Nakamura, T.; Matsumoto, J.; Takamura, Y.; Ishii, Y.; Sasahara, M.; Ono, T.; Nishijo, H. Relationships among Parvalbumin-Immunoreactive Neuron Density, Phase-Locked Gamma Oscillations, and Autistic/Schizophrenic Symptoms in PDGFR-β Knock-out and Control Mice. PLoS ONE 2015, 10, e0119258. [Google Scholar] [CrossRef]
- Nguyen, R.; Morrissey, M.D.; Mahadevan, V.; Cajanding, J.D.; Woodin, M.A.; Yeomans, J.S.; Takehara-Nishiuchi, K.; Kim, J.C. Parvalbumin and GAD65 Interneuron Inhibition in the Ventral Hippocampus Induces Distinct Behavioral Deficits Relevant to Schizophrenia. J. Neurosci. 2014, 34, 14948–14960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkowicz, S.R.; Featherby, T.J.; Qu, Z.; Giousoh, A.; Borg, N.A.; Heng, J.I.; Whisstock, J.C.; Bird, P.I. Brinp1 −/− Mice Exhibit Autism-like Behaviour, Altered Memory, Hyperactivity and Increased Parvalbumin-Positive Cortical Interneuron Density. Mol. Autism 2016, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Saunders, J.A.; Tatard-Leitman, V.M.; Suh, J.; Billingslea, E.N.; Roberts, T.P.; Siegel, S.J. Knockout of NMDA Receptors in Parvalbumin Interneurons Recreates Autism-like Phenotypes. Autism Res. 2013, 6, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Wöhr, M.; Orduz, D.; Gregory, P.; Moreno, H.; Khan, U.; Vörckel, K.J.; Wolfer, D.P.; Welzl, H.; Gall, D.; Schiffmann, S.N.; et al. Lack of Parvalbumin in Mice Leads to Behavioral Deficits Relevant to All Human Autism Core Symptoms and Related Neural Morphofunctional Abnormalities. Transl. Psychiatry 2015, 5, e525. [Google Scholar] [CrossRef] [PubMed]
- Baharnoori, M.; Bhardwaj, S.K.; Srivastava, L.K. Neonatal Behavioral Changes in Rats with Gestational Exposure to Lipopolysaccharide: A Prenatal Infection Model for Developmental Neuropsychiatric Disorders. Schizophr. Bull. 2012, 38, 444–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basta-Kaim, A.; Fijał, K.; Budziszewska, B.; Regulska, M.; Leśkiewicz, M.; Kubera, M.; Gołembiowska, K.; Lasoń, W.; Wędzony, K. Prenatal Lipopolysaccharide Treatment Enhances MK-801-Induced Psychotomimetic Effects in Rats. Pharmacol. Biochem. Behav. 2011, 98, 241–249. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Reutiman, T.J.; Folsom, T.D.; Huang, H.; Oishi, K.; Mori, S.; Smee, D.F.; Pearce, D.A.; Winter, C.; Sohr, R.; et al. Maternal Infection Leads to Abnormal Gene Regulation and Brain Atrophy in Mouse Offspring: Implications for Genesis of Neurodevelopmental Disorders. Schizophr. Res. 2008, 99, 56–70. [Google Scholar] [CrossRef] [Green Version]
- Meyer, U. Prenatal Poly(I:C) Exposure and Other Developmental Immune Activation Models in Rodent Systems. Biol. Psychiatry 2014, 75, 307–315. [Google Scholar] [CrossRef]
- Nakamura, J.P.; Gillespie, B.; Gibbons, A.; Jaehne, E.J.; Du, X.; Chan, A.; Schroeder, A.; van den Buuse, M.; Sundram, S.; Hill, R.A. Maternal Immune Activation Targeted to a Window of Parvalbumin Interneuron Development Improves Spatial Working Memory: Implications for Autism. Brain. Behav. Immun. 2021, 91, 339–349. [Google Scholar] [CrossRef]
- Patterson, P.H. Maternal Infection and Immune Involvement in Autism. Trends Mol. Med. 2011, 17, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Batinić, B.; Santrač, A.; Divović, B.; Timić, T.; Stanković, T.; Obradović, A.L.; Joksimović, S.; Savić, M.M. Lipopolysaccharide Exposure during Late Embryogenesis Results in Diminished Locomotor Activity and Amphetamine Response in Females and Spatial Cognition Impairment in Males in Adult, but Not Adolescent Rat Offspring. Behav. Brain Res. 2016, 299, 72–80. [Google Scholar] [CrossRef]
- Henry, C.J.; Huang, Y.; Wynne, A.M.; Godbout, J.P. Peripheral Lipopolysaccharide (LPS) Challenge Promotes Microglial Hyperactivity in Aged Mice That Is Associated with Exaggerated Induction of Both pro-Inflammatory IL-1β and Anti-Inflammatory IL-10 Cytokines. Brain Behav. Immun. 2009, 23, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solati, J.; Kleehaupt, E.; Kratz, O.; Moll, G.H.; Golub, Y. Inverse Effects of Lipopolysaccharides on Anxiety in Pregnant Mice and Their Offspring. Physiol. Behav. 2015, 139, 369–374. [Google Scholar] [CrossRef]
- Van Amersfoort, E.S.; Van Berkel, T.J.C.; Kuiper, J. Receptors, Mediators, and Mechanisms Involved in Bacterial Sepsis and Septic Shock. Clin. Microbiol. Rev. 2003, 16, 379–414. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Pan, Z.-L.; Pang, Y.; Evans, O.B.; Rhodes, P.G. Cytokine Induction in Fetal Rat Brains and Brain Injury in Neonatal Rats after Maternal Lipopolysaccharide Administration. Pediatr. Res. 2000, 47, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, E.; Guaza, C.; Castellano, B.; Borrell, J. Ontogeny of Sensorimotor Gating and Immune Impairment Induced by Prenatal Immune Challenge in Rats: Implications for the Etiopathology of Schizophrenia. Mol. Psychiatry 2010, 15, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boksa, P. Effects of Prenatal Infection on Brain Development and Behavior: A Review of Findings from Animal Models. Brain Behav. Immun. 2010, 24, 881–897. [Google Scholar] [CrossRef]
- McPartland, J.C.; Law, K.; Dawson, G. Autism Spectrum Disorder. In Encyclopedia of Mental Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 124–130. ISBN 978-0-12-397753-3. [Google Scholar]
- Ochoa, S.; Usall, J.; Cobo, J.; Labad, X.; Kulkarni, J. Gender Differences in Schizophrenia and First-Episode Psychosis: A Comprehensive Literature Review. Schizophr. Res. Treat. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Werling, D.M.; Geschwind, D.H. Sex Differences in Autism Spectrum Disorders. Curr. Opin. Neurol. 2013, 26, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Potasiewicz, A.; Holuj, M.; Piotrowska, D.; Zajda, K.; Wojcik, M.; Popik, P.; Nikiforuk, A. Evaluation of Ultrasonic Vocalizations in a Neurodevelopmental Model of Schizophrenia during the Early Life Stages of Rats. Neuropharmacology 2019, 146, 28–38. [Google Scholar] [CrossRef]
- Basta-Kaim, A.; Fijał, K.; Ślusarczyk, J.; Trojan, E.; Głombik, K.; Budziszewska, B.; Leśkiewicz, M.; Regulska, M.; Kubera, M.; Lasoń, W.; et al. Prenatal Administration of Lipopolysaccharide Induces Sex-Dependent Changes in Glutamic Acid Decarboxylase and Parvalbumin in the Adult Rat Brain. Neuroscience 2015, 287, 78–92. [Google Scholar] [CrossRef]
- Wischhof, L.; Irrsack, E.; Osorio, C.; Koch, M. Prenatal LPS-Exposure—A Neurodevelopmental Rat Model of Schizophrenia—Differentially Affects Cognitive Functions, Myelination and Parvalbumin Expression in Male and Female Offspring. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 57, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Frankle, W.G.; Cho, R.Y.; Prasad, K.M.; Mason, N.S.; Paris, J.; Himes, M.L.; Walker, C.; Lewis, D.A.; Narendran, R. In Vivo Measurement of GABA Transmission in Healthy Subjects and Schizophrenia Patients. Am. J. Psychiatry 2015, 172, 1148–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curley, A.A.; Arion, D.; Volk, D.W.; Asafu-Adjei, J.K.; Sampson, A.R.; Fish, K.N.; Lewis, D.A. Cortical Deficits of Glutamic Acid Decarboxylase 67 Expression in Schizophrenia: Clinical, Protein, and Cell Type-Specific Features. Am. J. Psychiatry 2011, 168, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Glausier, J.R.; Fish, K.N.; Lewis, D.A. Altered Parvalbumin Basket Cell Inputs in the Dorsolateral Prefrontal Cortex of Schizophrenia Subjects. Mol. Psychiatry 2014, 19, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Konradi, C.; Yang, C.K.; Zimmerman, E.I.; Lohmann, K.M.; Gresch, P.; Pantazopoulos, H.; Berretta, S.; Heckers, S. Hippocampal Interneurons are Abnormal in Schizophrenia. Schizophr. Res. 2011, 131, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Burgos, G.; Hashimoto, T.; Lewis, D.A. Alterations of Cortical GABA Neurons and Network Oscillations in Schizophrenia. Curr. Psychiatry Rep. 2010, 12, 335–344. [Google Scholar] [CrossRef]
- Lewis, D.A.; Curley, A.A.; Glausier, J.R.; Volk, D.W. Cortical Parvalbumin Interneurons and Cognitive Dysfunction in Schizophrenia. Trends Neurosci. 2012, 35, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Kirsten, T.B.; Taricano, M.; Maiorka, P.C.; Palermo-Neto, J.; Bernardi, M.M. Prenatal Lipopolysaccharide Reduces Social Behavior in Male Offspring. Neuroimmunomodulation 2010, 17, 240–251. [Google Scholar] [CrossRef]
- Chen, Z.; Jalabi, W.; Shpargel, K.B.; Farabaugh, K.T.; Dutta, R.; Yin, X.; Kidd, G.J.; Bergmann, C.C.; Stohlman, S.A.; Trapp, B.D. Lipopolysaccharide-Induced Microglial Activation and Neuroprotection against Experimental Brain Injury Is Independent of Hematogenous TLR4. J. Neurosci. 2012, 32, 11706–11715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Dai, X.; Roller, A.; Carter, K.; Paul, I.; Bhatt, A.J.; Lin, R.C.S.; Fan, L.-W. Early Postnatal Lipopolysaccharide Exposure Leads to Enhanced Neurogenesis and Impaired Communicative Functions in Rats. PLoS ONE 2016, 11, e0164403. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, E.; Karns, C.M.; Davis, H.R.; Ziccardi, R.; Carper, R.A.; Tigue, Z.D.; Chisum, H.J.; Moses, P.; Pierce, K.; Lord, C.; et al. Unusual Brain Growth Patterns in Early Life in Patients with Autistic Disorder: An MRI Study. Neurology 2001, 57, 245–254. [Google Scholar] [CrossRef]
- Dawson, G.; Munson, J.; Webb, S.J.; Nalty, T.; Abbott, R.; Toth, K. Rate of Head Growth Decelerates and Symptoms Worsen in the Second Year of Life in Autism. Biol. Psychiatry 2007, 61, 458–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dementieva, Y.A.; Vance, D.D.; Donnelly, S.L.; Elston, L.A.; Wolpert, C.M.; Ravan, S.A.; DeLong, G.R.; Abramson, R.K.; Wright, H.H.; Cuccaro, M.L. Accelerated Head Growth in Early Development of Individuals with Autism. Pediatr. Neurol. 2005, 32, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Barkataki, I.; Kumari, V.; Das, M.; Taylor, P.; Sharma, T. Volumetric Structural Brain Abnormalities in Men with Schizophrenia or Antisocial Personality Disorder. Behav. Brain Res. 2006, 169, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Foley, K.A.; MacFabe, D.F.; Vaz, A.; Ossenkopp, K.; Kavaliers, M. Sexually Dimorphic Effects of Prenatal Exposure to Propionic Acid and Lipopolysaccharide on Social Behavior in Neonatal, Adolescent, and Adult Rats: Implications for Autism Spectrum Disorders. Int. J. Dev. Neurosci. 2014, 39, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Vojtechova, I.; Petrasek, T.; Maleninska, K.; Brozka, H.; Tejkalova, H.; Horacek, J.; Stuchlik, A.; Vales, K. Neonatal Immune Activation by Lipopolysaccharide Causes Inadequate Emotional Responses to Novel Situations but No Changes in Anxiety or Cognitive Behavior in Wistar Rats. Behav. Brain Res. 2018, 349, 42–53. [Google Scholar] [CrossRef]
- DeLisi, L.E. Speech Disorder in Schizophrenia: Review of the Literature and Exploration of Its Relation to the Uniquely Human Capacity for Language. Schizophr. Bull. 2001, 27, 481–496. [Google Scholar] [CrossRef]
- McKenna, P.J.; Oh, T.M.; Oh, T. Schizophrenic Speech: Making Sense of Bathroots and Ponds That Fall in Doorways; Cambridge University Press: Cambridge, UK, 2005; ISBN 978-0-521-81075-3. [Google Scholar]
- Hogg, S. A Review of the Validity and Variability of the Elevated Plus-Maze as an Animal Model of Anxiety. Pharmacol. Biochem. Behav. 1996, 54, 21–30. [Google Scholar] [CrossRef]
- Ramos, A. Animal Models of Anxiety: Do I Need Multiple Tests? Trends Pharmacol. Sci. 2008, 29, 493–498. [Google Scholar] [CrossRef]
- Depino, A.M. Early Prenatal Exposure to LPS Results in Anxiety- and Depression-Related Behaviors in Adulthood. Neuroscience 2015, 299, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-L.; Lin, S.-Y.; Wang, S. Prenatal Lipopolysaccharide Exposure Increases Anxiety-like Behaviors and Enhances Stress-Induced Corticosterone Responses in Adult Rats. Brain Behav. Immun. 2012, 26, 459–468. [Google Scholar] [CrossRef]
- Foley, K.A.; Ossenkopp, K.-P.; Kavaliers, M.; MacFabe, D.F. Pre- and Neonatal Exposure to Lipopolysaccharide or the Enteric Metabolite, Propionic Acid, Alters Development and Behavior in Adolescent Rats in a Sexually Dimorphic Manner. PLoS ONE 2014, 9, e87072. [Google Scholar] [CrossRef] [Green Version]
- Lysaker, P.H.; Salyers, M.P. Anxiety Symptoms in Schizophrenia Spectrum Disorders: Associations with Social Function, Positive and Negative Symptoms, Hope and Trauma History. Acta Psychiatr. Scand. 2007, 116, 290–298. [Google Scholar] [CrossRef]
- White, S.W.; Oswald, D.; Ollendick, T.; Scahill, L. Anxiety in Children and Adolescents with Autism Spectrum Disorders. Clin. Psychol. Rev. 2009, 29, 216–229. [Google Scholar] [CrossRef] [Green Version]
- Bubenikova-Valesova, V.; Horacek, J.; Vrajova, M.; Hoschl, C. Models of Schizophrenia in Humans and Animals Based on Inhibition of NMDA Receptors. Neurosci. Biobehav. Rev. 2008, 32, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, L.; Fraga, D.B.; De Luca, R.D.; Canever, L.; Ghedim, F.V.; Matos, M.P.P.; Streck, E.L.; Quevedo, J.; Zugno, A.I. Behavioral Changes and Mitochondrial Dysfunction in a Rat Model of Schizophrenia Induced by Ketamine. Metab. Brain Dis. 2011, 26, 69–77. [Google Scholar] [CrossRef]
- Shirai, Y.; Fujita, Y.; Hashimoto, K. Effects of the Antioxidant Sulforaphane on Hyperlocomotion and Prepulse Inhibition Deficits in Mice after Phencyclidine Administration. Clin. Psychopharmacol. Neurosci. 2012, 10, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Hu, G.; Li, M. Repeated Antipsychotic Treatment Progressively Potentiates Inhibition on Phencyclidine-Induced Hyperlocomotion, but Attenuates Inhibition on Amphetamine-Induced Hyperlocomotion: Relevance to Animal Models of Antipsychotic Drugs. Eur. J. Pharmacol. 2009, 602, 334–342. [Google Scholar] [CrossRef] [Green Version]
- Vojtechova, I.; Petrasek, T.; Hatalova, H.; Pistikova, A.; Vales, K.; Stuchlik, A. Dizocilpine (MK-801) Impairs Learning in the Active Place Avoidance Task but Has No Effect on the Performance during Task/Context Alternation. Behav. Brain Res. 2016, 305, 247–257. [Google Scholar] [CrossRef]
- Tejkalova, H.; Jelinek, F.; Klaschka, J.; Stastny, F. Effect of perinatal inflammatory reaction on the induction of psychotic-like behaviour: Experimental study. Psychiatrie 2007, 11, 12–15. [Google Scholar]
- Stigger, F.; Lovatel, G.; Marques, M.; Bertoldi, K.; Moysés, F.; Elsner, V.; Siqueira, I.R.; Achaval, M.; Marcuzzo, S. Inflammatory Response and Oxidative Stress in Developing Rat Brain and Its Consequences on Motor Behavior Following Maternal Administration of LPS and Perinatal Anoxia. Int. J. Dev. Neurosci. 2013, 31, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Foussias, G.; Remington, G. Negative Symptoms in Schizophrenia: Avolition and Occam’s Razor. Schizophr. Bull. 2010, 36, 359–369. [Google Scholar] [CrossRef] [Green Version]
- Foley, K.A.; MacFabe, D.F.; Kavaliers, M.; Ossenkopp, K.-P. Sexually Dimorphic Effects of Prenatal Exposure to Lipopolysaccharide, and Prenatal and Postnatal Exposure to Propionic Acid, on Acoustic Startle Response and Prepulse Inhibition in Adolescent Rats: Relevance to Autism Spectrum Disorders. Behav. Brain Res. 2015, 278, 244–256. [Google Scholar] [CrossRef]
- Fortier, M.-E.; Luheshi, G.N.; Boksa, P. Effects of Prenatal Infection on Prepulse Inhibition in the Rat Depend on the Nature of the Infectious Agent and the Stage of Pregnancy. Behav. Brain Res. 2007, 181, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Braff, D.L.; Geyer, M.A.; Light, G.A.; Sprock, J.; Perry, W.; Cadenhead, K.S.; Swerdlow, N.R. Impact of Prepulse Characteristics on the Detection of Sensorimotor Gating Deficits in Schizophrenia. Schizophr. Res. 2001, 49, 171–178. [Google Scholar] [CrossRef]
- Geyer, M.A.; Krebs-Thomson, K.; Braff, D.L.; Swerdlow, N.R. Pharmacological Studies of Prepulse Inhibition Models of Sensorimotor Gating Deficits in Schizophrenia: A Decade in Review. Psychopharmacology 2001, 156, 117–154. [Google Scholar] [CrossRef]
- Kohl, S.; Heekeren, K.; Klosterkötter, J.; Kuhn, J. Prepulse Inhibition in Psychiatric Disorders—Apart from Schizophrenia. J. Psychiatr. Res. 2013, 47, 445–452. [Google Scholar] [CrossRef]
- Madsen, G.F.; Bilenberg, N.; Cantio, C.; Oranje, B. Increased Prepulse Inhibition and Sensitization of the Startle Reflex in Autistic Children: Sensorimotor Gating in Autistic Children. Autism Res. 2014, 7, 94–103. [Google Scholar] [CrossRef]
- Perry, W.; Minassian, A.; Lopez, B.; Maron, L.; Lincoln, A. Sensorimotor Gating Deficits in Adults with Autism. Biol. Psychiatry 2007, 61, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Kohl, S.; Wolters, C.; Gruendler, T.O.J.; Vogeley, K.; Klosterkötter, J.; Kuhn, J. Prepulse Inhibition of the Acoustic Startle Reflex in High Functioning Autism. PLoS ONE 2014, 9, e92372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, U.; Feldon, J.; Dammann, O. Schizophrenia and Autism: Both Shared and Disorder-Specific Pathogenesis via Perinatal Inflammation? Pediatr. Res. 2011, 69, 26R–33R. [Google Scholar] [CrossRef] [Green Version]
- Marcondes, F.K.; Bianchi, F.J.; Tanno, A.P. Determination of the Estrous Cycle Phases of Rats: Some Helpful Considerations. Braz. J. Biol. 2002, 62, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Academic Press: Boston, MA, USA; Elsevier: Amsterdam, The Netherlands, 2007; ISBN 978-0-12-547612-6. [Google Scholar]
- Wallace, C.S.; Reitzenstein, J.; Withers, G.S. Diminished Experience-Dependent Neuroanatomical Plasticity: Evidence for an Improved Biomarker of Subtle Neurotoxic Damage to the Developing Rat Brain. Environ. Health Perspect. 2003, 111, 1294–1298. [Google Scholar] [CrossRef] [Green Version]
- Yan, E.B.; Hellewell, S.C.; Bellander, B.-M.; Agyapomaa, D.A.; Morganti-Kossmann, M.C. Post-Traumatic Hypoxia Exacerbates Neurological Deficit, Neuroinflammation and Cerebral Metabolism in Rats with Diffuse Traumatic Brain Injury. J. Neuroinflammation 2011, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Bures, J.; Fenton, A.A.; Kaminsky, Y.; Zinyuk, L. Place Cells and Place Navigation. Proc. Natl. Acad. Sci. USA 1997, 94, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Fenton, A.A.; Wesierska, M.; Kaminsky, Y.; Bures, J. Both Here and There: Simultaneous Expression of Autonomous Spatial Memories in Rats. Proc. Natl. Acad. Sci. USA 1998, 95, 11493–11498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuchlík, A.; Petrásek, T.; Prokopová, I.; Holubová, K.; Hatalová, H.; Valeš, K.; Kubík, Š.; Dockery, C.; Wesierska, M. Place Avoidance Tasks as Tools in the Behavioral Neuroscience of Learning and Memory. Physiol. Res. 2013, 62, S1–S19. [Google Scholar] [CrossRef] [PubMed]
- Friard, O.; Gamba, M. BORIS: A Free, Versatile Open-Source Event-Logging Software for Video/Audio Coding and Live Observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Burke, A.W.; Broadhurst, P.L. Behavioural Correlates of the Oestrous Cycle in the Rat. Nature 1966, 209, 223–224. [Google Scholar] [CrossRef]
- Feltenstein, M.W.; See, R.E. Plasma Progesterone Levels and Cocaine-Seeking in Freely Cycling Female Rats across the Estrous Cycle. Drug Alcohol Depend. 2007, 89, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousset, C.I.; Kassem, J.; Aubert, A.; Planchenault, D.; Gressens, P.; Chalon, S.; Belzung, C.; Saliba, E. Maternal Exposure to Lipopolysaccharide Leads to Transient Motor Dysfunction in Neonatal Rats. Dev. Neurosci. 2013, 35, 172–181. [Google Scholar] [CrossRef] [PubMed]







| Group | Number of Animals and Litters Per Group (n) | ||
|---|---|---|---|
| Brain Analysis (P28) | Behavior Analysis (P45, P90) | ||
| Brain Morphology | Immunohistochemistry | ||
| saline males | n = 6 (4 litters) | n = 5 (4 litters) | n = 15 (5 litters) |
| LPS males | n = 10 (7 litters) | n = 5 (5 litters) | n = 14 (7 litters) |
| saline females | n = 10 (4 litters) | n = 5 (4 litters) | n = 13 (5 litters) |
| LPS females | n = 10 (7 litters) | n = 5 (5 litters) | n = 14 (7 litters) |
| Training | Test | Training | Test | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Trial | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Width | 4.5 cm | 4.5 cm | 4.5 cm | 4.5 cm | 2.2 cm | 2.2 cm | 2.2 cm | 2.2 cm | 2.2 cm |
| Length | 0.5 m | 1 m | 2 m | 2 m | 1 m | 2 m | 2 m | 2 m | 2 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vojtechova, I.; Maleninska, K.; Kutna, V.; Klovrza, O.; Tuckova, K.; Petrasek, T.; Stuchlik, A. Behavioral Alterations and Decreased Number of Parvalbumin-Positive Interneurons in Wistar Rats after Maternal Immune Activation by Lipopolysaccharide: Sex Matters. Int. J. Mol. Sci. 2021, 22, 3274. https://doi.org/10.3390/ijms22063274
Vojtechova I, Maleninska K, Kutna V, Klovrza O, Tuckova K, Petrasek T, Stuchlik A. Behavioral Alterations and Decreased Number of Parvalbumin-Positive Interneurons in Wistar Rats after Maternal Immune Activation by Lipopolysaccharide: Sex Matters. International Journal of Molecular Sciences. 2021; 22(6):3274. https://doi.org/10.3390/ijms22063274
Chicago/Turabian StyleVojtechova, Iveta, Kristyna Maleninska, Viera Kutna, Ondrej Klovrza, Klara Tuckova, Tomas Petrasek, and Ales Stuchlik. 2021. "Behavioral Alterations and Decreased Number of Parvalbumin-Positive Interneurons in Wistar Rats after Maternal Immune Activation by Lipopolysaccharide: Sex Matters" International Journal of Molecular Sciences 22, no. 6: 3274. https://doi.org/10.3390/ijms22063274