
Ixodes ricinus Salivary Serpin Iripin-8 Inhibits the Intrinsic Pathway of Coagulation and Complement

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
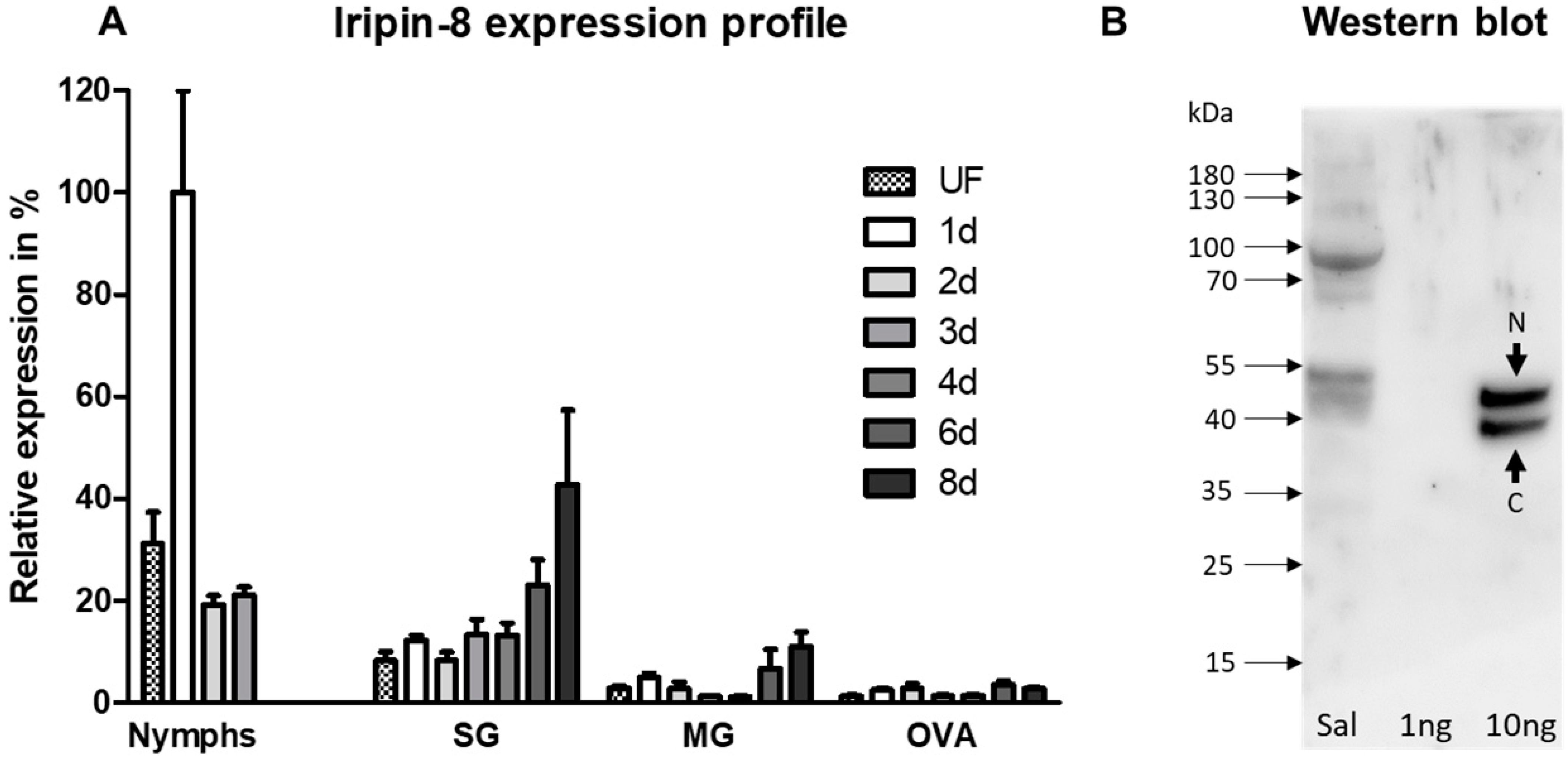
2.1. Iripin-8 Is Predominantly a Salivary Protein with Increased Expression during Tick Feeding
2.2. Sequence Analysis and Production of Recombinant Iripin-8
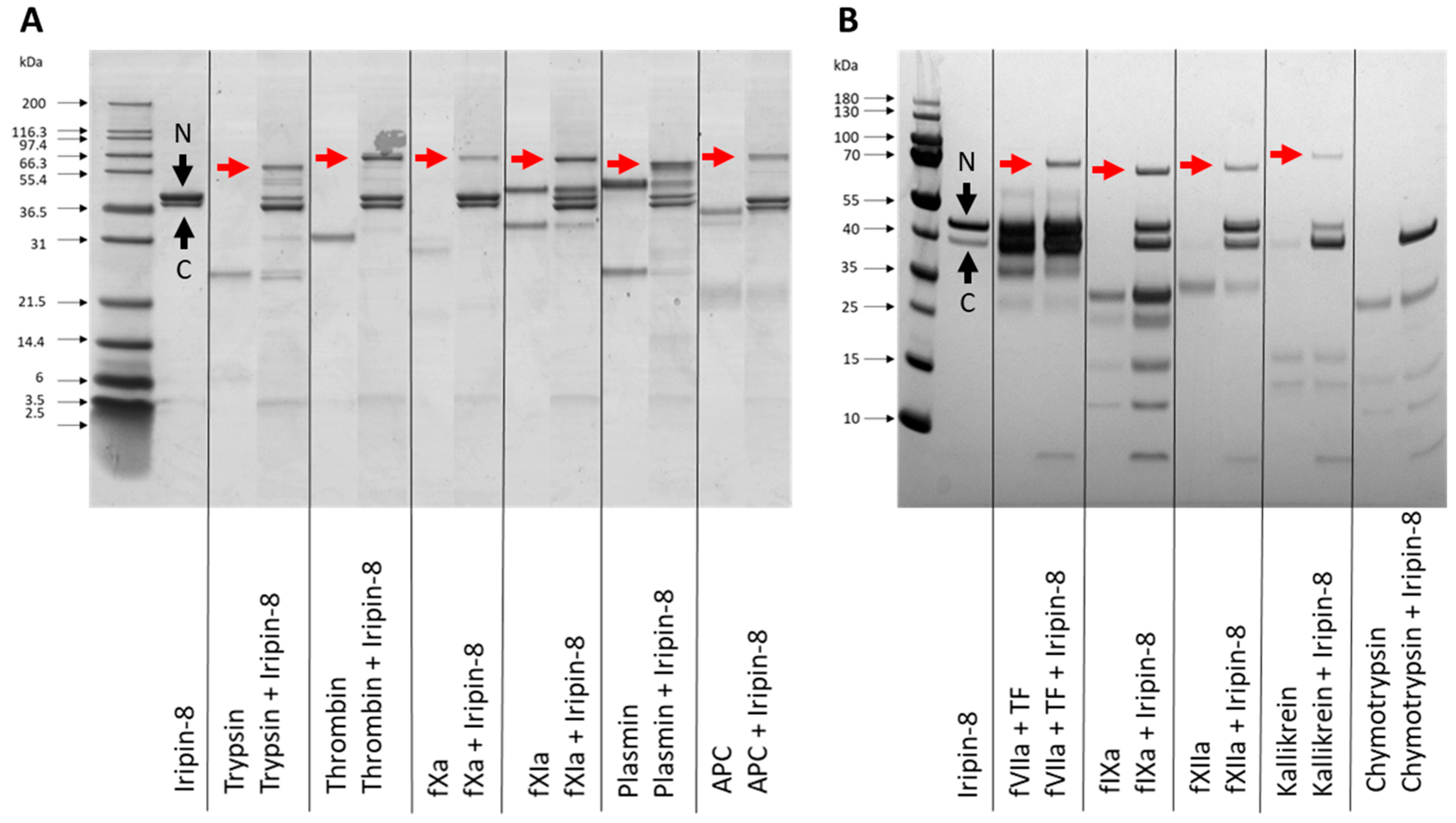
2.3. Iripin-8 Inhibits Serine Proteases Involved in Coagulation
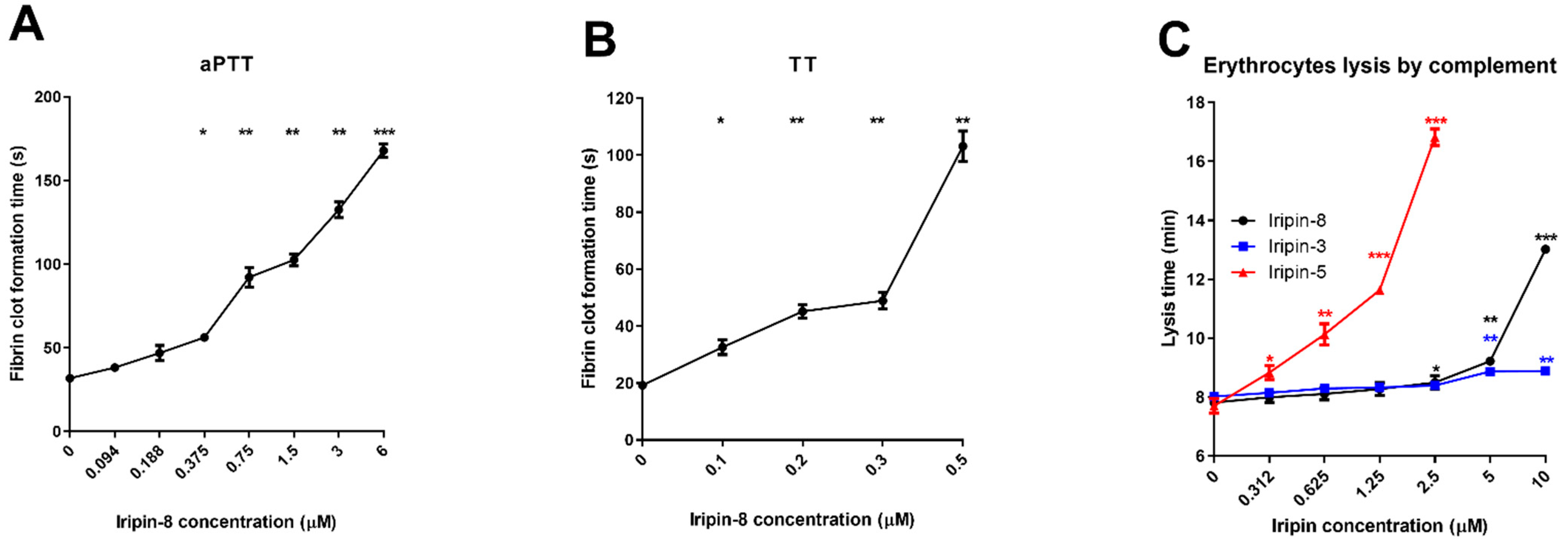
2.4. Iripin-8 Inhibits the Intrinsic and Common Pathways of Blood Coagulation
2.5. Anticomplement Activity of Iripin-8
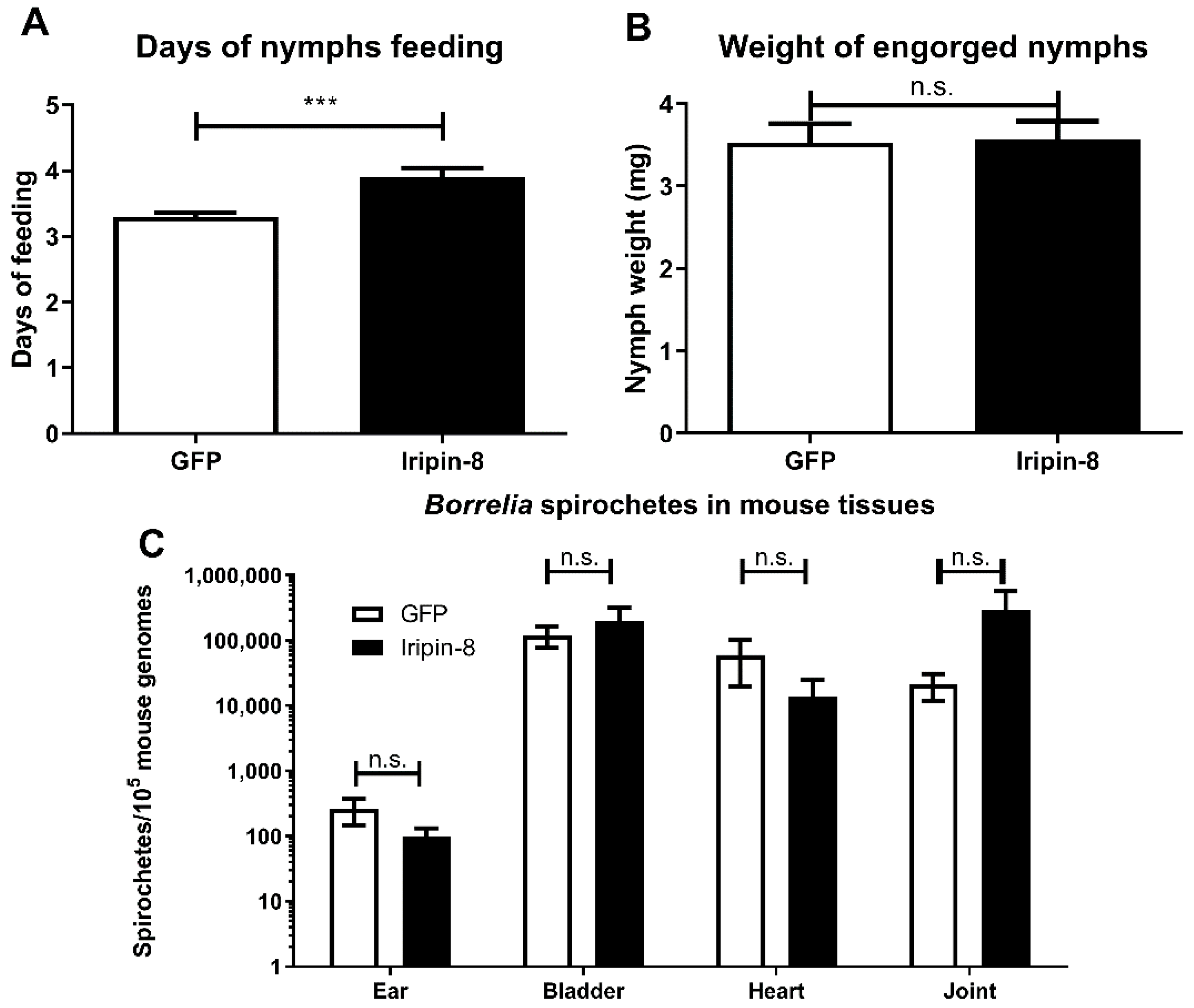
2.6. Iripin-8 Knockdown Influences Tick Feeding but Not Borrelia Transmission
2.7. Role of Iripin-8 in Modulating Host Immunity
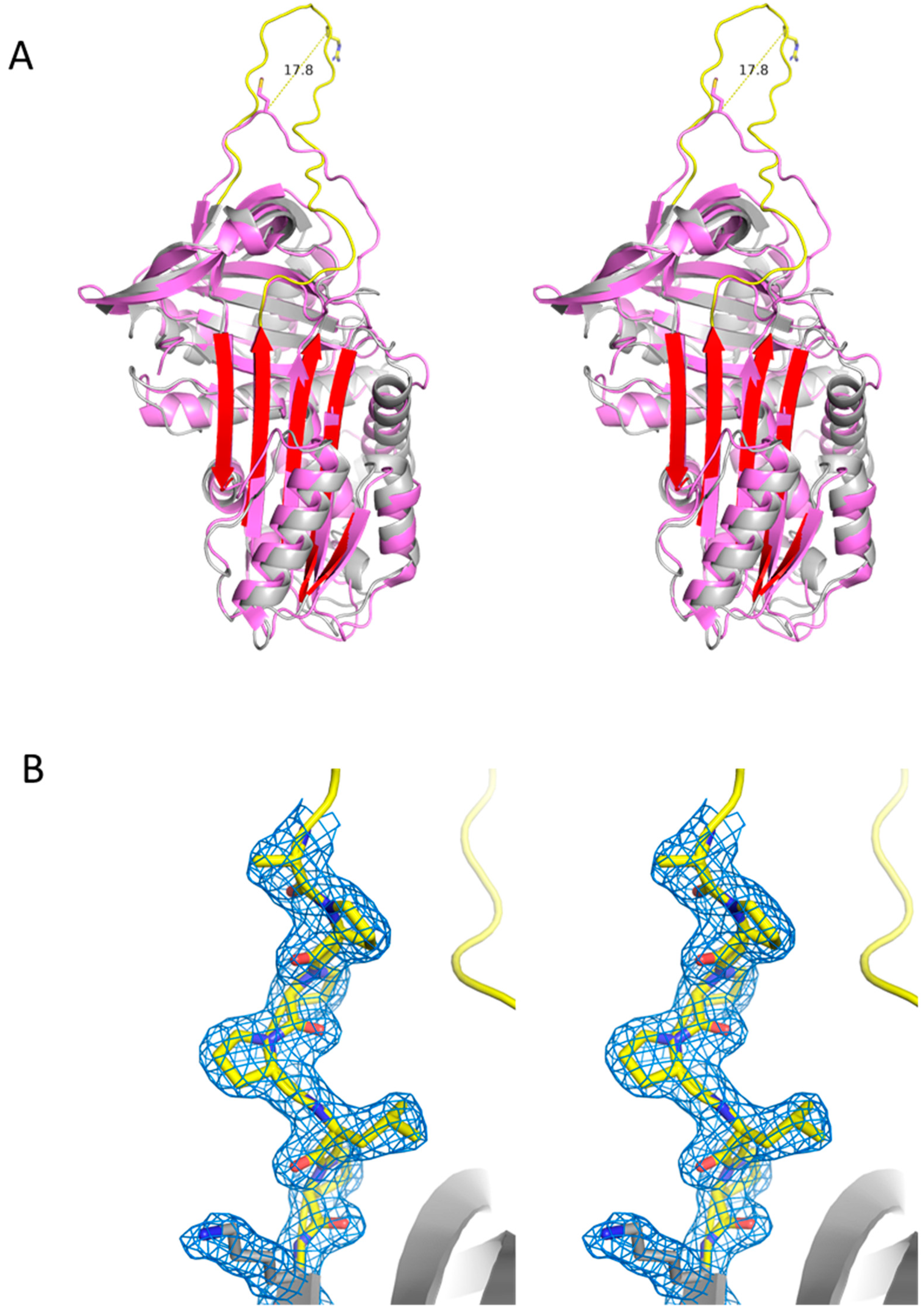
2.8. Structural Features of Iripin-8
3. Discussion
4. Materials and Methods
4.1. Ticks and Laboratory Animals
4.2. Gene Expression Profiling
4.3. RNA Silencing and Borrelia Transmission
4.4. Cloning, Expression, and Purification of Iripin-8
4.5. SDS-PAGE of Complex Formation
4.6. Determination of Inhibition Constants
4.7. Anti-Iripin-8 Serum Production and Western Blotting
4.8. Coagulation Assays
4.9. Crystal Structure Determination
4.10. Complement Assay
4.11. Immunological Assays
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindgren, E.; Talleklint, L.; Polfeldt, T. Impact of climatic change on the northern latitude limit and population density of the disease-transmitting European tick Ixodes ricinus. Environ. Health Perspect. 2000, 108, 119–123. [Google Scholar] [CrossRef]
- Sonenshine, D.E. Biology of Ticks, 2nd ed.; Oxford University Press: Oxford, UK, 2014. [Google Scholar]
- Chmelar, J.; Calvo, E.; Pedra, J.H.; Francischetti, I.M.; Kotsyfakis, M. Tick salivary secretion as a source of antihemostatics. J Proteom. 2012, 75, 3842–3854. [Google Scholar] [CrossRef] [Green Version]
- Kotal, J.; Langhansova, H.; Lieskovska, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecky, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelar, J. Modulation of host immunity by tick saliva. J. Proteom. 2015, 128, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Kazimirova, M.; Stibraniova, I. Tick salivary compounds: Their role in modulation of host defences and pathogen transmission. Front. Cell Infect. Microbiol. 2013, 3, 43. [Google Scholar] [CrossRef] [Green Version]
- Simo, L.; Kazimirova, M.; Richardson, J.; Bonnet, S.I. The Essential Role of Tick Salivary Glands and Saliva in Tick Feeding and Pathogen Transmission. Front. Cell Infect. Microbiol. 2017, 7, 281. [Google Scholar] [CrossRef]
- Stibraniova, I.; Bartikova, P.; Holikova, V.; Kazimirova, M. Deciphering Biological Processes at the Tick-Host Interface Opens New Strategies for Treatment of Human Diseases. Front. Physiol. 2019, 10, 830. [Google Scholar] [CrossRef] [Green Version]
- Krober, T.; Guerin, P.M. An in vitro feeding assay to test acaricides for control of hard ticks. Pest Manag. Sci. 2007, 63, 17–22. [Google Scholar] [CrossRef]
- Bajda, S.; Dermauw, W.; Panteleri, R.; Sugimoto, N.; Douris, V.; Tirry, L.; Osakabe, M.; Vontas, J.; Van Leeuwen, T. A mutation in the PSST homologue of complex I (NADH:ubiquinone oxidoreductase) from Tetranychus urticae is associated with resistance to METI acaricides. Insect Biochem. Mol. Biol. 2017, 80, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Amara, U.; Rittirsch, D.; Flierl, M.; Bruckner, U.; Klos, A.; Gebhard, F.; Lambris, J.D.; Huber-Lang, M. Interaction between the coagulation and complement system. Adv. Exp. Med. Biol. 2008, 632, 71–79. [Google Scholar] [PubMed] [Green Version]
- Manning, J.E.; Cantaert, T. Time to Micromanage the Pathogen-Host-Vector Interface: Considerations for Vaccine Development. Vaccines 2019, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pingen, M.; Schmid, M.A.; Harris, E.; McKimmie, C.S. Mosquito Biting Modulates Skin Response to Virus Infection. Trends Parasitol. 2017, 33, 645–657. [Google Scholar] [CrossRef]
- Chmelar, J.; Kotal, J.; Langhansova, H.; Kotsyfakis, M. Protease Inhibitors in Tick Saliva: The Role of Serpins and Cystatins in Tick-host-Pathogen Interaction. Front. Cell Infect. Microbiol. 2017, 7, 216. [Google Scholar] [CrossRef]
- Wikel, S. Ticks and tick-borne pathogens at the cutaneous interface: Host defenses, tick countermeasures, and a suitable environment for pathogen establishment. Front. Microbiol. 2013, 4, 337. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, H.; Skelly, P.J.; Zipfel, P.F.; Losson, B.; Vanderplasschen, A. Subversion of complement by hematophagous parasites. Dev. Comp. Immunol. 2009, 33, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Nunn, M.A.; Sharma, A.; Paesen, G.C.; Adamson, S.; Lissina, O.; Willis, A.C.; Nuttall, P.A. Complement inhibitor of C5 activation from the soft tick Ornithodoros moubata. J. Immunol. 2005, 174, 2084–2091. [Google Scholar] [CrossRef] [Green Version]
- Huntington, J.A. Serpin structure, function and dysfunction. J. Thromb. Haemost. JTH 2011, 9 (Suppl. 1), 26–34. [Google Scholar] [CrossRef]
- Law, R.H.; Zhang, Q.; McGowan, S.; Buckle, A.M.; Silverman, G.A.; Wong, W.; Rosado, C.J.; Langendorf, C.G.; Pike, R.N.; Bird, P.I.; et al. An overview of the serpin superfamily. Genome Biol. 2006, 7, 216. [Google Scholar] [CrossRef] [Green Version]
- Silverman, G.A.; Whisstock, J.C.; Bottomley, S.P.; Huntington, J.A.; Kaiserman, D.; Luke, C.J.; Pak, S.C.; Reichhart, J.M.; Bird, P.I. Serpins flex their muscle: I. Putting the clamps on proteolysis in diverse biological systems. J. Biol. Chem. 2010, 285, 24299–24305. [Google Scholar] [CrossRef] [Green Version]
- Whisstock, J.C.; Silverman, G.A.; Bird, P.I.; Bottomley, S.P.; Kaiserman, D.; Luke, C.J.; Pak, S.C.; Reichhart, J.M.; Huntington, J.A. Serpins flex their muscle: II. Structural insights into target peptidase recognition, polymerization, and transport functions. J. Biol. Chem. 2010, 285, 24307–24312. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Whisstock, J.C.; Bottomley, S.P. Molecular gymnastics: Serpin structure, folding and misfolding. Curr. Opin. Struct. Biol. 2006, 16, 761–768. [Google Scholar] [CrossRef]
- Meekins, D.A.; Kanost, M.R.; Michel, K. Serpins in arthropod biology. Semin. Cell Dev. Biol. 2017, 62, 105–119. [Google Scholar] [CrossRef] [Green Version]
- Porter, L.; Radulovic, Z.; Kim, T.; Braz, G.R.; Da Silva Vaz, I., Jr.; Mulenga, A. Bioinformatic analyses of male and female Amblyomma americanum tick expressed serine protease inhibitors (serpins). Ticks Tick-Borne Dis. 2015, 6, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Ribeiro, J.M. An Insight into the Sialome of the Lone Star Tick, Amblyomma americanum, with a Glimpse on Its Time Dependent Gene Expression. PLoS ONE 2015, 10, e0131292. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F., 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirloni, L.; Kim, T.K.; Pinto, A.F.M.; Yates, J.R., 3rd; da Silva Vaz, I., Jr.; Mulenga, A. Tick-Host Range Adaptation: Changes in Protein Profiles in Unfed Adult Ixodes scapularis and Amblyomma americanum Saliva Stimulated to Feed on Different Hosts. Front. Cell Infect. Microbiol. 2017, 7, 517. [Google Scholar] [CrossRef] [Green Version]
- Leboulle, G.; Crippa, M.; Decrem, Y.; Mejri, N.; Brossard, M.; Bollen, A.; Godfroid, E. Characterization of a novel salivary immunosuppressive protein from Ixodes ricinus ticks. J. Biol. Chem. 2002, 277, 10083–10089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevot, P.P.; Adam, B.; Boudjeltia, K.Z.; Brossard, M.; Lins, L.; Cauchie, P.; Brasseur, R.; Vanhaeverbeek, M.; Vanhamme, L.; Godfroid, E. Anti-hemostatic effects of a serpin from the saliva of the tick Ixodes ricinus. J. Biol. Chem. 2006, 281, 26361–26369. [Google Scholar] [CrossRef] [Green Version]
- Chmelar, J.; Oliveira, C.J.; Rezacova, P.; Francischetti, I.M.; Kovarova, Z.; Pejler, G.; Kopacek, P.; Ribeiro, J.M.; Mares, M.; Kopecky, J.; et al. A tick salivary protein targets cathepsin G and chymase and inhibits host inflammation and platelet aggregation. Blood 2011, 117, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Palenikova, J.; Lieskovska, J.; Langhansova, H.; Kotsyfakis, M.; Chmelar, J.; Kopecky, J. Ixodes ricinus salivary serpin IRS-2 affects Th17 differentiation via inhibition of the interleukin-6/STAT-3 signaling pathway. Infect. Immun. 2015, 83, 1949–1956. [Google Scholar] [CrossRef] [Green Version]
- Chlastakova, A.; Kotal, J.; Berankova, Z.; Kascakova, B.; Martins, L.A.; Langhansova, H.; Prudnikova, T.; Ederova, M.; Kuta Smatanova, I.; Kotsyfakis, M.; et al. Iripin-3, a New Salivary Protein Isolated From Ixodes ricinus Ticks, Displays Immunomodulatory and Anti-Hemostatic Properties In Vitro. Front. Immunol. 2021, 12, 626200. [Google Scholar] [CrossRef]
- Kim, T.K.; Tirloni, L.; Radulovic, Z.; Lewis, L.; Bakshi, M.; Hill, C.; da Silva Vaz, I., Jr.; Logullo, C.; Termignoni, C.; Mulenga, A. Conserved Amblyomma americanum tick Serpin19, an inhibitor of blood clotting factors Xa and XIa, trypsin and plasmin, has anti-haemostatic functions. Int. J. Parasitol. 2015, 45, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Yan, Y.; Zhang, H.; Cao, J.; Zhou, Y.; Xu, Q.; Zhou, J. A serpin from the tick Rhipicephalus haemaphysaloides: Involvement in vitellogenesis. Vet. Parasitol. 2020, 279, 109064. [Google Scholar] [CrossRef]
- Xu, T.; Lew-Tabor, A.; Rodriguez-Valle, M. Effective inhibition of thrombin by Rhipicephalus microplus serpin-15 (RmS-15) obtained in the yeast Pichia pastoris. Ticks Tick-Borne Dis. 2016, 7, 180–187. [Google Scholar] [CrossRef]
- Kovarova, Z.; Chmelar, J.; Sanda, M.; Brynda, J.; Mares, M.; Rezacova, P. Crystallization and diffraction analysis of the serpin IRS-2 from the hard tick Ixodes ricinus. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2010, 66 Pt 11, 1453–1457. [Google Scholar] [CrossRef] [Green Version]
- Ellisdon, A.M.; Zhang, Q.; Henstridge, M.A.; Johnson, T.K.; Warr, C.G.; Law, R.H.; Whisstock, J.C. High resolution structure of cleaved Serpin 42 Da from Drosophila melanogaster. BMC Struct. Biol. 2014, 14, 14. [Google Scholar] [CrossRef] [Green Version]
- Schreuder, H.A.; de Boer, B.; Dijkema, R.; Mulders, J.; Theunissen, H.J.; Grootenhuis, P.D.; Hol, W.G. The intact and cleaved human antithrombin III complex as a model for serpin-proteinase interactions. Nat. Struct. Biol. 1994, 1, 48–54. [Google Scholar] [CrossRef]
- Akazawa, T.; Ogawa, M.; Hayakawa, S.; Hirata, M.; Niwa, T. Structural change of ovalbumin-related protein X by alkali treatment. Poult. Sci. 2018, 97, 1730–1737. [Google Scholar] [CrossRef]
- Yang, L.; Irving, J.A.; Dai, W.; Aguilar, M.I.; Bottomley, S.P. Probing the folding pathway of a consensus serpin using single tryptophan mutants. Sci. Rep. 2018, 8, 2121. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.H.; Souza, M.E.; Vargas, A.P.; Martins, M.S.; Penido, C.M.; Diotaiuti, L. Anticoagulant activity of Triatoma infestans and Panstrongylus megistus saliva (Hemiptera/Triatominae). Acta Trop. 1996, 61, 255–261. [Google Scholar] [CrossRef]
- Lestinova, T.; Rohousova, I.; Sima, M.; de Oliveira, C.I.; Volf, P. Insights into the sand fly saliva: Blood-feeding and immune interactions between sand flies, hosts, and Leishmania. PLoS Negl. Trop. Dis. 2017, 11, e0005600. [Google Scholar] [CrossRef]
- Hoffman, M. Remodeling the blood coagulation cascade. J. Thromb. Thrombolysis 2003, 16, 17–20. [Google Scholar] [CrossRef]
- Kascakova, B.; Kotal, J.; Martins, L.A.; Berankova, Z.; Langhansova, H.; Calvo, E.; Crossley, J.A.; Havlickova, P.; Dycka, F.; Prudnikova, T.; et al. Structural and biochemical characterization of the novel serpin Iripin-5 from Ixodes ricinus. Acta Crystallogr. Sect. D 2021, 77. [Google Scholar] [CrossRef]
- Kotal, J.; Stergiou, N.; Busa, M.; Chlastakova, A.; Berankova, Z.; Rezacova, P.; Langhansova, H.; Schwarz, A.; Calvo, E.; Kopecky, J.; et al. The structure and function of Iristatin, a novel immunosuppressive tick salivary cystatin. Cell. Mol. Life Sci. CMLS 2019, 76, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Ricklin, D.; Hajishengallis, G.; Yang, K.; Lambris, J.D. Complement: A key system for immune surveillance and homeostasis. Nat. Immunol. 2010, 11, 785–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, F.; de Carvalho, A.M.; de Oliveira, C.I. Sand-fly saliva-leishmania-man: The trigger trio. Front. Immunol. 2013, 4, 375. [Google Scholar] [CrossRef] [Green Version]
- Awuoche, E.O. Tsetse fly saliva: Could it be useful in fly infection when feeding in chronically aparasitemic mammalian hosts. Open Vet. J. 2012, 2, 95–105. [Google Scholar]
- Caljon, G.; Van Den Abbeele, J.; Sternberg, J.M.; Coosemans, M.; De Baetselier, P.; Magez, S. Tsetse fly saliva biases the immune response to Th2 and induces anti-vector antibodies that are a useful tool for exposure assessment. Int. J. Parasitol. 2006, 36, 1025–1035. [Google Scholar] [CrossRef]
- Kashuba, E.; Bailey, J.; Allsup, D.; Cawkwell, L. The kinin-kallikrein system: Physiological roles, pathophysiology and its relationship to cancer biomarkers. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2013, 18, 279–296. [Google Scholar] [CrossRef]
- Caljon, G.; De Ridder, K.; De Baetselier, P.; Coosemans, M.; Van Den Abbeele, J. Identification of a tsetse fly salivary protein with dual inhibitory action on human platelet aggregation. PLoS ONE 2010, 5, e9671. [Google Scholar] [CrossRef] [Green Version]
- Mans, B.J. Chemical Equilibrium at the Tick-Host Feeding Interface:A Critical Examination of Biological Relevance in Hematophagous Behavior. Front. Physiol. 2019, 10, 530. [Google Scholar] [CrossRef] [Green Version]
- Calvo, E.; Dao, A.; Pham, V.M.; Ribeiro, J.M. An insight into the sialome of Anopheles funestus reveals an emerging pattern in anopheline salivary protein families. Insect Biochem. Mol. Biol. 2007, 37, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Draxler, D.F.; Sashindranath, M.; Medcalf, R.L. Plasmin: A Modulator of Immune Function. Semin. Thromb. Hemost. 2017, 43, 143–153. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Lawrie, C.H.; Sim, R.B.; Nuttall, P.A. Investigation of the mechanisms of anti-complement activity in Ixodes ricinus ticks. Mol. Immunol. 2005, 42, 31–38. [Google Scholar] [CrossRef]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Valenzuela, J.G.; Charlab, R.; Mather, T.N.; Ribeiro, J.M. Purification, cloning, and expression of a novel salivary anticomplement protein from the tick, Ixodes scapularis. J. Biol. Chem. 2000, 275, 18717–18723. [Google Scholar] [CrossRef] [Green Version]
- Tyson, K.; Elkins, C.; Patterson, H.; Fikrig, E.; de Silva, A. Biochemical and functional characterization of Salp20, an Ixodes scapularis tick salivary protein that inhibits the complement pathway. Insect Mol. Biol. 2007, 16, 469–479. [Google Scholar] [CrossRef]
- Tyson, K.R.; Elkins, C.; de Silva, A.M. A novel mechanism of complement inhibition unmasked by a tick salivary protein that binds to properdin. J. Immunol. 2008, 180, 3964–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daix, V.; Schroeder, H.; Praet, N.; Georgin, J.P.; Chiappino, I.; Gillet, L.; de Fays, K.; Decrem, Y.; Leboulle, G.; Godfroid, E.; et al. Ixodes ticks belonging to the Ixodes ricinus complex encode a family of anticomplement proteins. Insect Mol. Biol. 2007, 16, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Schuijt, T.J.; Coumou, J.; Narasimhan, S.; Dai, J.; Deponte, K.; Wouters, D.; Brouwer, M.; Oei, A.; Roelofs, J.J.; van Dam, A.P.; et al. A tick mannose-binding lectin inhibitor interferes with the vertebrate complement cascade to enhance transmission of the lyme disease agent. Cell Host Microbe 2011, 10, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Chmelar, J.; Kotal, J.; Kopecky, J.; Pedra, J.H.; Kotsyfakis, M. All For One and One For All on the Tick-Host Battlefield. Trends Parasitol. 2016, 32, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Sojka, D.; Franta, Z.; Horn, M.; Caffrey, C.R.; Mares, M.; Kopacek, P. New insights into the machinery of blood digestion by ticks. Trends Parasitol. 2013, 29, 276–285. [Google Scholar] [CrossRef]
- Franta, Z.; Frantova, H.; Konvickova, J.; Horn, M.; Sojka, D.; Mares, M.; Kopacek, P. Dynamics of digestive proteolytic system during blood feeding of the hard tick Ixodes ricinus. Parasites Vectors 2010, 3, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, F.A.; Lins, U.; Bechara, G.H.; Oliveira, P.L. Tracing heme in a living cell: Hemoglobin degradation and heme traffic in digest cells of the cattle tick Boophilus microplus. J. Exp. Biol. 2005, 208 Pt 16, 3093–3101. [Google Scholar] [CrossRef] [Green Version]
- Blisnick, A.A.; Foulon, T.; Bonnet, S.I. Serine Protease Inhibitors in Ticks: An Overview of Their Role in Tick Biology and Tick-Borne Pathogen Transmission. Front. Cell Infect. Microbiol. 2017, 7, 199. [Google Scholar] [CrossRef] [Green Version]
- Mulenga, A.; Tsuda, A.; Onuma, M.; Sugimoto, C. Four serine proteinase inhibitors (serpin) from the brown ear tick, Rhiphicephalus appendiculatus; cDNA cloning and preliminary characterization. Insect Biochem. Mol. Biol. 2003, 33, 267–276. [Google Scholar] [CrossRef]
- Imamura, S.; da Silva Vaz Junior, I.; Sugino, M.; Ohashi, K.; Onuma, M. A serine protease inhibitor (serpin) from Haemaphysalis longicornis as an anti-tick vaccine. Vaccine 2005, 23, 1301–1311. [Google Scholar] [CrossRef]
- Rodriguez-Valle, M.; Vance, M.; Moolhuijzen, P.M.; Tao, X.; Lew-Tabor, A.E. Differential recognition by tick-resistant cattle of the recombinantly expressed Rhipicephalus microplus serine protease inhibitor-3 (RMS-3). Ticks Tick-Borne Dis. 2012, 3, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Valle, M.; Xu, T.; Kurscheid, S.; Lew-Tabor, A.E. Rhipicephalus microplus serine protease inhibitor family: Annotation, expression and functional characterisation assessment. Parasites Vectors 2015, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Schussler, G.C. The thyroxine-binding proteins. Thyroid 2000, 10, 141–149. [Google Scholar] [CrossRef]
- Lin, H.Y.; Muller, Y.A.; Hammond, G.L. Molecular and structural basis of steroid hormone binding and release from corticosteroid-binding globulin. Mol. Cell Endocrinol. 2010, 316, 3–12. [Google Scholar] [CrossRef]
- Kim, T.K.; Radulovic, Z.; Mulenga, A. Target validation of highly conserved Amblyomma americanum tick saliva serine protease inhibitor 19. Ticks Tick-Borne Dis. 2016, 7, 405–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mans, B.J. Tick histamine-binding proteins and related lipocalins: Potential as therapeutic agents. Curr. Opin. Investig. Drugs 2005, 6, 1131–1135. [Google Scholar]
- Mans, B.J.; Louw, A.I.; Neitz, A.W. Evolution of hematophagy in ticks: Common origins for blood coagulation and platelet aggregation inhibitors from soft ticks of the genus Ornithodoros. Mol. Biol. Evol. 2002, 19, 1695–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraganore, J.M.; Chao, B.; Joseph, M.L.; Jablonski, J.; Ramachandran, K.L. Anticoagulant activity of synthetic hirudin peptides. J. Biol. Chem. 1989, 264, 8692–8698. [Google Scholar] [CrossRef]
- Stepanova-Tresova, G.; Kopecky, J.; Kuthejlova, M. Identification of Borrelia burgdorferi sensu stricto, Borrelia garinii and Borrelia afzelii in Ixodes ricinus ticks from southern Bohemia using monoclonal antibodies. Zent. Bakteriol. Int. J. Med. Microbiol. 2000, 289, 797–806. [Google Scholar]
- Pospisilova, T.; Urbanova, V.; Hes, O.; Kopacek, P.; Hajdusek, O.; Sima, R. Tracking of Borrelia afzelii transmission from infected Ixodes ricinus nymphs to mice. Infect. Immun. 2019, 87, e00896-18. [Google Scholar] [CrossRef] [Green Version]
- Fong, S.W.; Kini, R.M.; Ng, L.F.P. Mosquito Saliva Reshapes Alphavirus Infection and Immunopathogenesis. J. Virol. 2018, 92, e01004-17. [Google Scholar] [CrossRef] [Green Version]
- Stibraniova, I.; Lahova, M.; Bartikova, P. Immunomodulators in tick saliva and their benefits. Acta Virol. 2013, 57, 200–216. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, D.; Cantaert, T.; Misse, D. Aedes Mosquito Salivary Components and Their Effect on the Immune Response to Arboviruses. Front. Cell Infect. Microbiol. 2020, 10, 407. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Wang, P.; Adusumilli, S.; Booth, C.J.; Narasimhan, S.; Anguita, J.; Fikrig, E. Antibodies against a tick protein, Salp15, protect mice from the Lyme disease agent. Cell Host Microbe 2009, 6, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopacek, P.; Zdychova, J.; Yoshiga, T.; Weise, C.; Rudenko, N.; Law, J.H. Molecular cloning, expression and isolation of ferritins from two tick species--Ornithodoros moubata and Ixodes ricinus. Insect Biochem. Mol. Biol. 2003, 33, 103–113. [Google Scholar] [CrossRef]
- Kyckova, K.; Kopecky, J. Effect of tick saliva on mechanisms of innate immune response against Borrelia afzelii. J. Med. Entomol. 2006, 43, 1208–1214. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; Arca, B.; Lombardo, F.; Calvo, E.; Phan, V.M.; Chandra, P.K.; Wikel, S.K. An annotated catalogue of salivary gland transcripts in the adult female mosquito, Aedes aegypti. BMC Genom. 2007, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, T.; Kroger, B.; Bialojan, S.; Lemaire, H.G.; Hoffken, H.W.; Reuschenbach, P.; Otte, M.; Dodt, J. A Kazal-type inhibitor with thrombin specificity from Rhodnius prolixus. J. Biol. Chem. 1993, 268, 16216–16222. [Google Scholar] [CrossRef]
- Lange, U.; Keilholz, W.; Schaub, G.A.; Landmann, H.; Markwardt, F.; Nowak, G. Biochemical characterization of a thrombin inhibitor from the bloodsucking bug Dipetalogaster maximus. Haemostasis 1999, 29, 204–211. [Google Scholar] [CrossRef]
- Bussacos, A.C.; Nakayasu, E.S.; Hecht, M.M.; Parente, J.A.; Soares, C.M.; Teixeira, A.R.; Almeida, I.C. Diversity of anti-haemostatic proteins in the salivary glands of Rhodnius species transmitters of Chagas disease in the greater Amazon. J. Proteom. 2011, 74, 1664–1672. [Google Scholar] [CrossRef]
- Cappello, M.; Bergum, P.W.; Vlasuk, G.P.; Furmidge, B.A.; Pritchard, D.I.; Aksoy, S. Isolation and characterization of the tsetse thrombin inhibitor: A potent antithrombotic peptide from the saliva of Glossina morsitans morsitans. Am. J. Trop. Med. Hyg. 1996, 54, 475–480. [Google Scholar] [CrossRef]
- Cappello, M.; Li, S.; Chen, X.; Li, C.B.; Harrison, L.; Narashimhan, S.; Beard, C.B.; Aksoy, S. Tsetse thrombin inhibitor: Bloodmeal-induced expression of an anticoagulant in salivary glands and gut tissue of Glossina morsitans morsitans. Proc. Natl. Acad. Sci. USA 1998, 95, 14290–14295. [Google Scholar] [CrossRef] [Green Version]
- Chagas, A.C.; Oliveira, F.; Debrabant, A.; Valenzuela, J.G.; Ribeiro, J.M.; Calvo, E. Lundep, a sand fly salivary endonuclease increases Leishmania parasite survival in neutrophils and inhibits XIIa contact activation in human plasma. PLoS Pathog. 2014, 10, e1003923. [Google Scholar] [CrossRef]
- Kato, H.; Gomez, E.A.; Fujita, M.; Ishimaru, Y.; Uezato, H.; Mimori, T.; Iwata, H.; Hashiguchi, Y. Ayadualin, a novel RGD peptide with dual antihemostatic activities from the sand fly Lutzomyia ayacuchensis, a vector of Andean-type cutaneous leishmaniasis. Biochimie 2015, 112, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraganore, J.M.; Bourdon, P.; Jablonski, J.; Ramachandran, K.L.; Fenton, J.W. 2nd, Design and characterization of hirulogs: A novel class of bivalent peptide inhibitors of thrombin. Biochemistry 1990, 29, 7095–7101. [Google Scholar] [CrossRef]
- USAN Council. List No. 374. New names. Bivalirudin. Desirudin. Clin. Pharmacol. Ther. 1995, 58, 241. [Google Scholar] [CrossRef]
- Poulin, R. Evolutionary Ecology of Parasites, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Schrodinger, LLC. The PyMOL Molecular Graphics System; Version 1.8; Schrodinger, LLC: New York, NY, USA, 2015. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protease | k2 (M−1 s−1) | ±SE |
|---|---|---|
| Plasmin | 225,064 | 14,183 |
| Trypsin | 29,447 | 3508 |
| Kallikrein | 16,682 | 1119 |
| fXIa | 16,328 | 948 |
| Thrombin | 13,794 | 1040 |
| fXIIa | 3324 | 409 |
| fXa | 2088 | 115 |
| APC | 523 | 35 |
| fVIIa + TF | 456 | 35 |
| fIXa | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotál, J.; Polderdijk, S.G.I.; Langhansová, H.; Ederová, M.; Martins, L.A.; Beránková, Z.; Chlastáková, A.; Hajdušek, O.; Kotsyfakis, M.; Huntington, J.A.; et al. Ixodes ricinus Salivary Serpin Iripin-8 Inhibits the Intrinsic Pathway of Coagulation and Complement. Int. J. Mol. Sci. 2021, 22, 9480. https://doi.org/10.3390/ijms22179480
Kotál J, Polderdijk SGI, Langhansová H, Ederová M, Martins LA, Beránková Z, Chlastáková A, Hajdušek O, Kotsyfakis M, Huntington JA, et al. Ixodes ricinus Salivary Serpin Iripin-8 Inhibits the Intrinsic Pathway of Coagulation and Complement. International Journal of Molecular Sciences. 2021; 22(17):9480. https://doi.org/10.3390/ijms22179480
Chicago/Turabian StyleKotál, Jan, Stéphanie G. I. Polderdijk, Helena Langhansová, Monika Ederová, Larissa A. Martins, Zuzana Beránková, Adéla Chlastáková, Ondřej Hajdušek, Michail Kotsyfakis, James A. Huntington, and et al. 2021. "Ixodes ricinus Salivary Serpin Iripin-8 Inhibits the Intrinsic Pathway of Coagulation and Complement" International Journal of Molecular Sciences 22, no. 17: 9480. https://doi.org/10.3390/ijms22179480