
G-Quadruplex Structures Colocalize with Transcription Factories and Nuclear Speckles Surrounded by Acetylated and Dimethylated Histones H3


Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Cultivation and Treatment
2.2. Cell Transfection
2.3. Immunofluorescence Staining
2.4. Laser Scanning Confocal Microscopy
2.5. Statistical Analyses and Quantification of Fluorescence Intensity
3. Results
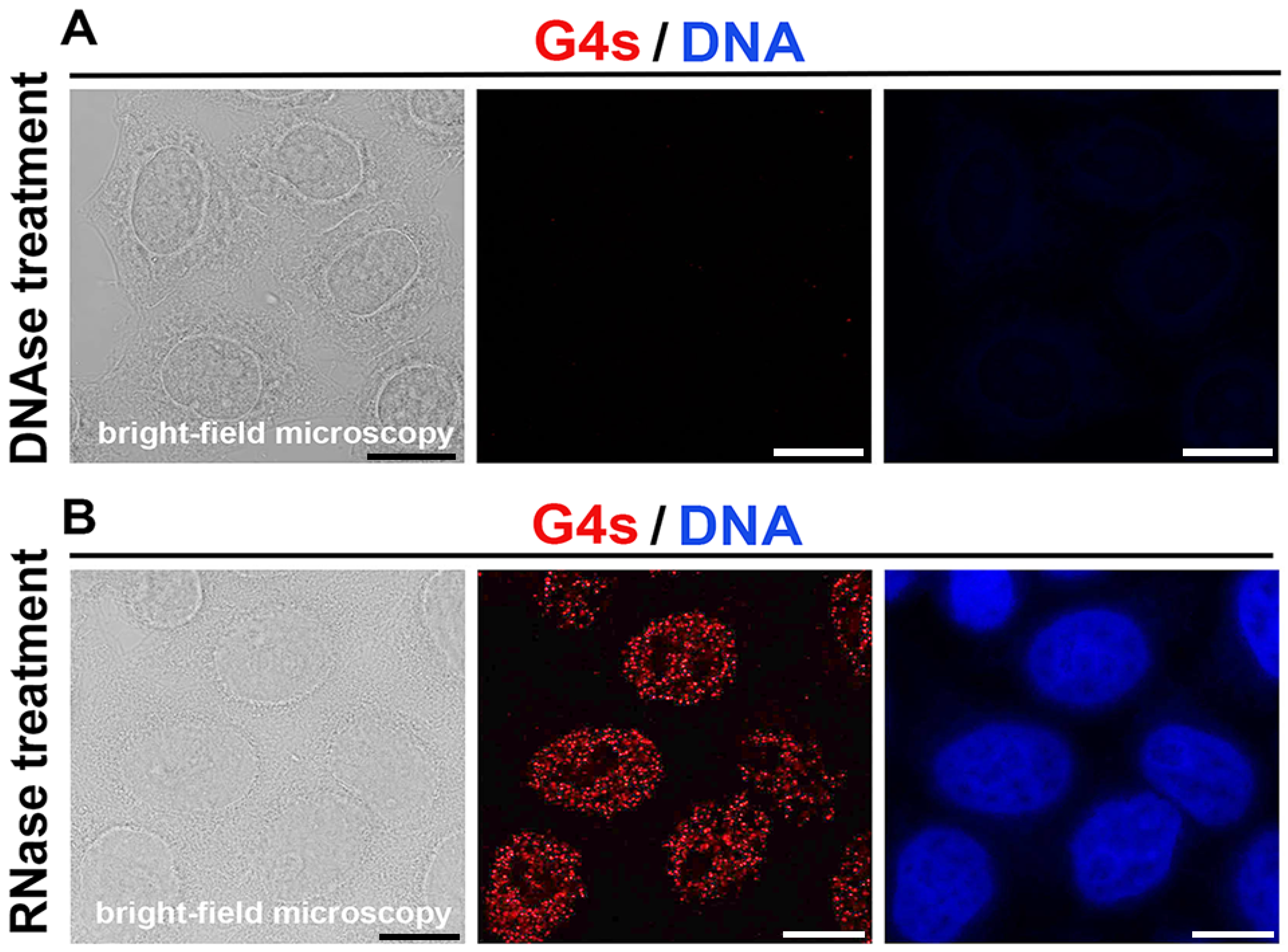
3.1. Immunodetection of G-Quadruplex Structures in DNA
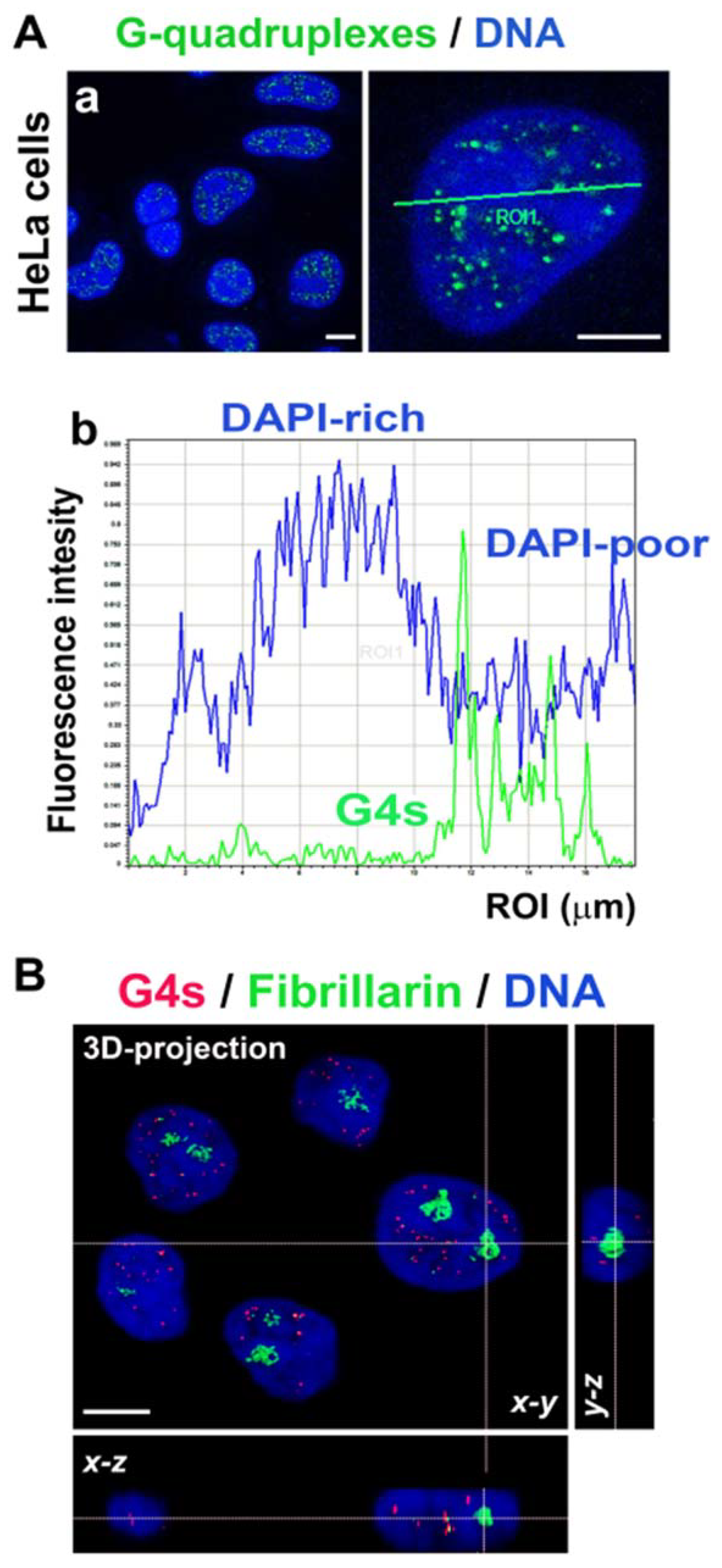
3.2. Nuclear Localization of G-Quadruplex Structures Is in GC-Rich Chromatin and Outside the Fibrillarin-Positive Region of Nucleoli
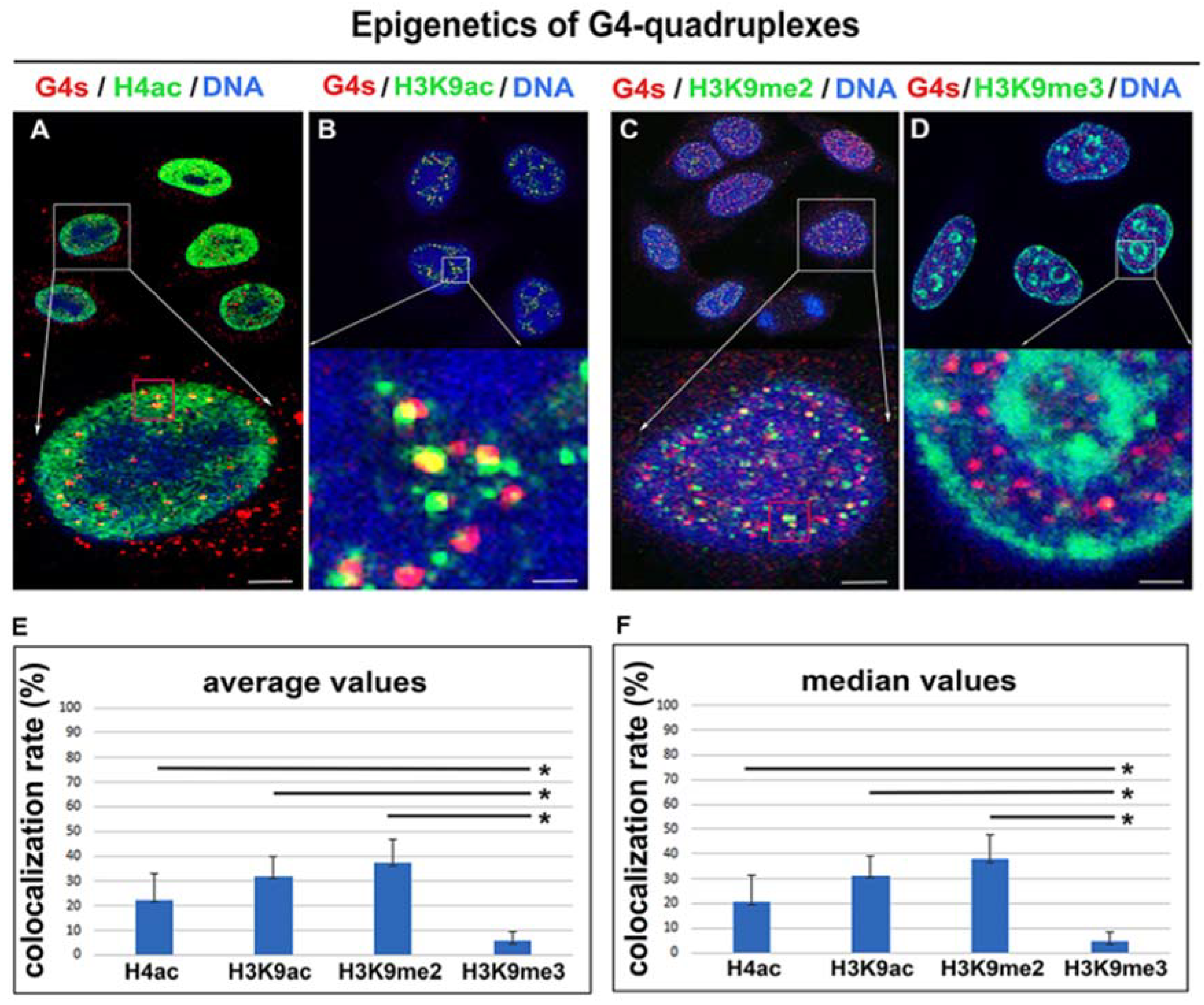
3.3. Epigenetic Landscape of G-Quadruplex Structures
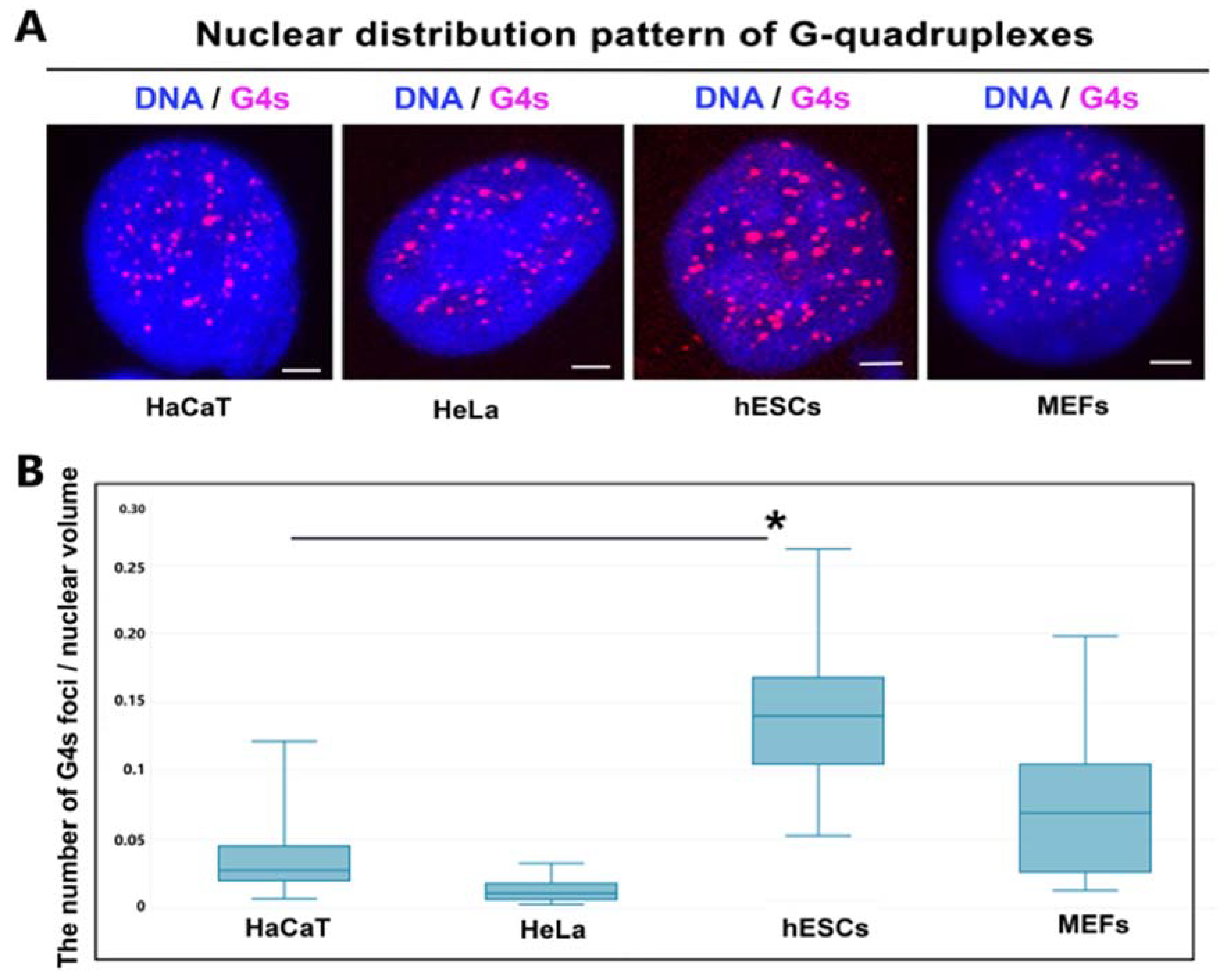
3.4. Nuclear Distribution Pattern of G-Quadruplexes in Distinct Cell Types
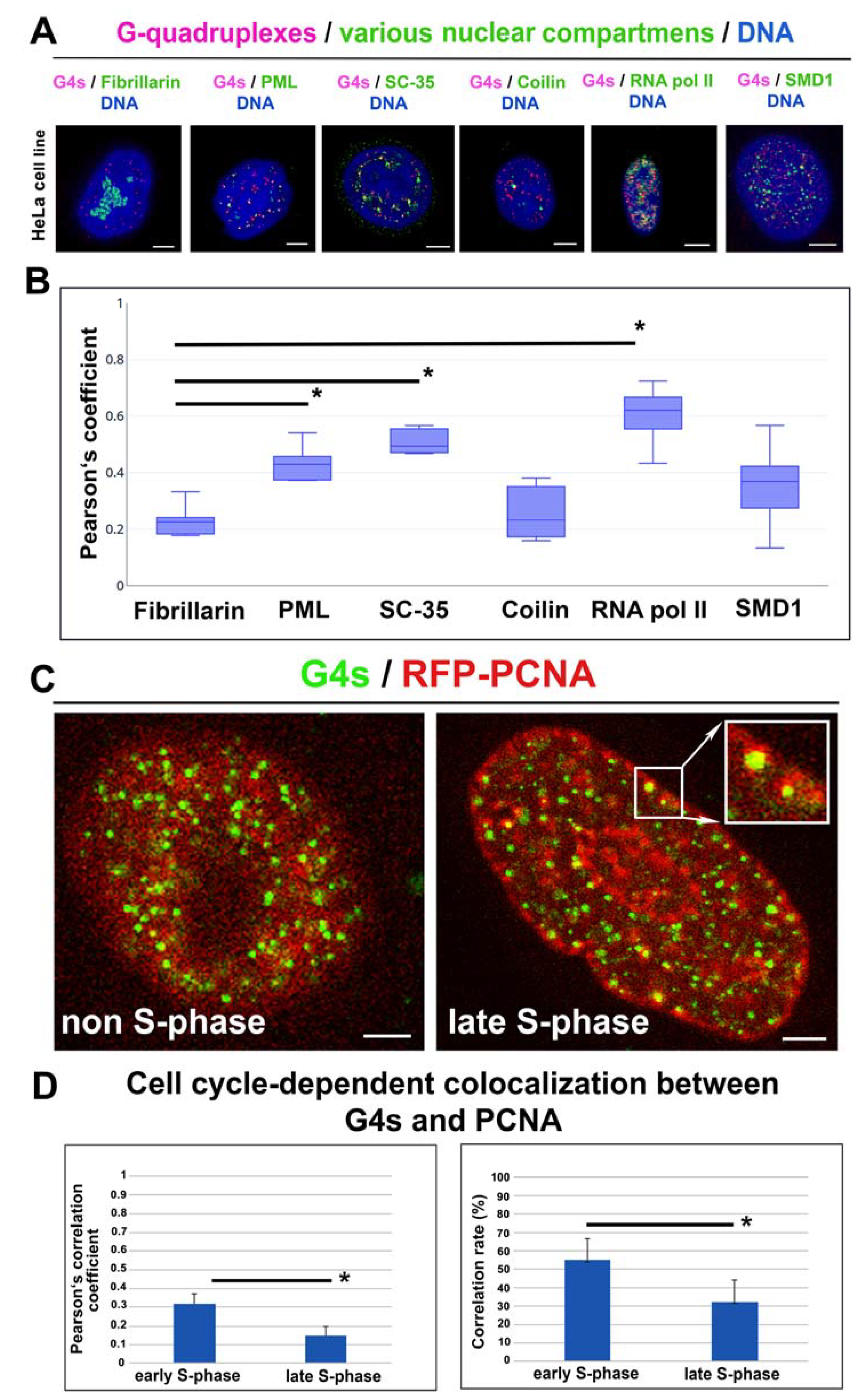
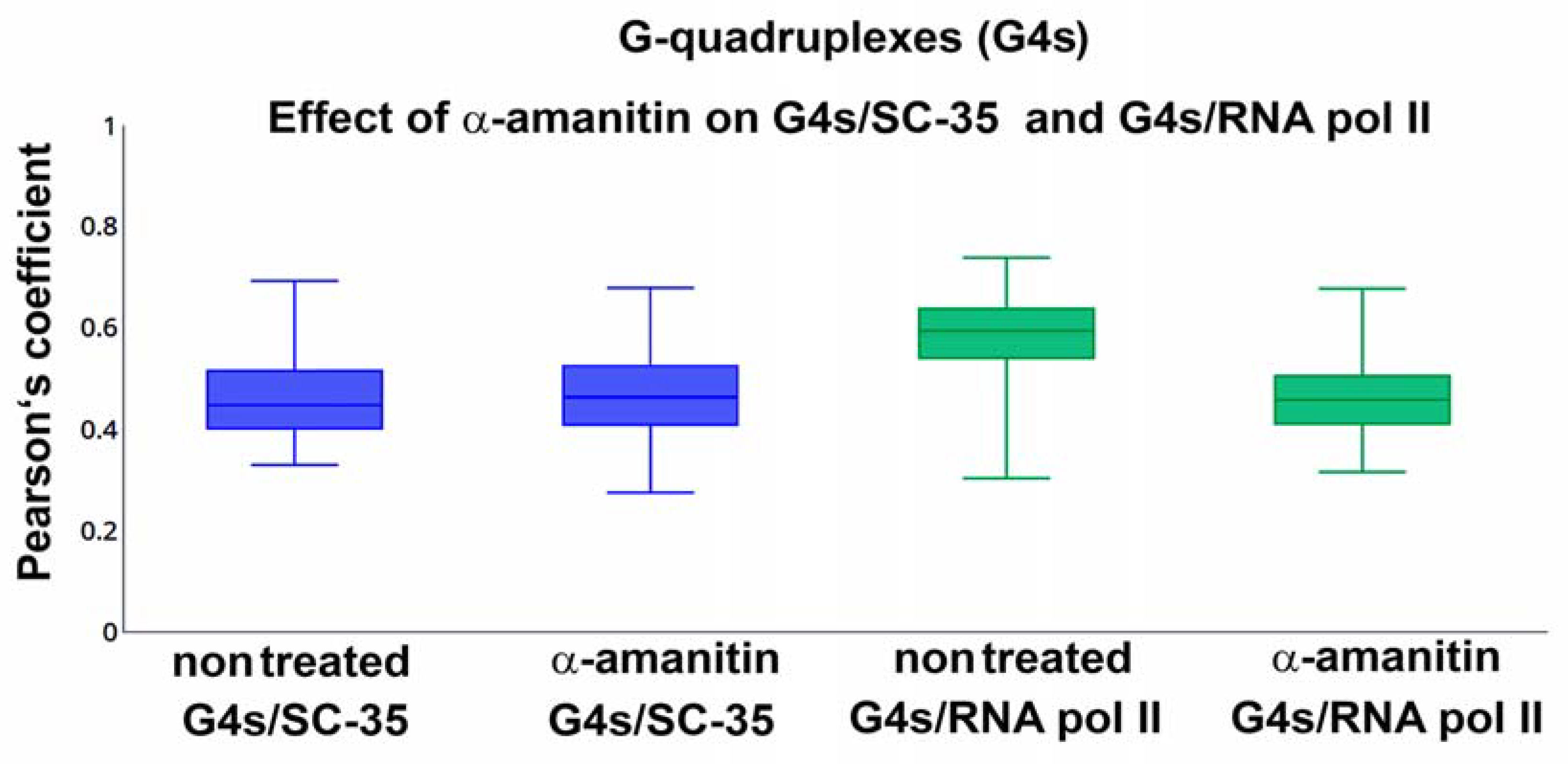
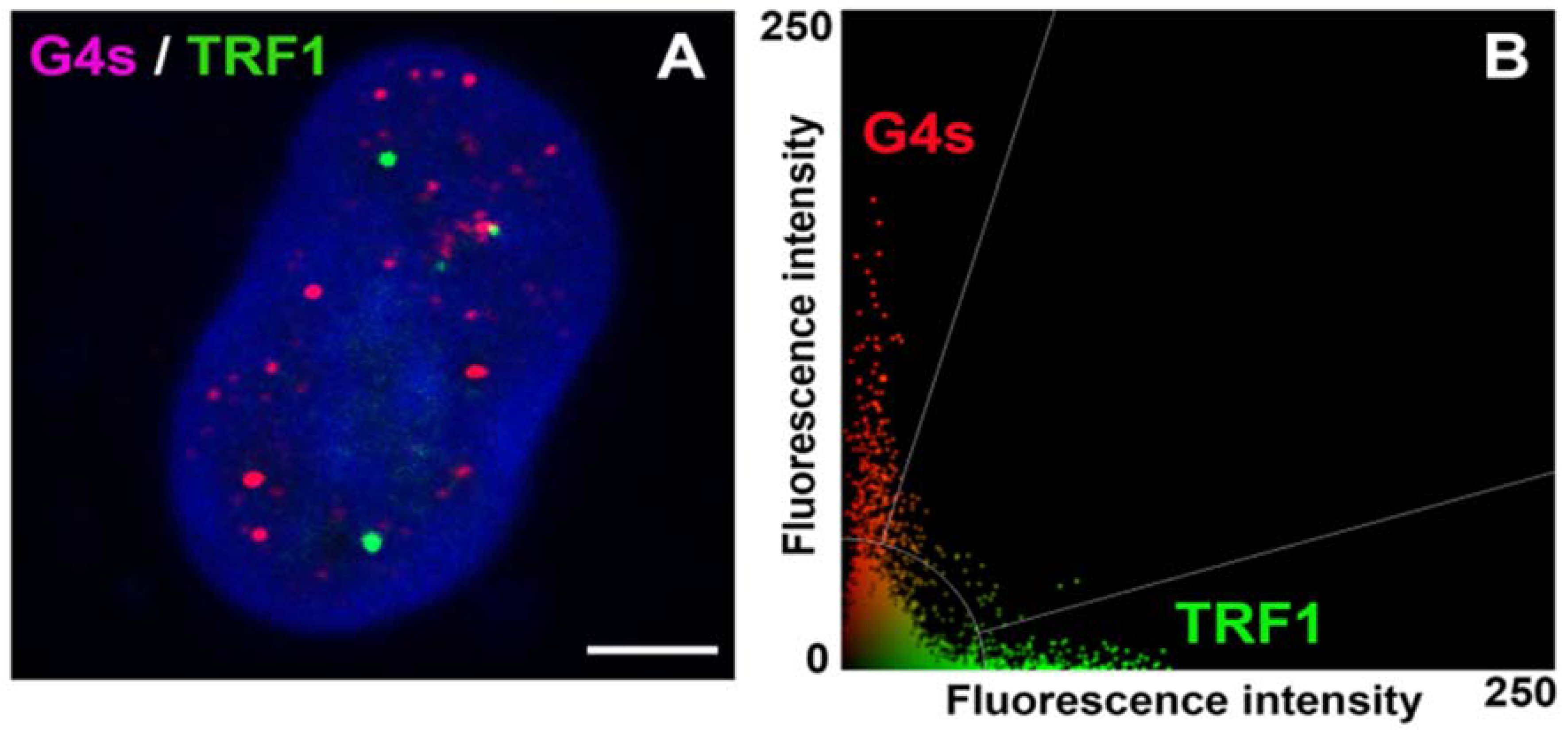
3.5. The Link of G-Quadruplex Structures to Nucleoli, PML Bodies, Cajal Bodies, Transcription Factories, Nuclear Speckles, and Replication Foci: A Degree of Colocalization
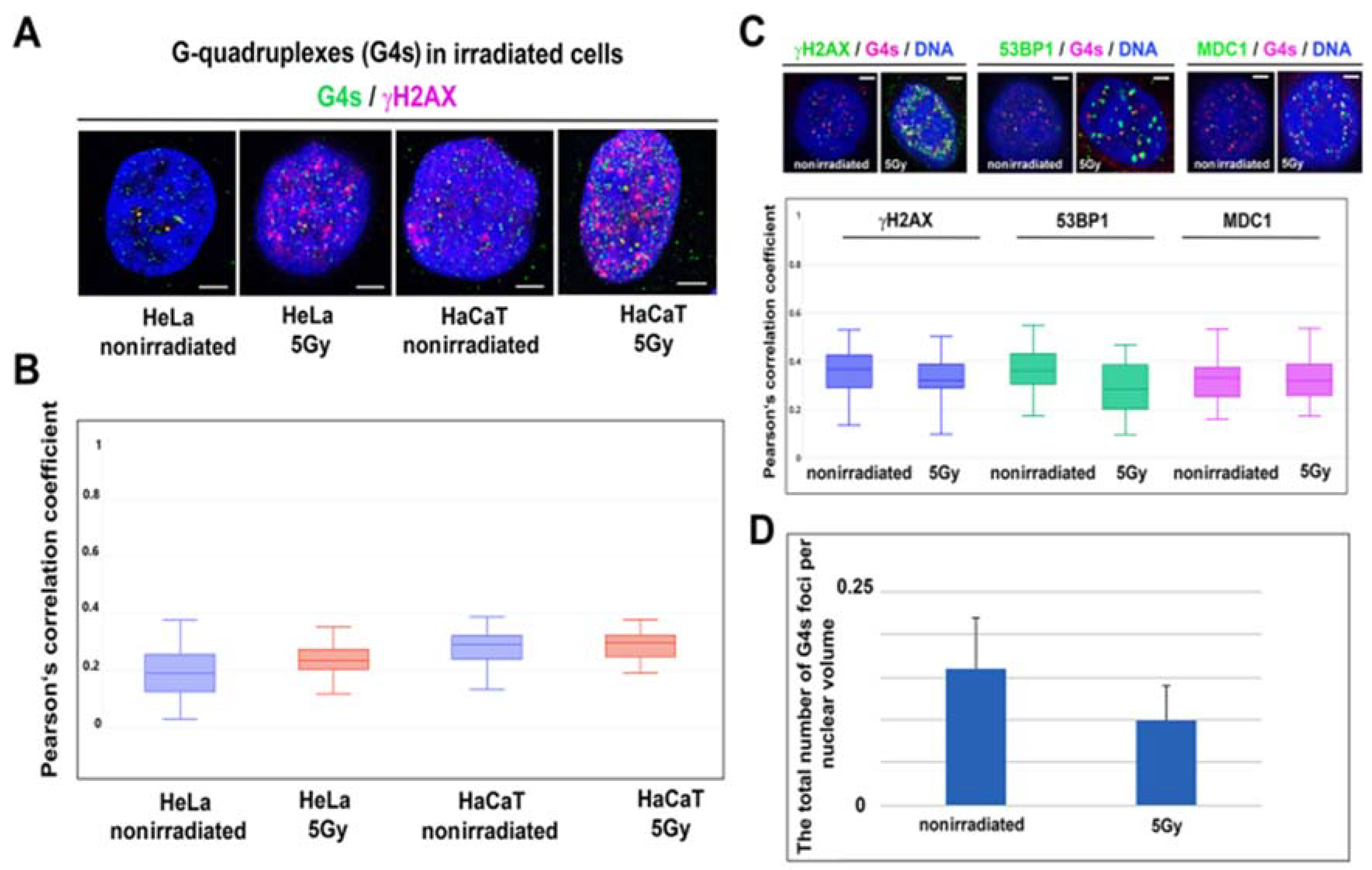
3.6. G-Quadruplex Structures and DNA Repair
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Bcl-2 | B-cell lymphoma 2 |
| CBs | Cajal bodies |
| c-myc | Avian myelocytomatosis virus oncogene cellular homolog |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DNMT1 | DNA methyltransferase 1 |
| ES cells | Embryonic stem cells |
| EXO1 | Exonuclease 1 |
| FEN1 | Flap endonuclease 1 |
| FCS | Fetal calf serum |
| GFP | Green fluorescein protein |
| G4 | G-quadruplexes |
| HaCaT | Human keratinocytes |
| HeLa | Cervical carcinoma cells |
| hESCs | Human embryonic stem cells |
| hTR | The RNA template of telomerase |
| H3K9 (ac, me) | Epigenetic modification of protein histone H3 on lyzine (K) 9 |
| H4 | Protein histone H4 |
| HRR | Homologous recombination repair |
| IgH | Immunoglobulin heavy chain |
| IRIF | γ-irradiation-induced foci |
| MDC1 | Mediator of DNA damage checkpoint protein 1 |
| MEFs | Mouse embryonic fibroblasts |
| MeOH | Methanol |
| NHEJ | Nonhomologous end joining |
| Oct4 | Octamer-binding transcription factor 4 |
| PBS | Phosphate-buffered saline |
| PCNA | Proliferating cell nuclear antigen |
| PFA | Paraformaldehyde |
| PML | Promyelocytic leukemia |
| RFP | Red fluorescein protein |
| RNAP II | RNA polymerase II |
| RNAP II- phS5 | Phosphorylated form of RNAP II |
| RT | Room temperature |
| SC35 | Nuclear speckle |
| SMD1 | Small nuclear ribonucleoprotein |
| ssDNA | Single stranged stranded DNA |
| TRF-1 | Telomeric-repeat binding factor 1 |
| WLL | White light laser |
| γH2AX | Phosphorylated histone H2AX |
| 3D | Three-dimensional |
| 53BP1 | tumor suppressor p53-binding protein 1 |
References
- Gellert, M.; Lipsett, M.N.; Davies, D.R. Helix Formation by Guanylic Acid. Proc. Natl. Acad. Sci. USA 1962, 48, 2013–2018. [Google Scholar] [CrossRef] [Green Version]
- Sen, D.; Gilbert, W. Formation of Parallel Four-Stranded Complexes by Guanine-Rich Motifs in DNA and Its Implications for Meiosis. Nature 1988, 334, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, Topology and Structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moye, A.L.; Porter, K.C.; Cohen, S.B.; Phan, T.; Zyner, K.G.; Sasaki, N.; Lovrecz, G.O.; Beck, J.L.; Bryan, T.M. Telomeric G-Quadruplexes Are a Substrate and Site of Localization for Human Telomerase. Nat. Commun. 2015, 6, 7643. [Google Scholar] [CrossRef] [Green Version]
- Fajkus, P.; Peška, V.; Závodník, M.; Fojtová, M.; Fulnečková, J.; Dobias, Š.; Kilar, A.; Dvořáčková, M.; Zachová, D.; Nečasová, I.; et al. Telomerase RNAs in Land Plants. Nucleic Acids Res. 2019, 47, 9842–9856. [Google Scholar] [CrossRef] [Green Version]
- De Lange, T. Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.S.; Chen, Q.; Yatsunyk, L.A.; Nicoludis, J.M.; Garcia, M.S.; Kranaster, R.; Balasubramanian, S.; Monchaud, D.; Teulade-Fichou, M.-P.; Abramowitz, L.; et al. Rudimentary G-Quadruplex-Based Telomere Capping in Saccharomyces Cerevisiae. Nat. Struct. Mol. Biol. 2011, 18, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative Visualization of DNA G-Quadruplex Structures in Human Cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-Quadruplexes in Gene Promoters: A Novel Anticancer Strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Hurley, L. Structure of the Biologically Relevant G-Quadruplex in the c-MYC Promoter. Nucleosides Nucleotides Nucleic Acids 2006, 25, 951–968. [Google Scholar] [CrossRef]
- Brooks, T.A.; Hurley, L.H. Targeting MYC Expression through G-Quadruplexes. Genes Cancer 2010, 1, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Haluska, F.G.; Finver, S.; Tsujimoto, Y.; Croce, C.M. The t(8; 14) Chromosomal Translocation Occurring in B-Cell Malignancies Results from Mistakes in V-D-J Joining. Nature 1986, 324, 158–161. [Google Scholar] [CrossRef]
- Raghavan, S.C.; Swanson, P.C.; Wu, X.; Hsieh, C.L.; Lieber, M.R. A Non-B-DNA Structure at the Bcl-2 Major Breakpoint Region Is Cleaved by the RAG Complex. Nature 2004, 428, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Galteland, E.; Sivertsen, E.A.; Svendsrud, D.H.; Smedshammer, L.; Kresse, S.H.; Meza-Zepeda, L.A.; Myklebost, O.; Suo, Z.; Mu, D.; Deangelis, P.M.; et al. Translocation t(14;18) and Gain of Chromosome 18/BCL2: Effects on BCL2 Expression and Apoptosis in B-Cell Non-Hodgkin’s Lymphomas. Leukemia 2005, 19, 2313–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De, S.; Michor, F. DNA Secondary Structures and Epigenetic Determinants of Cancer Genome Evolution. Nat. Struct. Mol. Biol. 2011, 18, 950–955. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.Q.; Ghanbarian, A.T.; Spiegel, J.; Martínez Cuesta, S.; Beraldi, D.; Di Antonio, M.; Marsico, G.; Hänsel-Hertsch, R.; Tannahill, D.; Balasubramanian, S. DNA G-Quadruplex Structures Mold the DNA Methylome. Nat. Struct. Mol. Biol. 2018, 25, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Renčiuk, D.; Ryneš, J.; Kejnovská, I.; Foldynová-Trantírková, S.; Andäng, M.; Trantírek, L.; Vorlíčková, M. G-Quadruplex Formation in the Oct4 Promoter Positively Regulates Oct4 Expression. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.; Jurk, D.; Marques, F.D.M.; Correia-Melo, C.; Hardy, T.; Gackowska, A.; Anderson, R.; Taschuk, M.; Mann, J.; Passos, J.F. Telomeres Are Favoured Targets of a Persistent DNA Damage Response in Ageing and Stress-Induced Senescence. Nat. Commun. 2012, 3, 708. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gilbert, W. The Yeast KEM1 Gene Encodes a Nuclease Specific for G4 Tetraplex DNA: Implication of in Vivo Functions for This Novel DNA Structure. Cell 1994, 77, 1083–1092. [Google Scholar] [CrossRef]
- Vallur, A.C.; Maizels, N. Activities of Human Exonuclease 1 That Promote Cleavage of Transcribed Immunoglobulin Switch Regions. Proc. Natl. Acad. Sci. USA 2008, 105, 16508–16512. [Google Scholar] [CrossRef] [Green Version]
- Saharia, A.; Guittat, L.; Crocker, S.; Lim, A.; Steffen, M.; Kulkarni, S.; Stewart, S.A. Flap Endonuclease 1 Contributes to Telomere Stability. Curr. Biol. 2008, 18, 496–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, R.; Miller, K.M.; Forment, J.V.; Bradshaw, C.R.; Nikan, M.; Britton, S.; Oelschlaegel, T.; Xhemalce, B.; Balasubramanian, S.; Jackson, S.P. Small-Molecule-Induced DNA Damage Identifies Alternative DNA Structures in Human Genes. Nat. Chem. Biol. 2012, 8, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lago, S.; Tosoni, E.; Nadai, M.; Palumbo, M.; Richter, S.N. The Cellular Protein Nucleolin Preferentially Binds Long-Looped G-Quadruplex Nucleic Acids. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1371–1381. [Google Scholar] [CrossRef] [PubMed]
- Bártová, E.; Šustáčková, G.; Stixová, L.; Kozubek, S.; Legartová, S.; Foltánková, V. Recruitment of Oct4 Protein to UV-Damaged Chromatin in Embryonic Stem Cells. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stixová, L.; Matula, P.; Kozubek, S.; Gombitová, A.; Cmarko, D.; Raška, I.; Bártová, E. Trajectories and Nuclear Arrangement of PML Bodies Are Influenced by A-Type Lamin Deficiency. Biol. Cell 2012, 104, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Henderson, A.; Wu, Y.; Huang, Y.C.; Chavez, E.A.; Platt, J.; Johnson, F.B.; Brosh, R.M.; Sen, D.; Lansdorp, P.M.; Lansdorp, P.M. Detection of G-Quadruplex DNA in Mammalian Cells. Nucleic Acids Res. 2014, 42, 860–869. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-Z.; Jenjaroenpun, P.; Wongsurawat, T.; Byrum, S.D.; Shponka, V.; Tannahill, D.; Chavez, E.A.; Hung, S.S.; Steidl, C.; Balasubramanian, S.; et al. Activation-Induced Cytidine Deaminase Localizes to G-Quadruplex Motifs at Mutation Hotspots in Lymphoma. NAR Cancer 2020, 2. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.K.; Zybailov, B.L.; Maddukuri, L.; Gao, J.; Marecki, J.C.; Jaiswal, M.; Bell, M.R.; Griffin, W.C.; Reed, M.R.; Chib, S.; et al. Evidence That G-Quadruplex DNA Accumulates in the Cytoplasm and Participates in Stress Granule Assembly in Response to Oxidative Stress. J. Biol. Chem. 2016, 291, 18041–18057. [Google Scholar] [CrossRef] [Green Version]
- Canesin, G.; Di Ruscio, A.; Li, M.; Ummarino, S.; Hedblom, A.; Choudhury, R.; Krzyzanowska, A.; Csizmadia, E.; Palominos, M.; Stiehm, A.; et al. Scavenging of Labile Heme by Hemopexin Is a Key Checkpoint in Cancer Growth and Metastases. Cell Rep. 2020, 32, 108181. [Google Scholar] [CrossRef]
- Biffi, G.; Di Antonio, M.; Tannahill, D.; Balasubramanian, S. Visualization and Selective Chemical Targeting of RNA G-Quadruplex Structures in the Cytoplasm of Human Cells. Nat. Chem. 2014, 6, 75–80. [Google Scholar] [CrossRef]
- David, A.P.; Pipier, A.; Pascutti, F.; Binolfi, A.; Weiner, A.M.J.; Challier, E.; Heckel, S.; Calsou, P.; Gomez, D.; Calcaterra, N.B.; et al. CNBP Controls Transcription by Unfolding DNA G-Quadruplex Structures. Nucleic Acids Res. 2019, 47, 7901–7913. [Google Scholar] [CrossRef]
- Bharti, S.K.; Sommers, J.A.; Awate, S.; Bellani, M.A.; Khan, I.; Bradley, L.; King, G.A.; Seol, Y.; Vidhyasagar, V.; Wu, Y.; et al. A Minimal Threshold of FANCJ Helicase Activity Is Required for Its Response to Replication Stress or Double-Strand Break Repair. Nucleic Acids Res. 2018, 46, 6238–6256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, R.F.; Moshkin, Y.M.; Mouton, S.; Grzeschik, N.A.; Kalicharan, R.D.; Kuipers, J.; Wolters, A.H.G.; Nishida, K.; Romashchenko, A.V.; Postberg, J.; et al. Guanine Quadruplex Structures Localize to Heterochromatin. Nucleic Acids Res. 2016, 44, 152–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemier, H.G.; Paeschke, K.; Lansdorp, P.M. Guanine Quadruplex Monoclonal Antibody 1H6 Cross-Reacts with Restrained Thymidine-Rich Single Stranded DNA. Nucleic Acids Res. 2017, 45, 5913–5919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, S.; Tillo, D.; Boer, R.E.; Assad, N.; Barshai, M.; Wu, G.; Orenstein, Y.; Yang, D.; Schneekloth, J.S.; Vinson, C. Custom DNA Microarrays Reveal Diverse Binding Preferences of Proteins and Small Molecules to Thousands of G-Quadruplexes. ACS Chem. Biol. 2020, 15, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Murat, P.; Balasubramanian, S. Existence and Consequences of G-Quadruplex Structures in DNA. Curr. Opin. Genet. Dev. 2014, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.M.; Lyons, S.M.; Ivanov, P. RNA G-Quadruplexes in Biology: Principles and Molecular Mechanisms. J. Mol. Biol. 2017, 2127–2147. [Google Scholar] [CrossRef] [PubMed]
- Paeschke, K.; Capra, J.A.; Zakian, V.A. DNA Replication through G-Quadruplex Motifs Is Promoted by the Saccharomyces Cerevisiae Pif1 DNA Helicase. Cell 2011, 145, 678–691. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.; Lipps, H.J. G-Quadruplexes and Their Regulatory Roles in Biology. Nucleic Acids Res. 2015, 43, 8627–8637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benetti, R.; García-Cao, M.; Blasco, M.A. Telomere Length Regulates the Epigenetic Status of Mammalian Telomeres and Subtelomeres. Nat. Genet. 2007, 39, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Takahama, K.; Takada, A.; Tada, S.; Shimizu, M.; Sayama, K.; Kurokawa, R.; Oyoshi, T. Regulation of Telomere Length by G-Quadruplex Telomere DNA- and TERRA-Binding Protein TLS/FUS. Chem. Biol. 2013, 20, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyoshi, T.; Masuzawa, T. Modulation of Histone Modifications and G-Quadruplex Structures by G-Quadruplex-Binding Proteins. Biochem. Biophys. Res. Commun. 2020, 531, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct Evidence for a G-Quadruplex in a Promoter Region and Its Targeting with a Small Molecule to Repress c-MYC Transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonsson, T.; Henriksson, M. C-Myc Suppression in Burkitt’s Lymphoma Cells. Biochem. Biophys. Res. Commun. 2002, 290, 11–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, E.Y.N.; Beraldi, D.; Tannahill, D.; Balasubramanian, S. G-Quadruplex Structures Are Stable and Detectable in Human Genomic DNA. Nat. Commun. 2013, 4, 1796. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Komiyama, M. Evidence for G-Quadruplex DNA in Human Cells. ChemBioChem 2013, 14, 927–928. [Google Scholar] [CrossRef]
- Bryan, T.M. G-Quadruplexes at Telomeres: Friend or Foe? Molecules 2020, 25, 3686. [Google Scholar] [CrossRef]
- Saha, A.; Duchambon, P.; Masson, V.; Loew, D.; Bombard, S.; Teulade-Fichou, M.P. Nucleolin Discriminates Drastically between Long-Loop and Short-Loop Quadruplexes. Biochemistry 2020. [Google Scholar] [CrossRef]
- Brooks, T.A.; Hurley, L.H. The Role of Supercoiling in Transcriptional Control of MYC and Its Importance in Molecular Therapeutics. Nat. Rev. Cancer 2009, 849–861. [Google Scholar] [CrossRef]
- González, V.; Guo, K.; Hurley, L.; Sun, D. Identification and Characterization of Nucleolin as a C-Myc G-Quadruplex-Binding Protein. J. Biol. Chem. 2009, 284, 23622–23635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, V.; Hurley, L.H. The C-Terminus of Nucleolin Promotes the Formation of the c-MYC G-Quadruplex and Inhibits c-MYC Promoter Activity. Biochemistry 2010, 49, 9706–9714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spotheim-Maurizot, M.; Davídková, M. Radiation Damage to DNA in DNA-Protein Complexes. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2011, 711, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Vartak, S.V.; Dahal, S.; Kumari, S.; Desai, S.S.; Gopalakrishnan, V.; Choudhary, B.; Raghavan, S.C. G-Quadruplex Structures Contribute to Differential Radiosensitivity of the Human Genome. iScience 2019, 21, 288–307. [Google Scholar] [CrossRef] [Green Version]
- Williamson, A.K.; Zhu, Z.; Yuan, Z.M. Epigenetic Mechanisms behind Cellular Sensitivity to DNA Damage. Cell Stress 2018, 2, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Jakob, B.; Splinter, J.; Conrad, S.; Voss, K.O.; Zink, D.; Durante, M.; Löbrich, M.; Taucher-Scholz, G. DNA Double-Strand Breaks in Heterochromatin Elicit Fast Repair Protein Recruitment, Histone H2AX Phosphorylation and Relocation to Euchromatin. Nucleic Acids Res. 2011, 39, 6489–6499. [Google Scholar] [CrossRef]
- García-Nieto, P.E.; Schwartz, E.K.; King, D.A.; Paulsen, J.; Collas, P.; Herrera, R.E.; Morrison, A.J. Carcinogen Susceptibility Is Regulated by Genome Architecture and Predicts Cancer Mutagenesis. EMBO J. 2017, 36, 2829–2843. [Google Scholar] [CrossRef]
- Fortuny, A.; Polo, S.E. The Response to DNA Damage in Heterochromatin Domains. Chromosoma 2018, 127, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulos-Soares, I.; Parada, G.E.; Wong, H.Y.; Miska, E.A.; Kwok, C.K.; Hemberg, M. Alternative Splicing Modulation by G-Quadruplexes. bioRxiv Genom. 2019, 700575. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Colocalization Partners- Treatment | Pearson’s Correlation Coefficient (PCC) | Correlation Rate (%) |
|---|---|---|
| G4s/γH2AX-control | 0.37 ± 0.09 | 34 ± 9 |
| G4s/γH2AX-5Gy | 0.32 ± 0.08 | 23 ± 7 |
| G4s/53BP1-control | 0.36 ± 0.08 | 31 ± 10 |
| G4s/53BP1-5Gy | 0.28 ± 0.10 | 23 ± 11 |
| G4s/MDC1-control | 0.33 ± 0.08 | 27 ± 9 |
| G4s/MDC1-5Gy | 0.32 ± 0.08 | 38 ± 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komůrková, D.; Svobodová Kovaříková, A.; Bártová, E. G-Quadruplex Structures Colocalize with Transcription Factories and Nuclear Speckles Surrounded by Acetylated and Dimethylated Histones H3. Int. J. Mol. Sci. 2021, 22, 1995. https://doi.org/10.3390/ijms22041995
Komůrková D, Svobodová Kovaříková A, Bártová E. G-Quadruplex Structures Colocalize with Transcription Factories and Nuclear Speckles Surrounded by Acetylated and Dimethylated Histones H3. International Journal of Molecular Sciences. 2021; 22(4):1995. https://doi.org/10.3390/ijms22041995
Chicago/Turabian StyleKomůrková, Denisa, Alena Svobodová Kovaříková, and Eva Bártová. 2021. "G-Quadruplex Structures Colocalize with Transcription Factories and Nuclear Speckles Surrounded by Acetylated and Dimethylated Histones H3" International Journal of Molecular Sciences 22, no. 4: 1995. https://doi.org/10.3390/ijms22041995