
Challenges and Costs of Asexuality: Variation in Premeiotic Genome Duplication in Gynogenetic Hybrids from Cobitis taenia Complex


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Studied and Crossing Experiments
2.2. Pachytene Chromosomes and Immunofluorescent Staining
2.3. Diplotene Chromosomes
2.4. Fluorescence In Situ Hybridization
2.5. Whole-Mount Immunofluorescence Staining
2.6. Whole-Mount Fluorescence In Situ Hybridization
2.7. Confocal Laser Scanning Microscopy
2.8. Wide-Field and Fluorescence Microscopy
3. Results
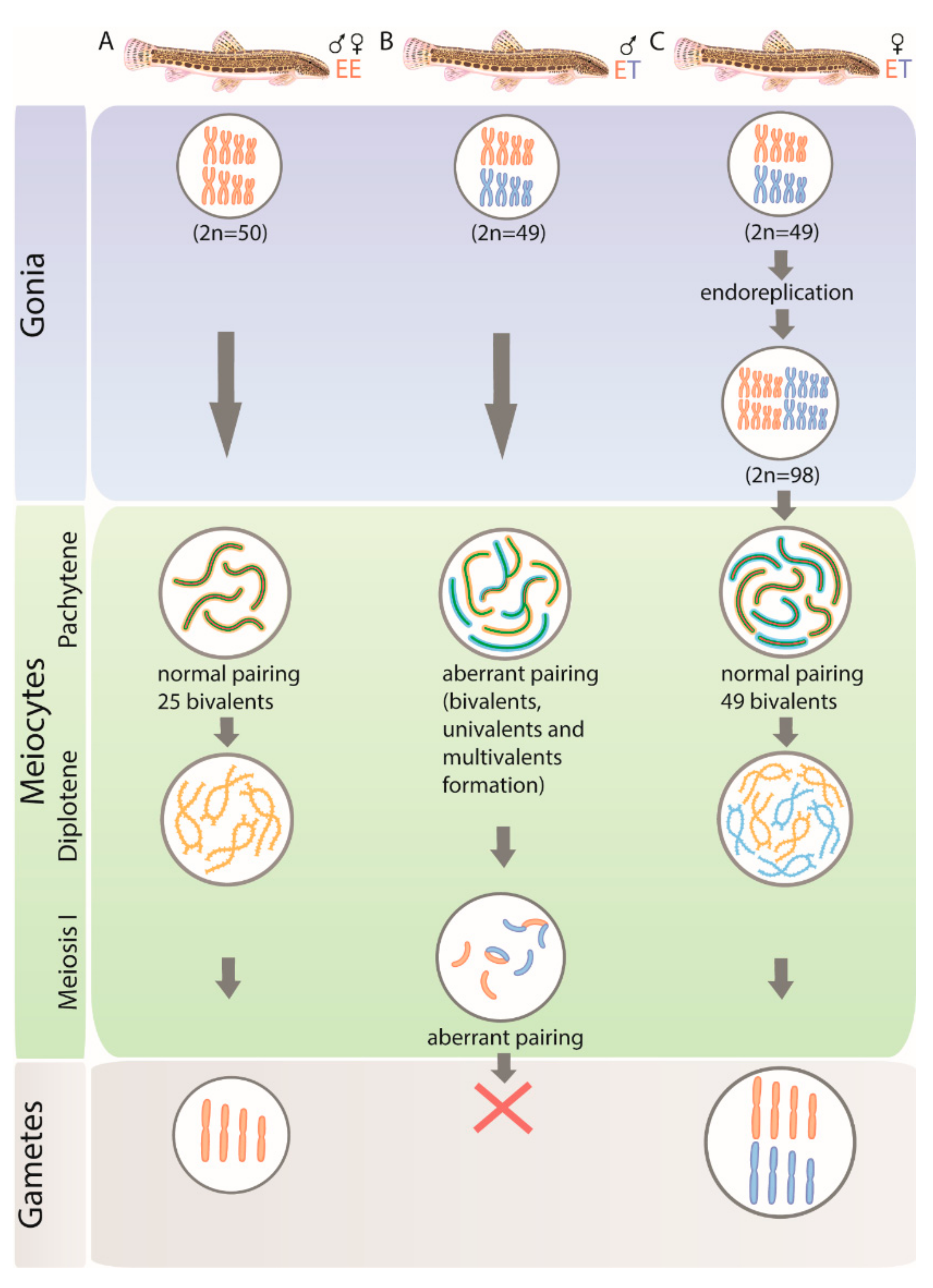
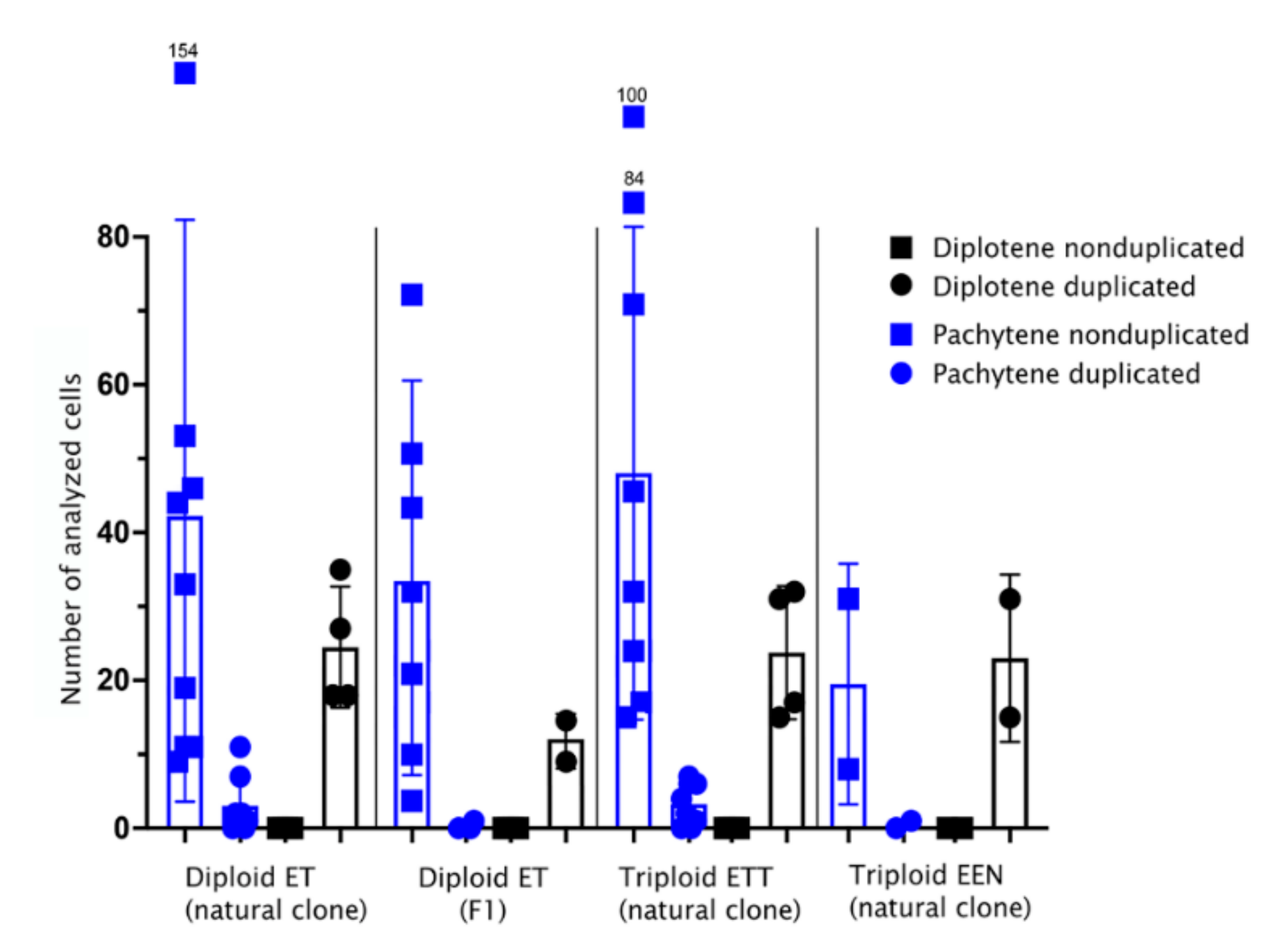
3.1. All Diplotene Oocytes Have a Duplicated Genome and Properly Paired Chromosomes with Bivalents
3.2. Pachytene Falls into Two Types: Those with Duplicated Chromosomes and Proper Bivalents and Those with Few Bivalents and Many Univalents
3.3. Pachytene Oocytes Containing Bivalents and Univalents Do Not Have a Duplicated Genome, While Those with Only Bivalents Do
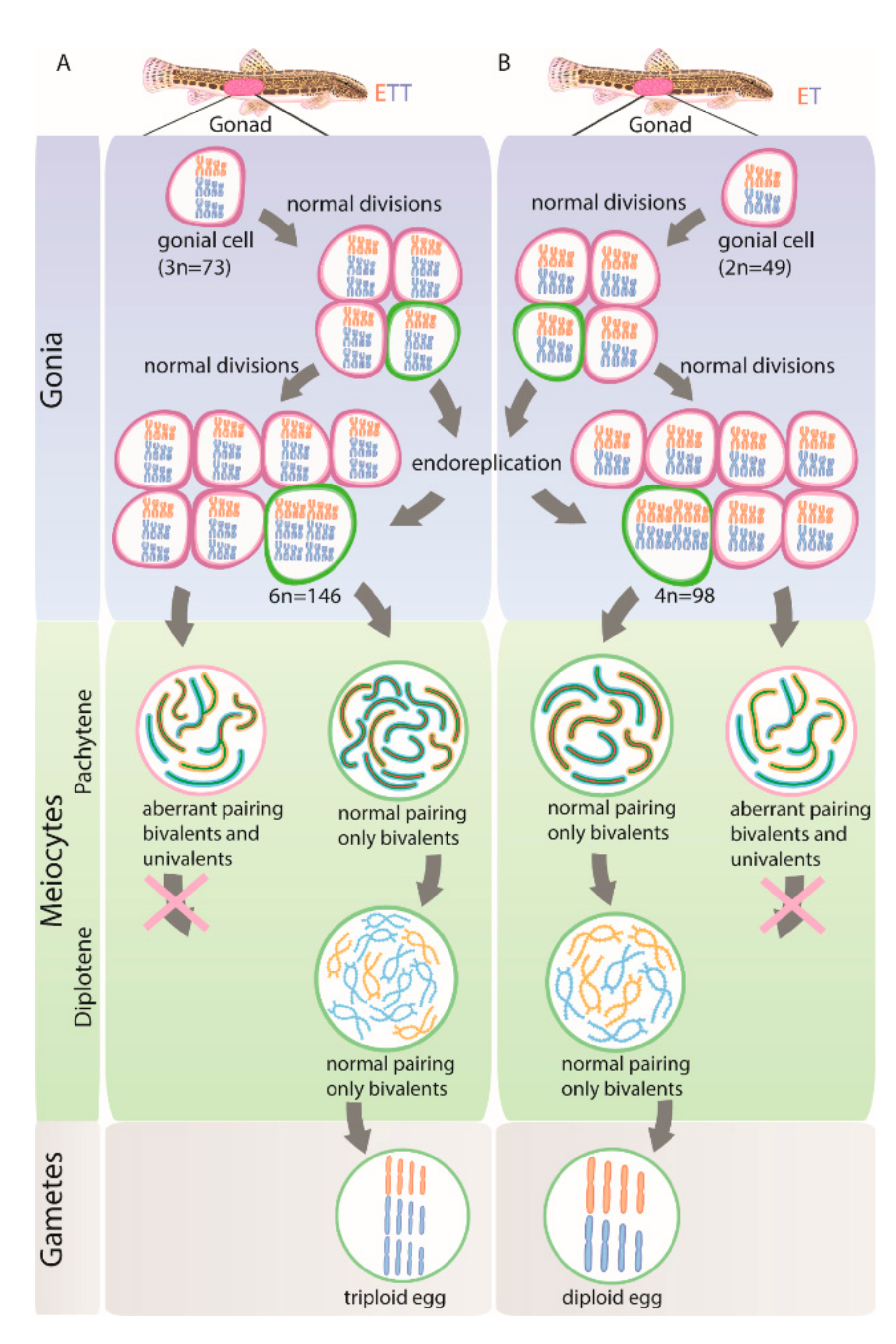
3.4. Gonocytes (Germ Cells) Also Occur in Unduplicated and Duplicated Forms within Intact Gonad of Hybrid Females
4. Discussion
4.1. Genome Duplication Is Restricted to the Minor Cell Population While the Majority of Cells May Not Proceed beyond Pachytene
4.2. Sex-Specific Differences in Meiotic Checkpoints and Stringency of Pairing Chromosomal Control
4.3. Initiation of Premeiotic Genome Duplication
4.4. Implications for Ecology and Evolution of Asexual Organisms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates Sunderland: Sunderland, MA, USA, 2004. [Google Scholar]
- Avise, J.C. Speciation (review). Perspect. Biol. Med. 2005, 48, 315–316. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H.; Willis, J.H. Plant speciation. Science 2007, 317, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.L.; Hodges, S.A. Are natural hybrids fit or unfit relative to their parents? Trends Ecol. Evol. 1995, 10, 67–71. [Google Scholar] [CrossRef]
- Rieseberg, L.H. Chromosomal rearrangements and speciation. Trends Ecol. Evol. 2001, 16, 351–358. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. The evolutionary genetics of speciation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1998, 353, 287–305. [Google Scholar] [CrossRef]
- Maheshwari, S.; Barbash, D.A. The genetics of hybrid incompatibilities. Annu. Rev. Genet. 2011, 45, 331–355. [Google Scholar] [CrossRef]
- Payseur, B.A.; Krenz, J.G.; Nachman, M.W. Differential patterns of introgression across the X chromosome in a hybrid zone between two species of house mice. Evolution 2004, 58, 2064–2078. [Google Scholar] [CrossRef]
- Geraldes, A.; Ferrand, N.; Nachman, M.W. Contrasting patterns of introgression at X-linked loci across the hybrid zone between subspecies of the European rabbit (Oryctolagus cuniculus). Genetics 2006, 173, 919–933. [Google Scholar] [CrossRef] [Green Version]
- Ernst, A. Bastardierung Als Ursache der Apogamie Im Pflanzenreich. Eine Hypothese zur Experimentellen Vererbungs- und Abstammungslehre; Fischer: Jena, Germany, 1918; pp. 1–704. [Google Scholar]
- Bullini, L. Origin and evolution of animal hybrid species. Trends Ecol. Evol. 1994, 9, 422–426. [Google Scholar] [CrossRef]
- Choleva, L.; Janko, K.; Gelas, K.D.; Bohlen, J.; Šlechtová, V.; Rábová, M.; Ráb, P. Synthesis of clonality and polyploidy in vertebrate animals by hybridization between two sexual species. Evolution 2012, 66, 2191–2203. [Google Scholar] [CrossRef]
- Stenberg, P.; Saura, A. Meiosis and its deviations in polyploid animals. Cytogenet. Genome Res. 2013, 140, 185–203. [Google Scholar] [CrossRef]
- Lenormand, T.; Engelstädter, J.; Johnston, S.E.; Wijnker, E.; Haag, C.R. Evolutionary mysteries in meiosis. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2016, 371, 20160001. [Google Scholar] [CrossRef] [Green Version]
- Lampert, K.P. Facultative parthenogenesis in vertebrates: Reproductive error or chance? Sex. Dev. 2008, 2, 290–301. [Google Scholar] [CrossRef]
- Brownfield, L.; Köhler, C. Unreduced gamete formation in plants: Mechanisms and prospects. J. Exp. Bot. 2011, 62, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Mason, A.S.; Pires, J.C. Unreduced gametes: Meiotic mishap or evolutionary mechanism? Trends Genet. 2015, 31, 5–10. [Google Scholar] [CrossRef]
- Stenberg, P.; Saura, A. Cytology of asexual animals. In Lost Sex: The Evolutionary Biology of Parthenogenesis; Schön, I., Martens, K., Dijk, P., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 63–74. [Google Scholar]
- Neaves, W.B.; Baumann, P. Unisexual reproduction among vertebrates. Trends Genet. 2011, 27, 81–88. [Google Scholar] [CrossRef]
- Stöck, M.; Dedukh, D.; Reifová, R.; Lamatsch, D.K.; Starostová, Z.; Janko, K. Sex chromosomes in meiotic, hemiclonal, clonal and polyploid hybrid vertebrates: Along the ‘extended speciation continuum’. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200103. [Google Scholar] [CrossRef]
- Otto, S.P.; Lenormand, T. Resolving the paradox of sex and recombination. Nat. Rev. Genet. 2002, 3, 252–261. [Google Scholar] [CrossRef]
- Avise, J.C. Clonality: The Genetics, Ecology, and Evolution of Sexual Abstinence in Vertebrate Animals; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Janko, K.; Pačes, J.; Wilkinson-Herbots, H.; Costa, R.J.; Roslein, J.; Drozd, P.; Iakovenko, N.; Rídl, J.; Hroudová, M.; Kočí, J.; et al. Hybrid asexuality as a primary postzygotic barrier between nascent species: On the interconnection between asexuality, hybridization and speciation. Mol. Ecol. 2018, 27, 248–263. [Google Scholar] [CrossRef] [Green Version]
- Dedukh, D.; Majtánová, Z.; Marta, A.; Pšenička, M.; Kotusz, J.; Klíma, J.; Juchno, D.; Boron, A.; Janko, K. Parthenogenesis as a solution to hybrid sterility: The mechanistic basis of meiotic distortions in clonal and sterile hybrids. Genetics 2020, 215, 975–987. [Google Scholar] [CrossRef]
- Suomalainen, E. Cytology and Evolution in Parthenogenesis; CRC Press: Boca Raton, FL, USA, 1987. [Google Scholar]
- Storme, N.D.; Geelen, D. Sexual polyploidization in plants—Cytological mechanisms and molecular regulation. New Phytol. 2013, 198, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Macgregor, H.C.; Uzzell, T.M. Gynogenesis in salamanders related to Ambystoma jeffersonianum. Science 1964, 143, 1043–1045. [Google Scholar] [CrossRef]
- Lutes, A.A.; Neaves, W.B.; Baumann, D.P.; Wiegraebe, W.; Baumann, P. Sister chromosome pairing maintains heterozygosity in parthenogenetic lizards. Nature 2010, 464, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Fujimoto, T.; Murakami, M.; Yamaha, E.; Arai, K. Clonal reproduction assured by sister chromosome pairing in dojo loach, a teleost fish. Chromosome Res. 2018, 26, 243–253. [Google Scholar] [CrossRef]
- Dedukh, D.; Riumin, S.; Chmielewska, M.; Rozenblut-Kościsty, B.; Kolenda, K.; Kazmierczak, M.; Dudzik, A.; Ogielska, M.; Krasikova, A. Micronuclei in germ cells of hybrid frogs from Pelophylax esculentus complex contain gradually eliminated chromosomes. Sci. Rep. 2020, 10, 1–13. [Google Scholar]
- Kuroda, M.; Fujimoto, T.; Murakami, M.; Yamaha, E.; Arai, K. Aberrant meiotic configurations cause sterility in clone-origin triploid and inter-group hybrid males of the dojo loach, Misgurnus anguillicaudatus. Cytogenet. Genome Res. 2019, 158, 46–54. [Google Scholar] [CrossRef]
- Bateson, W. Heredity and variation in modern lights. In Darwin and Modern Science; Cambridge University Press: Cambridge, UK, 1909; pp. 85–101. [Google Scholar]
- Moritz, C. Genetic diversity and the dynamics of hybrid parthenogenesis in Cnemidophorus (Teiidae) and Heteronotia (Gekkonidae). In Evolution and Ecology of Unisexual Vertebrates, Bulletin; Dawley, R.M., Bogart, J.P., Eds.; New York State Museum: Albany, NY, USA, 1989; Volume 466, pp. 87–112. [Google Scholar]
- Russell, S.T. Evolution of intrinsic post-zygotic reproductive isolation in fish. Ann. Zool. Fenn. 2003, 40, 321–329. [Google Scholar]
- Hamaguchi, S.; Sakaizumi, M. Sexually differentiated mechanisms of sterility in interspecific hybrids between Oryzias latipes and O. curvinotus. J. Exp. Zool. 1992, 263, 323–329. [Google Scholar] [CrossRef]
- Shimizu, Y.; Shibata, N.; Sakaizumi, M.; Yamashita, M. Production of diploid eggs through premeiotic endomitosis in the hybrid medaka between Oryzias latipes and O. curvinotus. Zool. Sci. 2000, 17, 951–958. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.A.; Schnittker, R.R.; Yu, Z.; Munday, S.S.; Baumann, D.P.; Neaves, W.B.; Baumann, P. Widespread failure to complete meiosis does not impair fecundity in parthenogenetic whiptail lizards. Development 2016, 143, 4486–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohlen, J.; Ráb, P. Species and hybrid richness in spined loaches of the genus Cobitis (Teleostei: Cobitidae), with a checklist of European forms and suggestions for conservation. J. Fish Biol. 2001, 59, 75–89. [Google Scholar] [CrossRef]
- Janko, K.; Flajšhans, M.; Choleva, L.; Bohlen, J.; Šlechtová, V.; Rábová, M.; Lajbner, Z.; Šlechta, V.; Ivanova, P.; Dobrovolov, I.; et al. Diversity of European spined loaches (genus Cobitis l.): An update of the geographic distribution of the Cobitis taenia hybrid complex with a description of new molecular tools for species and hybrid determination. J. Fish Biol. 2007, 71, 387–408. [Google Scholar] [CrossRef]
- Majtánová, Z.; Choleva, L.; Symonová, R.; Ráb, P.; Kotusz, J.; Pekárik, L.; Janko, K. Asexual reproduction does not apparently increase the rate of chromosomal evolution: Karyotype stability in diploid and triploid clonal hybrid fish (Cobitis, Cypriniformes, Teleostei). PLoS ONE 2016, 11, e0146872. [Google Scholar] [CrossRef] [Green Version]
- Marta, A.; Dedukh, D.; Bartoš, O.; Majtánová, Z.; Janko, K. Cytogenetic characterization of seven novel satDNA markers in two species of spined loaches (Cobitis) and their clonal hybrids. Genes 2020, 11, 617. [Google Scholar] [CrossRef]
- Juchno, D.; Boroń, A. Comparative histology of the testes of the spined loach Cobitis taenia l. and natural allotetraploids of Cobitis (Pisces, Cobitidae). Hydrobiologia 2006, 573, 45–53. [Google Scholar] [CrossRef]
- Juchno, D.; Arai, K.; Boroń, A.; Kujawa, R. Meiotic chromosome configurations in oocytes of Cobitis taenia and its polyploid hybrids. Ichthyol. Res. 2017, 64, 240–243. [Google Scholar] [CrossRef] [Green Version]
- Janko, K.; Vasil’ev, V.P.; Ráb, P.; Rábová, M. Genetic and morphological analyses of 50-chromosome spined loaches (Cobitis, Cobitidae, Pisces) from the black sea basin that are morphologically similar to C. taenia, with the description of a new species. Folia Zool. 2005, 54, 405. [Google Scholar]
- Janko, K.; Kotusz, J.; De Gelas, K.; Slechtová, V.; Opoldusová, Z.; Drozd, P.; Choleva, L.; Popiołek, M.; Baláž, M. Dynamic formation of asexual diploid and polyploid lineages: Multilocus analysis of Cobitis reveals the mechanisms maintaining the diversity of clones. PLoS ONE 2012, 7, e45384. [Google Scholar] [CrossRef] [Green Version]
- Araya-Jaime, C.; Serrano, É.A.; de Andrade Silva, D.M.Z.; Yamashita, M.; Iwai, T.; Oliveira, C.; Foresti, F. Surface-spreading technique of meiotic cells and immunodetection of synaptonemal complex proteins in teleostean fishes. Mol. Cytogenet. 2015, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Blokhina, Y.P.; Nguyen, A.D.; Draper, B.W.; Burgess, S.M. The telomere bouquet is a hub where meiotic double-strand breaks, synapsis, and stable homolog juxtaposition are coordinated in the zebrafish, Danio rerio. PLOS Genet. 2019, 15, e1007730. [Google Scholar] [CrossRef] [Green Version]
- Callan, H.G. Lampbrush Chromosomes; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Saito, K.; Sakai, C.; Kawasaki, T.; Sakai, N. Telomere distribution pattern and synapsis initiation during spermatogenesis in zebrafish. Dev. Dyn. 2014, 243, 1448–1456. [Google Scholar] [CrossRef]
- Itono, M.; Morishima, K.; Fujimoto, T.; Bando, E.; Yamaha, E.; Arai, K. Premeiotic endomitosis produces diploid eggs in the natural clone loach, Misgurnus anguillicaudatus (Teleostei: Cobitidae). J. Exp. Zool. Part A Comp. Exp. Biol. 2006, 305, 513–523. [Google Scholar] [CrossRef]
- Dedukh, D.; Litvinchuk, S.; Rosanov, J.; Mazepa, G.; Saifitdinova, A.; Shabanov, D.; Krasikova, A. Optional endoreplication and selective elimination of parental genomes during oogenesis in diploid and triploid hybrid European water frogs. PLoS ONE 2015, 10, e0123304. [Google Scholar] [CrossRef] [Green Version]
- Roeder, G.S.; Bailis, J.M. The pachytene checkpoint. Trends Genet. 2000, 16, 395–403. [Google Scholar] [CrossRef]
- Subramanian, V.V.; Hochwagen, A. The meiotic checkpoint network: Step-by-step through meiotic prophase. Cold Spring Harb. Perspect. Biol. 2014, 6, a016675. [Google Scholar] [CrossRef]
- MacQueen, A.J.; Hochwagen, A. Checkpoint mechanisms: The puppet masters of meiotic prophase. Trends Cell Biol. 2011, 21, 393–400. [Google Scholar] [CrossRef]
- Bohr, T.; Ashley, G.; Eggleston, E.; Firestone, K.; Bhalla, N. Synaptonemal complex components are required for meiotic checkpoint function in Caenorhabditis elegans. Genetics 2016, 204, 987–997. [Google Scholar] [CrossRef] [Green Version]
- Marcet-Ortega, M.; Pacheco, S.; Martínez-Marchal, A.; Castillo, H.; Flores, E.; Jasin, M.; Keeney, S.; Roig, I. P53 and TAp63 participate in the recombination-dependent pachytene arrest in mouse spermatocytes. PLoS Genet. 2017, 13, e1006845. [Google Scholar] [CrossRef]
- Chen, X.; Gaglione, R.; Leong, T.; Bednor, L.; de Los Santos, T.; Luk, E.; Airola, M.; Hollingsworth, N.M. Mek1 coordinates meiotic progression with DNA break repair by directly phosphorylating and inhibiting the yeast pachytene exit regulator Ndt80. PLoS Genet. 2018, 14, e1007832. [Google Scholar] [CrossRef]
- Musacchio, A.; Salmon, E.D. The spindle-assembly checkpoint in space and time. Nat. Rev. Mol. Cell Biol. 2007, 8, 379–393. [Google Scholar] [CrossRef]
- Lane, S.; Kauppi, L. Meiotic spindle assembly checkpoint and aneuploidy in males versus females. Cell Mol. Life Sci. 2019, 76, 1135–1150. [Google Scholar] [CrossRef] [Green Version]
- Eaker, S.; Cobb, J.; Pyle, A.; Handel, M.A. Meiotic prophase abnormalities and metaphase cell death in MLH1-deficient mouse spermatocytes: Insights into regulation of spermatogenic progress. Dev. Biol. 2002, 249, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Burgoyne, P.S.; Mahadevaiah, S.K.; Turner, J.M.A. The consequences of asynapsis for mammalian meiosis. Nat. Rev. Genet. 2009, 10, 207–216. [Google Scholar] [CrossRef]
- Kurahashi, H.; Kogo, H.; Tsutsumi, M.; Inagaki, H.; Ohye, T. Failure of homologous synapsis and sex-specific reproduction problems. Front. Genet. 2012, 3, 112. [Google Scholar] [CrossRef] [Green Version]
- Fielder, S.M.; Kempfer, R.; Kelly, W.G. Multiple sex-specific differences in the regulation of meiotic progression in C. elegans. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Shibata, N.; Yamashita, M. Spermiogenesis without preceding meiosis in the hybrid medaka between Oryzias latipes and O. curvinotus. J. Exp. Zool. 1997, 279, 102–112. [Google Scholar] [CrossRef]
- Zhang, Q.; Arai, K.; Yamashita, M. Cytogenetic mechanisms for triploid and haploid egg formation in the triploid loach Misgurnus anguillicaudatus. J. Exp. Zool. 1998, 281, 608–619. [Google Scholar] [CrossRef]
- Nagaoka, S.I.; Hodges, C.A.; Albertini, D.F.; Hunt, P.A. Oocyte-specific differences in cell-cycle control create an innate susceptibility to meiotic errors. Curr. Biol. 2011, 21, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, S.I.; Hassold, T.J.; Hunt, P.A. Human aneuploidy: Mechanisms and new insights into an age-old problem. Nat. Rev. Genet. 2012, 13, 493–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, C.R.; Sumpter, J.P. Oocyte growth and development in teleosts. Rev. Fish Biol. Fish. 1996, 6, 287–318. [Google Scholar] [CrossRef]
- Nakamura, S.; Kobayashi, K.; Nishimura, T.; Tanaka, M. Ovarian germline stem cells in the teleost fish, medaka (Oryzias latipes). Int. J. Biol. Sci. 2011, 7, 403–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wildner, D.D.; Grier, H.; Quagio-Grassiotto, I. Female germ cell renewal during the annual reproductive cycle in ostariophysians fish. Theriogenology 2013, 79, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, H.; Morishima, K.; Kusuda, S.; Yamaha, E.; Arai, K. Diploid sperm produced by artificially sex-reversed clone loaches. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2007, 307, 75–83. [Google Scholar] [CrossRef]
- Lee, H.O.; Davidson, J.M.; Duronio, R.J. Endoreplication: Polyploidy with purpose. Genes Dev. 2009, 23, 2461–2477. [Google Scholar] [CrossRef] [Green Version]
- Calvi, B.R. Making big cells: One size does not fit all. Proc. Natl. Acad. Sci. USA 2013, 110, 9621–9622. [Google Scholar] [CrossRef] [Green Version]
- Fox, D.T.; Duronio, R.J. Endoreplication and polyploidy: Insights into development and disease. Development 2013, 140, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Orr-Weaver, T.L. When bigger is better: The role of polyploidy in organogenesis. Trends Genet. 2015, 31, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Losick, V.P.; Fox, D.T.; Spradling, A.C. Polyploidization and cell fusion contribute to wound healing in the adult Drosophila epithelium. Curr. Biol. 2013, 23, 2224–2232. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, J.; Jackman, C.P.; Cox, A.H.; Trembley, M.A.; Balowski, J.J.; Cox, B.D.; De Simone, A.; Dickson, A.L.; Di Talia, S.; et al. Tension creates an endoreplication wavefront that leads regeneration of epicardial tissue. Dev. Cell 2017, 42, 600–615. [Google Scholar] [CrossRef] [Green Version]
- González-Rosa, J.M.; Sharpe, M.; Field, D.; Soonpaa, M.H.; Field, L.J.; Burns, C.E.; Burns, C.G. Myocardial polyploidization creates a barrier to heart regeneration in zebrafish. Dev. Cell 2018, 44, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Sauer, K.; Knoblich, J.A.; Richardson, H.; Lehner, C.F. Distinct modes of cyclin E/Cdc2c kinase regulation and S-phase control in mitotic and endoreduplication cycles of Drosophila embryogenesis. Genes Dev. 1995, 9, 1327–1339. [Google Scholar] [CrossRef] [Green Version]
- Diril, M.K.; Ratnacaram, C.K.; Padmakumar, V.C.; Du, T.; Wasser, M.; Coppola, V.; Tessarollo, L.; Kaldis, P. Cyclin-dependent kinase 1 (Cdk1) is essential for cell division and suppression of DNA re-replication but not for liver regeneration. Proc. Natl. Acad. Sci. USA 2012, 109, 3826–3831. [Google Scholar] [CrossRef] [Green Version]
- Nannas, N.J.; Murray, A.W. Complications dawn for kinetochore regulation by Aurora. Proc. Natl. Acad. Sci. USA 2012, 109, 15972–15973. [Google Scholar] [CrossRef] [Green Version]
- Rotelli, M.D.; Policastro, R.A.; Bolling, A.M.; Killion, A.W.; Weinberg, A.J.; Dixon, M.J.; Zentner, G.E.; Walczak, C.E.; Lilly, M.A.; Calvi, B.R. A Cyclin A-Myb-MuvB-Aurora B network regulates the choice between mitotic cycles and polyploid endoreplication cycles. PLoS Genet. 2019, 15, e1008253. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.R.; Maiato, H.; Earnshaw, W.C.; Carmena, M. Essential roles of Drosophila inner centromere protein (INCENP) and Aurora B in Histone H3 phosphorylation, metaphase chromosome alignment, kinetochore disjunction, and chromosome segregation. J. Cell Biol. 2001, 153, 865–880. [Google Scholar] [CrossRef]
- Giet, R.; Glover, D.M. Drosophila Aurora B kinase is required for histone H3 phosphorylation and condensin recruitment during chromosome condensation and to organize the central spindle during cytokinesis. J. Cell Biol. 2001, 152, 669–682. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Stout, J.R.; Dharmaiah, S.; Yde, S.; Calvi, B.R.; Walczak, C.E. Transient endoreplication down-regulates the kinesin-14 HSET and contributes to genomic instability. Mol. Biol. Cell 2016, 27, 2911–2923. [Google Scholar] [CrossRef]
- Schultz, R.J. Hybridization, unisexuality, and polyploidy in the teleost Poeciliopsis (Poeciliidae) and other vertebrates. Am. Nat. 1969, 103, 605–619. [Google Scholar] [CrossRef]
- Cuellar, O. On the origin of parthenogenesis in vertebrates: The cytogenetic factors. Am. Nat. 1974, 108, 625–648. [Google Scholar] [CrossRef]
- Sinclair, E.A.; Pramuk, J.B.; Bezy, R.L.; Crandall, K.A.; Sites, J.W., Jr. DNA evidence for nonhybrid origins of parthenogenesis in natural populations of vertebrates. Evol. Int. J. Org. Evol. 2010, 64, 1346–1357. [Google Scholar] [CrossRef]
- Maciak, S.; Janko, K.; Kotusz, J.; Choleva, L.; Boroń, A.; Juchno, D.; Kujawa, R.; Kozłowski, J.; Konarzewski, M. Standard metabolic rate (SMR) is inversely related to erythrocyte and genome size in allopolyploid fish of the Cobitis taenia hybrid complex. Funct. Ecol. 2011, 25, 1072–1078. [Google Scholar] [CrossRef]
- Juchno, D.; Boroń, A.; Kujawa, R.; Szlachciak, J.; Szacherski, S.; Spóz, A.; Grabowska, A. Comparison of egg and offspring size of karyologically identified spined loach, Cobitis taenia L., and hybrid triploid Cobitis females (Pisces, Cobitidae). Fish. Aquat. Life 2013, 21, 293–299. [Google Scholar]
- Juchno, D.; Jabłońska, O.; Boroń, A.; Kujawa, R.; Leska, A.; Grabowska, A.; Nynca, A.; Świgońska, S.; Król, M.; Spóz, A.; et al. Ploidy-dependent survival of progeny arising from crosses between natural allotriploid Cobitis females and diploid C. taenia males (Pisces, Cobitidae). Genetica 2014, 142, 351–359. [Google Scholar] [CrossRef]
- Sogard, S.M. Size-selective mortality in the juvenile stage of teleost fishes: A review. Bull. Mar. Sci. 1997, 60, 1129–1157. [Google Scholar]
- Schlupp, I.; Plath, M. Male mate choice and sperm allocation in a sexual/asexual mating complex of Poecilia (Poeciliidae, Teleostei). Biol. Lett. 2005, 1, 169–171. [Google Scholar] [CrossRef] [Green Version]
- Mee, J.A.; Otto, S.P. Variation in the strength of male mate choice allows long-term coexistence of sperm-dependent asexuals and their sexual hosts. Evolution 2010, 64, 2808–2819. [Google Scholar] [CrossRef] [PubMed]
- Morgado-Santos, M.; Pereira, H.M.; Vicente, L.; Collares-Pereira, M.J. Mate choice drives evolutionary stability in a hybrid complex. PLoS ONE 2015, 10, e0132760. [Google Scholar] [CrossRef] [Green Version]
- Bobyrev, A.; Burmensky, V.; Vasil’ev, V.; Kriksunov, E.; Lebedeva, E. Coexistence of triploid and diploid forms of spined loach, Cobitis taenia: A model-based approach. Folia Biol. 2003, 51, 55–60. [Google Scholar]
- Leung, C.; Angers, B. Imitating the cost of males: A hypothesis for coexistence of all-female sperm-dependent species and their sexual host. Ecol. Evol. 2018, 8, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Juchno, D.; Boroń, A. Fecundity of the spined loach, Cobitis taenia (Pisces, Cobitidae) and natural allopolyploids of Cobitis from a diploid-polyploid population. Folia Zool. 2010, 59, 35–43. [Google Scholar] [CrossRef]
- Kočí, J.; Röslein, J.; Pačes, J.; Kotusz, J.; Halačka, K.; Koščo, J.; Fedorčák, J.; Iakovenko, N.; Janko, K. No evidence for accumulation of deleterious mutations and fitness degradation in clonal fish hybrids: Abandoning sex without regrets. Mol. Ecol. 2020, 29, 3038–3055. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dedukh, D.; Marta, A.; Janko, K. Challenges and Costs of Asexuality: Variation in Premeiotic Genome Duplication in Gynogenetic Hybrids from Cobitis taenia Complex. Int. J. Mol. Sci. 2021, 22, 12117. https://doi.org/10.3390/ijms222212117
Dedukh D, Marta A, Janko K. Challenges and Costs of Asexuality: Variation in Premeiotic Genome Duplication in Gynogenetic Hybrids from Cobitis taenia Complex. International Journal of Molecular Sciences. 2021; 22(22):12117. https://doi.org/10.3390/ijms222212117
Chicago/Turabian StyleDedukh, Dmitrij, Anatolie Marta, and Karel Janko. 2021. "Challenges and Costs of Asexuality: Variation in Premeiotic Genome Duplication in Gynogenetic Hybrids from Cobitis taenia Complex" International Journal of Molecular Sciences 22, no. 22: 12117. https://doi.org/10.3390/ijms222212117