Colorectal Adenomas—Genetics and Searching for New Molecular Screening Biomarkers

 ,
,
Abstract
:1. Introduction
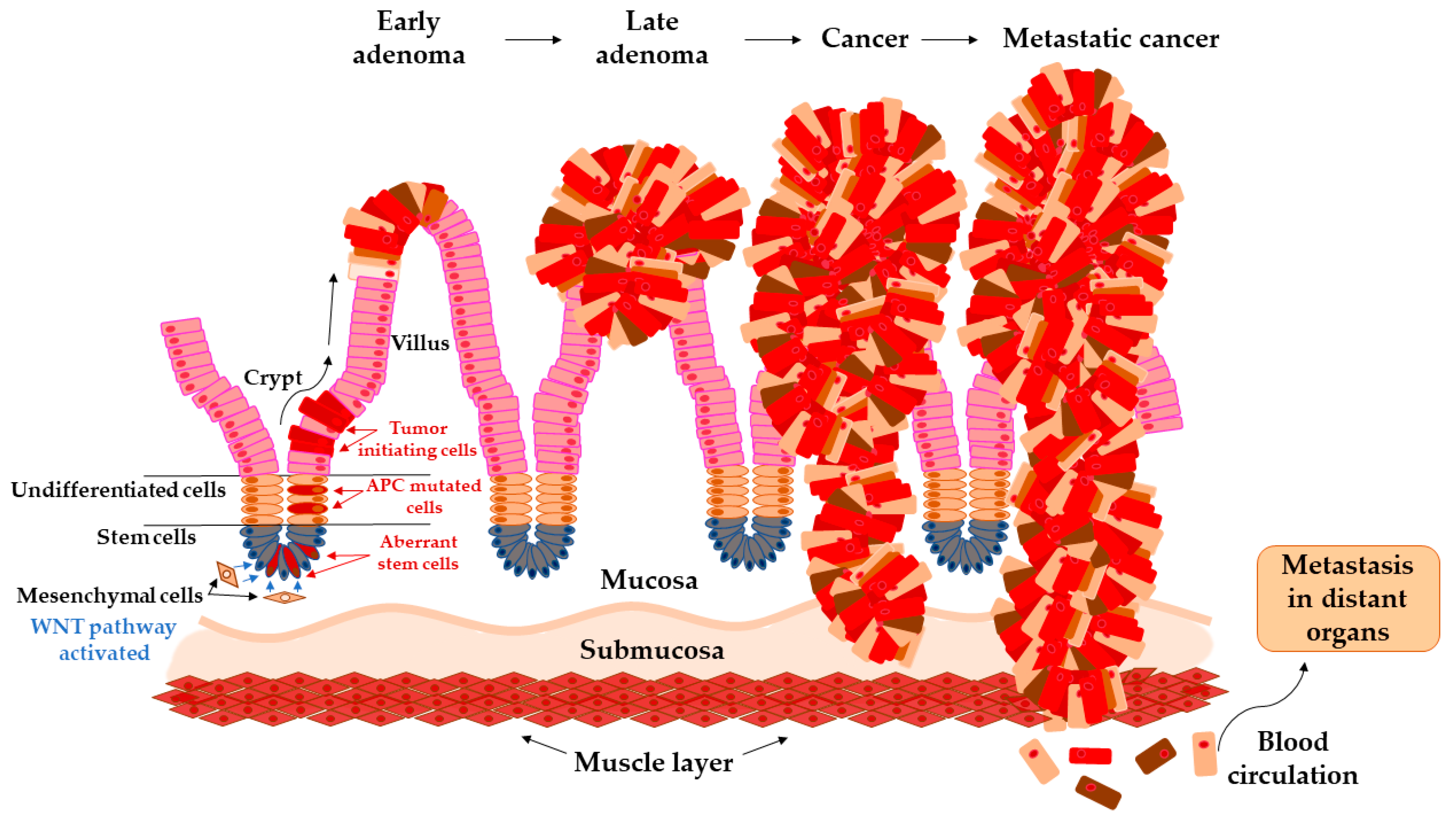
Transition of Adenoma to Carcinoma in Colon
2. Genetic Changes in Adenoma
2.1. Chromosomal Instability (CIN)
2.2. Microsatellite Instability (MSI)
2.3. DNA Methylation in CpG Islands
3. Insight into Novel Candidate Biomarkers of CRC
3.1. Long Non-Coding RNAs (lncRNAs)
3.2. MicroRNAs (miRNAs)
3.3. Circulating Cell-Free DNA
3.4. Circulating Tumor Cells (CTCs)
3.5. Circulating Inflammation Markers
3.6. Telomere Length
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALT | alternative lengthening of telomeres |
| aFAP | attenuated familial adenomatous polyposis |
| CC | colon cancer |
| CCE | colon capsule endoscopy |
| CCL20 | C-chemokine cysteine motif chemokine ligand 20 |
| cfDNA | cell-free DNA |
| CGH | comparative genomic hybridization |
| CIMP | CpG island methylator phenotype |
| CIN | chromosomal instability |
| CRC | colorectal cancer |
| CTCs | circulating tumor cells |
| CT | computed tomographic |
| ctDNA | circulating tumor DNA |
| EpCAM | epithelial cell adhesion molecule |
| FAP | familial adenomatous polyposis |
| FDA | Food and Drug Administration |
| FFPE | formalin-fixed paraffin-embedded |
| FIT | fecal immunochemical test |
| FOBT | fecal occult blood test |
| gFOBT | guaiac fecal occult blood test |
| Hb | hemoglobin |
| HNPCC | hereditary non-polyposis colorectal cancer |
| HPs | hyperplastic polyps |
| JPS | Juvenile polyposis syndrome |
| LOH | loss of heterozygosity |
| LS | Lynch syndrome |
| MAP | MUTYH-associated polyposis |
| MMR | mismatch repair pathway |
| mt-sDNA | multitarget stool DNA |
| ncRNA | non-coding RNA |
| NER | nucleotide excision repair genes |
| NGS | next-generation sequencing |
| NPV | negative predictive value |
| PHTS | hamartoma tumors syndrome |
| PJS | Peutz-Jeghers syndrome |
| PPAP | Polymerase proofreading-associated polyposis |
| PPV | positive predictive value |
| RC | rectal cancer |
| RLT | relative telomere length |
| SSA/Ps | sessile serrated polyps |
| UK | United Kingdom |
| USA | United States of America |
| WNT | Wingless/Int-1 pathway |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis Primers 2015, 1, 15065. [Google Scholar] [CrossRef] [Green Version]
- Pande, R.; Froggatt, P.; Baragwanath, P.; Harmston, C. Survival outcome of patients with screening versus symptomatically detected colorectal cancers. Colorectal Dis. 2013, 15, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Altobelli, E.; Rapacchietta, L.; Marziliano, C.; Campagna, G.; Profeta, V.F.; Fagnano, R. Differences in colorectal cancer surveillance epidemiology and screening in the WHO European Region. Oncol. Lett. 2019, 17, 2531–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Cancer Institute, Surveillance Research Program, Surveillance Systems Branch, Surveillance, Epidemiology, and End Results (SEER) Program. SEER*Stat Database: Incidence – SEER 18 Regs Research Data + Hurricane Katrina Impacted Louisiana Cases, Nov 2015 Sub (1973–2013 varying) – Linked To County Attributes – Total U.S., 1969–2014 Counties. 2016. Available online: https://seer.cancer.gov/data-software/documentation/seerstat/nov2016/ (accessed on 4 May 2020).
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Binefa, G.; Rodriguez-Moranta, F.; Teule, A.; Medina-Hayas, M. Colorectal cancer: From prevention to personalized medicine. World J. Gastroenterol. 2014, 20, 6786–6808. [Google Scholar] [CrossRef] [PubMed]
- Schreuders, E.H.; Grobbee, E.J.; Spaander, M.C.W.; Kuipers, E.J. Advances in Fecal Tests for Colorectal Cancer Screening. Curr. Treat. Options. Gastroenterol. 2016, 14, 52–162. [Google Scholar] [CrossRef] [Green Version]
- Mandel, J.S.; Bond, J.H.; Church, T.R.; Snover, D.C.; Bradley, G.M.; Schuman, L.M.; Ederer, F. Reducing Mortality from Colorectal Cancer by Screening for Fecal Occult Blood. N. Engl. J. Med. 1993, 328, 1365–1371. [Google Scholar] [CrossRef] [Green Version]
- Ladabaum, U.; Dominitz, J.A.; Kahi, C.; Schoen, R.E. Strategies for Colorectal Cancer Screening. Gastroenterology 2020, 158, 418–432. [Google Scholar] [CrossRef]
- Winawer, S.J.; Zauber, A.G.; Ho, M.N.; O’Brien, M.J.; Gottlieb, L.S.; Sternberg, S.S.; Waye, J.D.; Schapiro, M.; Bond, J.H.; Panish, J.F.; et al. Prevention of colorectal cancer by colonoscopic polypectomy. The National Polyp Study Workgroup. N. Engl. J. Med. 1993, 329, 1977–1981. [Google Scholar] [CrossRef]
- Lhewa, D.Y.; Strate, L.L. Pros and cons of colonoscopy in management of acute lower gastrointestinal bleeding. World J. Gastroenterol. 2012, 18, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Atkin, W.S.; Edwards, R.; Kralj-Hans, I.; Wooldrage, K.; Hart, A.R.; Northover, J.M.; Parkin, D.M.; Wardle, J.; Duffy, S.W.; Cuzick, J. Once-only flexible sigmoidoscopy screening in prevention of colorectal cancer: A multicentre randomised controlled trial. Lancet 2010, 375, 1624–1633. [Google Scholar] [CrossRef] [Green Version]
- Rokkas, T.; Papaxoinis, K.; Triantafyllou, K.; Ladas, S.D. A meta-analysis evaluating the accuracy of colon capsule endoscopy in detecting colon polyps. Gastrointest. Endosc. 2010, 71, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.D.; Chen, M.-H.; Toledano, A.Y.; Heiken, J.P.; Dachman, A.; Kuo, M.D.; Menias, C.O.; Siewert, B.; Cheema, J.I.; Obregon, R.G.; et al. Accuracy of CT Colonography for Detection of Large Adenomas and Cancers. N. Engl. J. Med. 2008, 359, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fei, F.; Zhang, M.; Li, Y.; Zhang, X.; Zhu, S.; Zhang, S. The role of (m)SEPT9 in screening, diagnosis, and recurrence monitoring of colorectal cancer. BMC Cancer 2019, 19, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Jia, J.; Peng, X.; Xiao, W.; Li, Y. The performance of the SEPT9 gene methylation assay and a comparison with other CRC screening tests: A meta-analysis. Sci. Rep. 2017, 7, 3032. [Google Scholar] [CrossRef] [Green Version]
- Rubio, C.A.; Jaramillo, E.; Lindblom, A.; Fogt, F. Classification of Colorectal Polyps: Guidelines for the Endoscopist. Endoscopy 2002, 34, 226–236. [Google Scholar] [CrossRef]
- Hassan, C.; Quintero, E.; Dumonceau, J.M.; Regula, J.; Brandao, C.; Chaussade, S.; Dekker, E.; Dinis-Ribeiro, M.; Ferlitsch, M.; Gimeno-Garcia, A.; et al. Post-polypectomy colonoscopy surveillance: European Society of Gastrointestinal Endoscopy (ESGE) Guideline. Endoscopy 2013, 45, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Levin, B.; Lieberman, D.A.; McFarland, B.; Smith, R.A.; Brooks, D.; Andrews, K.S.; Dash, C.; Giardiello, F.M.; Glick, S.; Levin, T.R.; et al. Screening and surveillance for the early detection of colorectal cancer and adenomatous polyps, 2008: A joint guideline from the American Cancer Society, the US Multi-Society Task Force on Colorectal Cancer, and the American College of Radiology. CA Cancer J. Clin. 2008, 58, 130–160. [Google Scholar] [CrossRef] [Green Version]
- Shussman, N.; Wexner, S.D. Colorectal polyps and polyposis syndromes. Gastroenterol. Rep. (Oxf) 2014, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.-H.; Huang, J.-Q.; Chen, J.-S. Interval colorectal cancer: A challenging field in colorectal cancer. Future Oncol. 2018, 14, 1307–1316. [Google Scholar] [CrossRef]
- Whiffin, N.; Hosking, F.J.; Farrington, S.M.; Palles, C.; Dobbins, S.E.; Zgaga, L.; Lloyd, A.; Kinnersley, B.; Gorman, M.; Tenesa, A.; et al. Identification of susceptibility loci for colorectal cancer in a genome-wide meta-analysis. Hum. Mol. Genet. 2014, 23, 4729–4737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archambault, A.N.; Su, Y.R.; Jeon, J.; Thomas, M.; Lin, Y.; Conti, D.V.; Win, A.K.; Sakoda, L.C.; Lansdorp-Vogelaar, I.; Peterse, E.F.P.; et al. Cumulative Burden of Colorectal Cancer-Associated Genetic Variants Is More Strongly Associated With Early-Onset vs. Late-Onset Cancer. Gastroenterology 2020, 158, 1274–1286.e12. [Google Scholar] [CrossRef]
- Brenner, H.; Hoffmeister, M.; Stegmaier, C.; Brenner, G.; Altenhofen, L.; Haug, U. Risk of progression of advanced adenomas to colorectal cancer by age and sex: Estimates based on 840,149 screening colonoscopies. Gut 2007, 56, 1585–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeve, F.; Boer, R.; Zauber, A.G.; Van Ballegooijen, M.; Van Oortmarssen, G.J.; Winawer, S.J.; Habbema, J.D. National Polyp Study data: Evidence for regression of adenomas. Int. J. Cancer 2004, 111, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Cross, W.; Kovac, M.; Mustonen, V.; Temko, D.; Davis, H.; Baker, A.M.; Biswas, S.; Arnold, R.; Chegwidden, L.; Gatenbee, C.; et al. The evolutionary landscape of colorectal tumorigenesis. Nat. Ecol. Evol. 2018, 2, 1661–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.F.; Ibrahim, A.E.; Arends, M.J. Molecular pathological classification of colorectal cancer. Virchows Arch. 2016, 469, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Pellino, G.; Gallo, G.; Pallante, P.; Capasso, R.; De Stefano, A.; Maretto, I.; Malapelle, U.; Qiu, S.; Nikolaou, S.; Barina, A.; et al. Noninvasive Biomarkers of Colorectal Cancer: Role in Diagnosis and Personalised Treatment Perspectives. Gastroenterol. Res. Pract. 2018, 2018, 2397863. [Google Scholar] [CrossRef]
- Winawer, S.J.; Zauber, A.G.; Fletcher, R.H.; Stillman, J.S.; O’Brien, M.J.; Levin, B.; Smith, R.A.; Lieberman, D.A.; Burt, R.W.; Levin, T.R.; et al. Guidelines for colonoscopy surveillance after polypectomy: A consensus update by the US Multi-Society Task Force on Colorectal Cancer and the American Cancer Society. CA Cancer J. Clin. 2006, 56, 143–159, quiz 184-5. [Google Scholar] [CrossRef]
- Cernat, L.; Blaj, C.; Jackstadt, R.; Brandl, L.; Engel, J.; Hermeking, H.; Jung, A.; Kirchner, T.; Horst, D. Colorectal Cancers Mimic Structural Organization of Normal Colonic Crypts. PLoS ONE 2014, 9, e104284. [Google Scholar] [CrossRef]
- Kinzler, K.W.; Vogelstein, B. Cancer-susceptibility genes. Gatekeepers and caretakers. Nature 1997, 386, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Tomasetti, C.; Vogelstein, B.; Parmigiani, G. Half or more of the somatic mutations in cancers of self-renewing tissues originate prior to tumor initiation. Proc. Natl. Acad. Sci. USA 2013, 110, 1999–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, E.; Fu, Y.; Colonna, V.; Mu, X.J.; Kang, H.M.; Lappalainen, T.; Sboner, A.; Lochovsky, L.; Chen, J.; Harmanci, A.; et al. Integrative annotation of variants from 1092 humans: Application to cancer genomics. Science 2013, 342, 1235587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozic, I.; Antal, T.; Ohtsuki, H.; Carter, H.; Kim, D.; Chen, S.; Karchin, R.; Kinzler, K.W.; Vogelstein, B.; Nowak, M.A. Accumulation of driver and passenger mutations during tumor progression. Proc. Natl. Acad. Sci. USA 2010, 107, 18545–18550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciriello, G.; Miller, M.L.; Aksoy, B.A.; Senbabaoglu, Y.; Schultz, N.; Sander, C. Emerging landscape of oncogenic signatures across human cancers. Nat. Genet. 2013, 45, 1127–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rowan, A.J.; Lamlum, H.; Ilyas, M.; Wheeler, J.; Straub, J.; Papadopoulou, A.; Bicknell, D.; Bodmer, W.F.; Tomlinson, I.P.M. APC mutations in sporadic colorectal tumors: A mutational “hotspot” and interdependence of the “two hits”. Proc. Natl. Acad. Sci. USA 2000, 97, 3352–3357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, L.N.; Dove, W.F. APC and its modifiers in colon cancer. Adv. Exp. Med. Biol. 2009, 656, 85–106. [Google Scholar]
- Burrell, R.A.; McClelland, S.E.; Endesfelder, D.; Groth, P.; Weller, M.C.; Shaikh, N.; Domingo, E.; Kanu, N.; Dewhurst, S.M.; Gronroos, E.; et al. Replication stress links structural and numerical cancer chromosomal instability. Nature 2013, 494, 492–496. [Google Scholar] [CrossRef] [Green Version]
- Sansregret, L.; Vanhaesebroeck, B.; Swanton, C. Determinants and clinical implications of chromosomal instability in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Tariq, K.; Ghias, K. Colorectal cancer carcinogenesis: A review of mechanisms. Cancer Bio. Med. 2016, 13, 120–135. [Google Scholar] [CrossRef] [Green Version]
- Hermsen, M.; Postma, C.; Baak, J.; Weiss, M.; Rapallo, A.; Sciutto, A.; Roemen, G.; Arends, J.W.; Williams, R.; Giaretti, W.; et al. Colorectal adenoma to carcinoma progression follows multiple pathways of chromosomal instability. Gastroenterology 2002, 123, 1109–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haan, J.C.; Labots, M.; Rausch, C.; Koopman, M.; Tol, J.; Mekenkamp, L.J.; van de Wiel, M.A.; Israeli, D.; van Essen, H.F.; van Grieken, N.C.; et al. Genomic landscape of metastatic colorectal cancer. Nat. Commun. 2014, 5, 5457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, C.R.; Goel, A. Microsatellite instability in colorectal cancer. Gastroenterology 2010, 138, 2073–2087.e3. [Google Scholar] [CrossRef] [PubMed]
- Peltomäki, P. Deficient DNA mismatch repair: A common etiologic factor for colon cancer. Hum. Mol. Gen. 2001, 10, 735–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loukola, A.; Salovaara, R.; Kristo, P.; Moisio, A.L.; Kääriäinen, H.; Ahtola, H.; Eskelinen, M.; Härkönen, N.; Julkunen, R.; Kangas, E.; et al. Microsatellite instability in adenomas as a marker for hereditary nonpolyposis colorectal cancer. Am. J. Pathol. 1999, 155, 1849–1853. [Google Scholar] [CrossRef] [Green Version]
- Steinke, V.; Engel, C.; Buttner, R.; Schackert, H.K.; Schmiegel, W.H.; Propping, P. Hereditary nonpolyposis colorectal cancer (HNPCC)/Lynch syndrome. Dtsch. Arztebl. Int. 2013, 110, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Hellmann, I.; Stadler, M.B.; Ramos, L.; Paabo, S.; Rebhan, M.; Schubeler, D. Distribution, silencing potential and evolutionary impact of promoter DNA methylation in the human genome. Nat. Genet. 2007, 39, 457–466. [Google Scholar] [CrossRef]
- Bird, A.P. CpG-rich islands and the function of DNA methylation. Nature 1986, 321, 209–213. [Google Scholar] [CrossRef]
- Kumar, S.; Chinnusamy, V.; Mohapatra, T. Epigenetics of Modified DNA Bases: 5-Methylcytosine and Beyond. Front. Genet. 2018, 9, 640. [Google Scholar] [CrossRef] [Green Version]
- Marcuello, M.; Vymetalkova, V.; Neves, R.P.L.; Duran-Sanchon, S.; Vedeld, H.M.; Tham, E.; van Dalum, G.; Flugen, G.; Garcia-Barberan, V.; Fijneman, R.J.; et al. Circulating biomarkers for early detection and clinical management of colorectal cancer. Mol. Aspects Med. 2019, 69, 107–122. [Google Scholar] [CrossRef]
- Toyota, M.; Ahuja, N.; Ohe-Toyota, M.; Herman, J.G.; Baylin, S.B.; Issa, J.-P.J. CpG island methylator phenotype in colorectal cancer. Proc. Natl. Acad. Sci. USA 1999, 96, 8681–8686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psofaki, V.; Kalogera, C.; Tzambouras, N.; Stephanou, D.; Tsianos, E.; Seferiadis, K.; Kolios, G. Promoter methylation status of hMLH1, MGMT, and CDKN2A/p16 in colorectal adenomas. World J. Gastroenterol. 2010, 16, 3553–3560. [Google Scholar] [CrossRef] [PubMed]
- Rashid, A.; Shen, L.; Morris, J.S.; Issa, J.-P.J.; Hamilton, S.R. CpG Island Methylation in Colorectal Adenomas. Am. J. Pathol. 2001, 159, 1129–1135. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-J.; Rashid, A.; Lee, J.-H.; Kim, S.G.; Hamilton, S.R.; Wu, T.-T. Frequent CpG Island Methylation in Serrated Adenomas of the Colorectum. Am. J. Pathol. 2003, 162, 815–822. [Google Scholar] [CrossRef]
- Karen Curtin, M.L.S.; Wade, S. Samowitz CpG Island Methylation in Colorectal Cancer: Past, Present and Future. Pathol. Res. Inter. 2011, 2011, 8. [Google Scholar]
- Nazemalhosseini Mojarad, E.; Kuppen, P.J.; Aghdaei, H.A.; Zali, M.R. The CpG island methylator phenotype (CIMP) in colorectal cancer. Gastroenterol. Hepatol. Bed. Bench. 2013, 6, 120–128. [Google Scholar]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef]
- An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [CrossRef]
- Siddiqui, H.; Al-Ghafari, A.; Choudhry, H.; Al Doghaither, H. Roles of long non-coding RNAs in colorectal cancer tumorigenesis: A Review. Mol. Clin. Oncol. 2019, 11, 167–172. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Z.; Wang, X.; Huang, Z.; He, Z.; Chen, Y. Long non-coding RNA: A new player in cancer. J. Hematol. Oncol. 2013, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Galamb, O.; Barták, B.K.; Kalmár, A.; Nagy, Z.B.; Szigeti, K.A.; Tulassay, Z.; Igaz, P.; Molnár, B. Diagnostic and prognostic potential of tissue and circulating long non-coding RNAs in colorectal tumors. World J. Gastroenterol. 2019, 25, 5026–5048. [Google Scholar] [CrossRef]
- Svoboda, M.; Slyskova, J.; Schneiderova, M.; Makovicky, P.; Bielik, L.; Levy, M.; Lipska, L.; Hemmelova, B.; Kala, Z.; Protivankova, M.; et al. HOTAIR long non-coding RNA is a negative prognostic factor not only in primary tumors, but also in the blood of colorectal cancer patients. Carcinogenesis 2014, 35, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Alaiyan, B.; Ilyayev, N.; Stojadinovic, A.; Izadjoo, M.; Roistacher, M.; Pavlov, V.; Tzivin, V.; Halle, D.; Pan, H.; Trink, B.; et al. Differential expression of colon cancer associated transcript1 (CCAT1) along the colonic adenoma-carcinoma sequence. BMC Cancer 2013, 13, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Tan, X.; Wang, X.; Jin, H.; Liu, L.; Ma, L.; Yu, H.; Fan, Z. C-Myc-activated long noncoding RNA CCAT1 promotes colon cancer cell proliferation and invasion. Tumor Biology 2014, 35, 12181–12188. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yu, Y.; Xu, B.; Zhang, M.; Li, Q.; Miao, L. Pivotal prognostic and diagnostic role of the long noncoding RNA colon cancerassociated transcript 1 expression in human cancer (Review). Mol. Med. Rep. 2019, 19, 771–782. [Google Scholar] [PubMed] [Green Version]
- Tsang, W.P.; Ng, E.K.O.; Ng, S.S.M.; Jin, H.; Yu, J.; Sung, J.J.Y.; Kwok, T.T. Oncofetal H19-derived miR-675 regulates tumor suppressor RB in human colorectal cancer. Carcinogenesis 2009, 31, 350–358. [Google Scholar] [CrossRef]
- Ohtsuka, M.; Ling, H.; Ivan, C.; Pichler, M.; Matsushita, D.; Goblirsch, M.; Stiegelbauer, V.; Shigeyasu, K.; Zhang, X.; Chen, M.; et al. H19 Noncoding RNA, an Independent Prognostic Factor, Regulates Essential Rb-E2F and CDK8-beta-Catenin Signaling in Colorectal Cancer. Ebio. Med. 2016, 13, 113–124. [Google Scholar]
- Yoruker, E.E.; Keskin, M.; Kulle, C.B.; Holdenrieder, S.; Gezer, U. Diagnostic and prognostic value of circulating lncRNA H19 in gastric cancer. Biomed. Rep. 2018, 9, 181–186. [Google Scholar]
- Yoshimizu, T.; Miroglio, A.; Ripoche, M.A.; Gabory, A.; Vernucci, M.; Riccio, A.; Colnot, S.; Godard, C.; Terris, B.; Jammes, H.; et al. The H19 locus acts in vivo as a tumor suppressor. Proc. Natl. Acad. Sci. USA 2008, 105, 12417–12422. [Google Scholar] [CrossRef] [Green Version]
- Graham, L.D.; Pedersen, S.K.; Brown, G.S.; Ho, T.; Kassir, Z.; Moynihan, A.T.; Vizgoft, E.K.; Dunne, R.; Pimlott, L.; Young, G.P.; et al. Colorectal Neoplasia Differentially Expressed (CRNDE), a Novel Gene with Elevated Expression in Colorectal Adenomas and Adenocarcinomas. Gen. Cancer 2011, 2, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, X.; Gao, S.; Jing, F.; Yang, Y.; Du, L.; Zheng, G.; Li, P.; Li, C.; Wang, C. Exosomal long noncoding RNA CRNDE-h as a novel serum-based biomarker for diagnosis and prognosis of colorectal cancer. Oncotarget 2016, 7, 85551–85563. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, Y.; Ai, M.; Wang, H.; Duan, Z.; Wang, H.; Zhao, L.; Yu, J.; Ding, Y.; Wang, S. Long Long noncoding RNA CRNDE stabilized by hnRNPUL2 accelerates cell proliferation and migration in colorectal carcinoma via activating Ras/MAPK signaling pathways. Cell Death Dis. 2017, 8, e2862. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Ye, X.; Du, Q.; Zhu, B.; Zhai, Q.; Li, X.X. The Long Non-Coding RNA CRNDE Promotes Colorectal Carcinoma Progression by Competitively Binding miR-217 with TCF7L2 and Enhancing the Wnt/beta-Catenin Signaling Pathway. Cell. Physiol. Biochem. 2017, 41, 2489–2502. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Li, B. Role of miRNA in transformation from normal tissue to colorectal adenoma and cancer. J. Cancer Res. Ther. 2019, 15, 278–285. [Google Scholar] [PubMed]
- Vymetalkova, V.; Vodicka, P.; Vodenkova, S.; Alonso, S.; Schneider-Stock, R. DNA methylation and chromatin modifiers in colorectal cancer. Mol. Aspects. Med. 2019, 69, 73–92. [Google Scholar] [CrossRef] [PubMed]
- Tsikitis, V.L.; Potter, A.; Mori, M.; Buckmeier, J.A.; Preece, C.R.; Harrington, C.A.; Bartley, A.N.; Bhattacharyya, A.K.; Hamilton, S.R.; Lance, M.P.; et al. MicroRNA Signatures of Colonic Polyps on Screening and Histology. Cancer Prev. Res. 2016, 9, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Kanth, P.; Hazel, M.W.; Boucher, K.M.; Yang, Z.; Wang, L.; Bronner, M.P.; Boylan, K.E.; Burt, R.W.; Westover, M.; Neklason, D.W.; et al. Small RNA sequencing of sessile serrated polyps identifies microRNA profile associated with colon cancer. Genes Chromosomes Cancer 2019, 58, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.I.; Hussein, S.; West, K.; Singh, B.; Jameson, J.S.; Pringle, J.H. MicroRNAs associated with initiation and progression of colonic polyp: A feasibility study. Int. J. Surg. 2015, 13, 272–279. [Google Scholar] [CrossRef]
- Oberg, A.L.; French, A.J.; Sarver, A.L.; Subramanian, S.; Morlan, B.W.; Riska, S.M.; Borralho, P.M.; Cunningham, J.M.; Boardman, L.A.; Wang, L.; et al. miRNA expression in colon polyps provides evidence for a multihit model of colon cancer. PLoS ONE 2011, 6, e20465. [Google Scholar] [CrossRef]
- Ito, M.; Mitsuhashi, K.; Igarashi, H.; Nosho, K.; Naito, T.; Yoshii, S.; Takahashi, H.; Fujita, M.; Sukawa, Y.; Yamamoto, E.; et al. MicroRNA-31 expression in relation to BRAF mutation, CpG island methylation and colorectal continuum in serrated lesions. Int. J. Cancer 2014, 135, 2507–2515. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Ren, L.L.; Zheng, P.; Zheng, H.M.; Yu, Y.N.; Wang, J.L.; Lin, Y.W.; Chen, Y.X.; Ge, Z.Z.; Chen, X.Y.; et al. miR-194 as a predictor for adenoma recurrence in patients with advanced colorectal adenoma after polypectomy. Cancer. Prev. Res. (Phila) 2014, 7, 607–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardila, H.J.; Sanabria-Salas, M.C.; Meneses, X.; Rios, R.; Huertas-Salgado, A.; Serrano, M.L. Circulating miR-141-3p, miR-143-3p and miR-200c-3p are differentially expressed in colorectal cancer and advanced adenomas. Mol. Clin. Oncol. 2019, 11, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, Z.B.; Wichmann, B.; Kalmar, A.; Galamb, O.; Bartak, B.K.; Spisak, S.; Tulassay, Z.; Molnar, B. Colorectal adenoma and carcinoma specific miRNA profiles in biopsy and their expression in plasma specimens. Clin. Epigenetics 2017, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Uratani, R.; Toiyama, Y.; Kitajima, T.; Kawamura, M.; Hiro, J.; Kobayashi, M.; Tanaka, K.; Inoue, Y.; Mohri, Y.; Mori, T.; et al. Diagnostic Potential of Cell-Free and Exosomal MicroRNAs in the Identification of Patients with High-Risk Colorectal Adenomas. PLoS ONE 2016, 11, e0160722. [Google Scholar] [CrossRef]
- Yau, T.O.; Tang, C.M.; Harriss, E.K.; Dickins, B.; Polytarchou, C. Faecal microRNAs as a non-invasive tool in the diagnosis of colonic adenomas and colorectal cancer: A meta-analysis. Sci. Rep. 2019, 9, 9491. [Google Scholar] [CrossRef]
- Wu, C.W.; Ng, S.C.; Dong, Y.; Tian, L.; Ng, S.S.; Leung, W.W.; Law, W.T.; Yau, T.O.; Chan, F.K.; Sung, J.J.; et al. Identification of microRNA-135b in stool as a potential noninvasive biomarker for colorectal cancer and adenoma. Clin. Cancer Res. 2014, 20, 2994–3002. [Google Scholar] [CrossRef] [Green Version]
- Schetter, A.J.; Leung, S.Y.; Sohn, J.J.; Zanetti, K.A.; Bowman, E.D.; Yanaihara, N.; Yuen, S.T.; Chan, T.L.; Kwong, D.L.; Au, G.K.; et al. MicroRNA expression profiles associated with prognosis and therapeutic outcome in colon adenocarcinoma. Jama 2008, 299, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Diosdado, B.; van de Wiel, M.A.; Terhaar Sive Droste, J.S.; Mongera, S.; Postma, C.; Meijerink, W.J.; Carvalho, B.; Meijer, G.A. MiR-17-92 cluster is associated with 13q gain and c-myc expression during colorectal adenoma to adenocarcinoma progression. Br. J. Cancer 2009, 101, 707–714. [Google Scholar] [CrossRef]
- Kanaan, Z.; Roberts, H.; Eichenberger, M.R.; Billeter, A.; Ocheretner, G.; Pan, J.; Rai, S.N.; Jorden, J.; Williford, A.; Galandiuk, S. A plasma microRNA panel for detection of colorectal adenomas: A step toward more precise screening for colorectal cancer. Ann. Surg. 2013, 258, 400–408. [Google Scholar] [CrossRef]
- Tsikitis, V.L.; White, I.; Mori, M.; Potter, A.; Bhattcharyya, A.; Hamilton, S.R.; Buckmeier, J.; Lance, P.; Thompson, P. Differential expression of microRNA-320a, -145, and -192 along the continuum of normal mucosa to high-grade dysplastic adenomas of the colorectum. Am. J. Surg. 2014, 207, 717–722, discussion 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Du, L.; Yang, X.; Zhang, X.; Wang, L.; Yang, Y.; Li, J.; Wang, C. Serum microRNA panel as biomarkers for early diagnosis of colorectal adenocarcinoma. Br. J. Cancer 2014, 111, 1985–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Groen, F.L.; Timmer, L.M.; Menezes, R.X.; Diosdado, B.; Hooijberg, E.; Meijer, G.A.; Steenbergen, R.D.; Carvalho, B. Oncogenic Role of miR-15a-3p in 13q Amplicon-Driven Colorectal Adenoma-to-Carcinoma Progression. PLoS ONE 2015, 10, e0132495. [Google Scholar] [CrossRef] [PubMed]
- Gattolliat, C.H.; Uguen, A.; Pesson, M.; Trillet, K.; Simon, B.; Doucet, L.; Robaszkiewicz, M.; Corcos, L. MicroRNA and targeted mRNA expression profiling analysis in human colorectal adenomas and adenocarcinomas. Eur. J. Cancer 2015, 51, 409–420. [Google Scholar] [CrossRef]
- Hibino, Y.; Sakamoto, N.; Naito, Y.; Goto, K.; Oo, H.Z.; Sentani, K.; Hinoi, T.; Ohdan, H.; Oue, N.; Yasui, W. Significance of miR-148a in Colorectal Neoplasia: Downregulation of miR-148a Contributes to the Carcinogenesis and Cell Invasion of Colorectal Cancer. Pathobiology 2015, 82, 233–241. [Google Scholar] [CrossRef]
- Ho, G.Y.F.; Jung, H.J.; Schoen, R.E.; Wang, T.; Lin, J.; Williams, Z.; Weissfeld, J.L.; Park, J.Y.; Loudig, O.; Suh, Y. Differential expression of circulating microRNAs according to severity of colorectal neoplasia. Transl. Res. 2015, 166, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Tadano, T.; Kakuta, Y.; Hamada, S.; Shimodaira, Y.; Kuroha, M.; Kawakami, Y.; Kimura, T.; Shiga, H.; Endo, K.; Masamune, A.; et al. MicroRNA-320 family is downregulated in colorectal adenoma and affects tumor proliferation by targeting CDK6. World J. Gastrointest. Oncol. 2016, 8, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, M.; Ding, Y.; Fan, Z.; Zhang, J.; Zhang, H.; Jiang, B.; Zhu, Y. Serum MicroRNA profile in patients with colon adenomas or cancer. BMC Med. Genomics 2017, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Raju, G.S.; Chang, D.W.; Lin, S.H.; Chen, Z.; Wu, X. Global and targeted circulating microRNA profiling of colorectal adenoma and colorectal cancer. Cancer 2018, 124, 785–796. [Google Scholar] [CrossRef]
- Aherne, S.T.; Madden, S.F.; Hughes, D.J.; Pardini, B.; Naccarati, A.; Levy, M.; Vodicka, P.; Neary, P.; Dowling, P.; Clynes, M. Circulating miRNAs miR-34a and miR-150 associated with colorectal cancer progression. BMC Cancer 2015, 15, 329. [Google Scholar] [CrossRef] [Green Version]
- Crowley, E.; Di Nicolantonio, F.; Loupakis, F.; Bardelli, A. Liquid biopsy: Monitoring cancer-genetics in the blood. Nat. Rev. Clin. Oncol. 2013, 10, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Jiraskova, K.; Hughes, D.J.; Brezina, S.; Gumpenberger, T.; Veskrnova, V.; Buchler, T.; Schneiderova, M.; Levy, M.; Liska, V.; Vodenkova, S.; et al. Functional Polymorphisms in DNA Repair Genes Are Associated with Sporadic Colorectal Cancer Susceptibility and Clinical Outcome. Int J. Mol. Sci. 2018, 20, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervena, K.; Vodicka, P.; Vymetalkova, V. Diagnostic and prognostic impact of cell-free DNA in human cancers: Systematic review. Mutat. Res. 2019, 781, 100–129. [Google Scholar] [CrossRef] [PubMed]
- Myint, N.N.M.; Verma, A.M.; Fernandez-Garcia, D.; Sarmah, P.; Tarpey, P.S.; Al-Aqbi, S.S.; Cai, H.; Trigg, R.; West, K.; Howells, L.M.; et al. Circulating tumor DNA in patients with colorectal adenomas: Assessment of detectability and genetic heterogeneity. Cell Death Dis. 2018, 9, 894. [Google Scholar] [CrossRef]
- Aravanis, A.M.; Lee, M.; Klausner, R.D. Next-Generation Sequencing of Circulating Tumor DNA for Early Cancer Detection. Cell 2017, 168, 571–574. [Google Scholar] [CrossRef] [Green Version]
- Bettegowda, C.; Sausen, M.; Leary, R.J.; Kinde, I.; Wang, Y.; Agrawal, N.; Bartlett, B.R.; Wang, H.; Luber, B.; Alani, R.M.; et al. Detection of circulating tumor DNA in early- and late-stage human malignancies. Sci. Transl. Med. 2014, 6, 224ra24. [Google Scholar] [CrossRef] [Green Version]
- Mead, R.; Duku, M.; Bhandari, P.; Cree, I.A. Circulating tumour markers can define patients with normal colons, benign polyps, and cancers. Br. J. Cancer. 2011, 105, 239–245. [Google Scholar] [CrossRef]
- Galanopoulos, M.; Papanikolaou, I.; Zografos, E.; Viazis, N.; Papatheodoridis, G.; Karamanolis, D.; Marinos, E.; Mantzaris, G.; Gazouli, M. Comparative Study of Mutations in Single Nucleotide Polymorphism Loci of KRAS and BRAF Genes in Patients Who Underwent Screening Colonoscopy, With and Without Premalignant Intestinal Polyps. Anticancer Res. 2017, 37, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Kopreski, M.S.; Benko, F.A.; Borys, D.J.; Khan, A.; McGarrity, T.J.; Gocke, C.D. Somatic Mutation Screening: Identification of Individuals Harboring K-ras Mutations With the Use of Plasma DNA. JNCI: J. Natl. Cancer Inst. 2000, 92, 918–923. [Google Scholar] [CrossRef] [Green Version]
- Perrone, F.; Lampis, A.; Bertan, C.; Verderio, P.; Ciniselli, C.M.; Pizzamiglio, S.; Frattini, M.; Nucifora, M.; Molinari, F.; Gallino, G.; et al. Circulating Free DNA in a Screening Program for Early Colorectal Cancer Detection. Tumori 2014, 100, 115–121. [Google Scholar] [CrossRef]
- Gausachs, M.; Borras, E.; Chang, K.; Gonzalez, S.; Azuara, D.; Delgado Amador, A.; Lopez-Doriga, A.; San Lucas, F.A.; Sanjuan, X.; Paules, M.J.; et al. Mutational Heterogeneity in APC and KRAS Arises at the Crypt Level and Leads to Polyclonality in Early Colorectal Tumorigenesis. Clin. Cancer Res. 2017, 23, 5936–5947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirlwell, C.; Will, O.C.; Domingo, E.; Graham, T.A.; McDonald, S.A.; Oukrif, D.; Jeffrey, R.; Gorman, M.; Rodriguez-Justo, M.; Chin-Aleong, J.; et al. Clonality assessment and clonal ordering of individual neoplastic crypts shows polyclonality of colorectal adenomas. Gastroenterology 2010, 138, 1441–1454.e7. [Google Scholar] [CrossRef] [PubMed]
- Fleshner, P.; Braunstein, G.D.; Ovsepyan, G.; Tonozzi, T.R.; Kammesheidt, A. Tumor-associated DNA mutation detection in individuals undergoing colonoscopy. Cancer Med. 2018, 7, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Fleischhacker, M.; Schmidt, B. Circulating nucleic acids (CNAs) and cancer—A survey. Biochim. Biophys. Acta. 2007, 1775, 181–232. [Google Scholar] [CrossRef]
- Martincorena, I.; Roshan, A.; Gerstung, M.; Ellis, P.; Van Loo, P.; McLaren, S.; Wedge, D.C.; Fullam, A.; Alexandrov, L.B.; Tubio, J.M.; et al. Tumor evolution. High burden and pervasive positive selection of somatic mutations in normal human skin. Science 2015, 348, 880–886. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Keohavong, P.; Finkelstein, S.D.; Swalsky, P.; Bakker, A.; Weissfeld, J.; Srivastava, S.; Whiteside, T.L. K-ras gene mutations in normal colorectal tissues from K-ras mutation-positive colorectal cancer patients. Cancer Res. 1997, 57, 2485–2492. [Google Scholar]
- Takayama, T.; Katsuki, S.; Takahashi, Y.; Ohi, M.; Nojiri, S.; Sakamaki, S.; Kato, J.; Kogawa, K.; Miyake, H.; Niitsu, Y. Aberrant crypt foci of the colon as precursors of adenoma and cancer. N. Engl. J. Med. 1998, 339, 1277–1284. [Google Scholar] [CrossRef]
- Tobi, M.; Luo, F.C.; Ronai, Z. Detection of K-ras mutation in colonic effluent samples from patients without evidence of colorectal carcinoma. J. Natl. Cancer Inst. 1994, 86, 1007–1010. [Google Scholar]
- Chen, A.; Braunstein, G.; Anselmo, M.; Jaboni, J.; Viloria, F.; Neidich, J.; Li, X.; Kammesheidt, A. Mutation detection with a liquid biopsy 96 mutation assay in cancer patients and healthy donors. Cancer Trans. Med. 2017, 3, 39. [Google Scholar]
- Gocke, C.D.; Benko, F.A.; Kopreski, M.S.; McGarrity, T.J. p53 and APC mutations are detectable in the plasma and serum of patients with colorectal cancer (CRC) or adenomas. Ann. N. Y. Acad. Sci. 2000, 906, 44–50. [Google Scholar] [CrossRef]
- Berger, B.M.; Ahlquist, D.A. Stool DNA screening for colorectal neoplasia: Biological and technical basis for high detection rates. Pathology 2012, 44, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Lidgard, G.P.; Domanico, M.J.; Bruinsma, J.J.; Light, J.; Gagrat, Z.D.; Oldham–Haltom, R.L.; Fourrier, K.D.; Allawi, H.; Yab, T.C.; Taylor, W.R.; et al. Clinical Performance of an Automated Stool DNA Assay for Detection of Colorectal Neoplasia. Clin. Gastroenterol. Hepatol. 2013, 11, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Imperiale, T.F.; Ransohoff, D.F.; Itzkowitz, S.H.; Levin, T.R.; Lavin, P.; Lidgard, G.P.; Ahlquist, D.A.; Berger, B.M. Multitarget Stool DNA Testing for Colorectal-Cancer Screening. N. Engl. J. Med. 2014, 370, 1287–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardingham, J.E.; Hewett, P.J.; Sage, R.E.; Finch, J.L.; Nuttall, J.D.; Kotasek, D.; Dobrovic, A. Molecular detection of blood-borne epithelial cells in colorectal cancer patients and in patients with benign bowel disease. Int. J. Cancer 2000, 89, 8–13. [Google Scholar] [CrossRef]
- Racila, E.; Euhus, D.; Weiss, A.J.; Rao, C.; McConnell, J.; Terstappen, L.W.; Uhr, J.W. Detection and characterization of carcinoma cells in the blood. Proc. Natl. Acad. Sci. USA 1998, 95, 4589–4594. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.M.; Ramani, V.C.; Jeffrey, S.S. Circulating tumor cell technologies. Mol. Oncol. 2016, 10, 374–394. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.H.S.; Becker, T.M.; Chua, W.; Ng, W.L.; de Souza, P.; Spring, K.J. Circulating tumour cells and the epithelial mesenchymal transition in colorectal cancer. J. Clin. Pathol. 2014, 67, 848–853. [Google Scholar] [CrossRef]
- Sotelo, M.J.; Sastre, J.; Maestro, M.L.; Veganzones, S.; Vieitez, J.M.; Alonso, V.; Gravalos, C.; Escudero, P.; Vera, R.; Aranda, E.; et al. Role of circulating tumor cells as prognostic marker in resected stage III colorectal cancer. Ann. Oncol. 2015, 26, 535–541. [Google Scholar] [CrossRef]
- Tan, Y.; Wu, H. The significant prognostic value of circulating tumor cells in colorectal cancer: A systematic review and meta-analysis. Cur. Prob. Cancer. 2018, 42, 95–106. [Google Scholar] [CrossRef]
- Huang, X.; Gao, P.; Song, Y.; Sun, J.; Chen, X.; Zhao, J.; Liu, J.; Xu, H.; Wang, Z. Relationship between circulating tumor cells and tumor response in colorectal cancer patients treated with chemotherapy: A meta-analysis. BMC Cancer 2014, 14, 976. [Google Scholar] [CrossRef]
- Guadagni, S.; Fiorentini, G.; De Simone, M.; Masedu, F.; Zoras, O.; Mackay, A.R.; Sarti, D.; Papasotiriou, I.; Apostolou, P.; Catarci, M.; et al. Precision oncotherapy based on liquid biopsies in multidisciplinary treatment of unresectable recurrent rectal cancer: A retrospective cohort study. J. Cancer Res. Clin. Oncol. 2020, 146, 205–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guadagni, S.; Clementi, M.; Mackay, A.R.; Ricevuto, E.; Fiorentini, G.; Sarti, D.; Palumbo, P.; Apostolou, P.; Papasotiriou, I.; Masedu, F.; et al. Real-life multidisciplinary treatment for unresectable colorectal cancer liver metastases including hepatic artery infusion with chemo-filtration and liquid biopsy precision oncotherapy: Observational cohort study. J. Cancer Res. Clin. Oncol. 2020, 146, 1273–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, W.-S.; You, J.-F.; Hung, H.-Y.; Hsieh, P.-S.; Hsieh, B.; Lenz, H.-J.; Idos, G.; Friedland, S.; Yi-Jiun Pan, J.; Shao, H.-J.; et al. Novel Circulating Tumor Cell Assay for Detection of Colorectal Adenomas and Cancer. Clin. Trans. Gastroenterol. 2019, 10, e00088. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-Y.; Berndt, S.I.; Shiels, M.S.; Katki, H.A.; Chaturvedi, A.K.; Wentzensen, N.; Trabert, B.; Kemp, T.J.; Pinto, L.A.; Hildesheim, A.; et al. Circulating inflammation markers and colorectal adenoma risk. Carcinogenesis 2019, 40, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Biondi, A.; Galvano, F.; Basile, F.; Sciacca, S.; Giovannucci, E.L.; Grosso, G. Markers of systemic inflammation and colorectal adenoma risk: Meta-analysis of observational studies. World J. Gastroenterol. 2017, 23, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Basavaraju, U.; Shebl, F.M.; Palmer, A.J.; Berry, S.; Hold, G.L.; El-Omar, E.M.; Rabkin, C.S. Cytokine gene polymorphisms, cytokine levels and the risk of colorectal neoplasia in a screened population of Northeast Scotland. Eur. J. Cancer. Prev. 2015, 24, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.J.; Sedjo, R.L.; Rozhok, A.; Salstrom, J.; Ahnen, D.; Levin, T.R.; D’Agostino, R.; Haffner, S.; DeGregori, J.; Byers, T. Lack of significant association between serum inflammatory cytokine profiles and the presence of colorectal adenoma. BMC Cancer 2015, 15, 123. [Google Scholar] [CrossRef] [Green Version]
- Murphy, N.; Cross, A.J.; Huang, W.-Y.; Rajabzadeh-Heshejin, V.; Stanczyk, F.; Hayes, R.; Gunter, M.J. A prospective evaluation of C-peptide levels and colorectal adenoma incidence. Cancer Epidemiology 2015, 39, 160–165. [Google Scholar] [CrossRef]
- Comstock, S.S.; Xu, D.; Hortos, K.; Kovan, B.; McCaskey, S.; Pathak, D.R.; Fenton, J.I. Association of serum cytokines with colorectal polyp number and type in adult males. Eur. J. Cancer. Prev. 2016, 25, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Peacock, S.D.; Massey, T.E.; Vanner, S.J.; King, W.D. Telomere length in the colon is related to colorectal adenoma prevalence. PLoS ONE 2018, 13, e0205697. [Google Scholar] [CrossRef]
- Kroupa, M.; Rachakonda, S.K.; Liska, V.; Srinivas, N.; Urbanova, M.; Jiraskova, K.; Schneiderova, M.; Vycital, O.; Vymetalkova, V.; Vodickova, L.; et al. Relationship of telomere length in colorectal cancer patients with cancer phenotype and patient prognosis. Br. J. Cancer 2019, 121, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Tomasova, K.; Kroupa, M.; Forsti, A.; Vodicka, P.; Vodickova, L. Telomere maintenance in interplay with DNA repair in pathogenesis and treatment of colorectal cancer. Mutagenesis 2020, geaa005. [Google Scholar] [CrossRef]
- Zöchmeister, C.; Brezina, S.; Hofer, P.; Baierl, A.; Bergmann, M.M.; Bachleitner-Hofmann, T.; Karner-Hanusch, J.; Stift, A.; Gerger, A.; Leeb, G.; et al. Leukocyte telomere length throughout the continuum of colorectal carcinogenesis. Oncotarget 2018, 9, 13582–13592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valls Bautista, C.; Pinol Felis, C.; Rene Espinet, J.M.; Buenestado Garcia, J.; Vinas Salas, J. Telomerase activity and telomere length in the colorectal polyp-carcinoma sequence. Rev. Esp. Enferm. Dig. 2009, 101, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roger, L.; Jones, R.E.; Heppel, N.H.; Williams, G.T.; Sampson, J.R.; Baird, D.M. Extensive Telomere Erosion in the Initiation of Colorectal Adenomas and Its Association With Chromosomal Instability. J. Natl. Cancer Inst. 2013, 105, 1202–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.R.; Kim, Y.J.; Kim, H.J.; Kim, S.K.; Lee, J.H. Telomere length changes in colorectal cancers and polyps. J. Korean Med. Sci. 2002, 17, 360–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suraweera, N.; Mouradov, D.; Li, S.; Jorissen, R.N.; Hampson, D.; Ghosh, A.; Sengupta, N.; Thaha, M.; Ahmed, S.; Kirwan, M.; et al. Relative telomere lengths in tumor and normal mucosa are related to disease progression and chromosome instability profiles in colorectal cancer. Oncotarget 2016, 7, 36474–36488. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Non-Neoplastic | Neoplastic | |||
|---|---|---|---|---|
| Sporadic | Hereditary | Sporadic | Hereditary | |
| Hyperplastic polyps | Hyperplastic polyposis | Benigns adenomas: | Tubular | Familial adenomatous polyposis (FAP) |
| Villous | ||||
| Tubulovillous | ||||
| Inflammatory polyps | Juvenile polyposis | Serrated adenomas: | Sessile serrated | Hereditary non-polyposis colorectal cancer (HNPCC) |
| Juvenile polyps | Traditional serrated | |||
| Peutz-Jeghers syndrome | Malignant lesions: | Carcinoma in situ | ||
| MUTYH associated polyposis | ||||
| Lymphoid polyps | Intramucosal CRC | |||
| Invasive CRC | ||||
| Neoplasia Found | Recommanded Interval for Colonoscopy Examination | Comment |
|---|---|---|
| Small rectal hyperplastic polyps | 10 years | Exception are patients with hyperplastic polyposis syndrome, who need more intensive follow up. |
| One or two small (<1 cm) tubular adenomas with only low-grade dysplasia | 5–10 years | The precise timing within this interval should be based on other clinical factors (such as prior colonoscopy findings, family history, and the preferences of the patient and judgment of the physician). |
| 3 to 10 adenomas, or any adenoma ≥ 1 cm, or any adenoma with villous features, or high-grade dysplasia | 3 years | Adenomas must have been completely removed. If the follow up colonoscopy is normal or shows only 1 or 2 small, tubular adenomas with low-grade dysplasia, then the interval for the subsequent examination should be 5 years. |
| More than 10 adenomas at one examination | < 3 years | The interval should be based on the clinician judgement and consider the possibility of an underlying familial syndrome. |
| Sessile adenomas that are removed piecemeal | 2 to 6 months | Once complete removal has been established, subsequent surveillance needs to be individualized based on the endoscopist’s judgment. Completeness of removal should be based on both endoscopic and pathologic assessments. |
| Reference | Origin of Study | Source | Number of Patients | miRNAs | Significant Relevant |
|---|---|---|---|---|---|
| [89] | USA | tissue | 84 adenomas | miR-21 | ↑ expression associated with poor survival |
| [90] | Netherlands | tissue | 25 CRC 30 adenomas | miR-17-92 cluster | ↑ expression across adenoma carcinoma sequence |
| [81] | USA | tissue | 222 CRC 41 adenomas 52 controls | miR-135b miR-31 miR-1 miR-137 miR-9 miR-99a | ability to distinguish adenomas vs. controls |
| [91] | USA | plasma | 20 CRC 9 adenomas 12 controls | miR-532 miR-331 miR-195 miR-17 miR-142 miR-15b miR-532 miR-652 miR-15b miR-21 miR-339 | ability to distinguish adenomas vs. controls and CRC |
| [83] | China | tissue | 227 adenomas 37 controls | miR-194a | predictor for adenoma recurrence |
| [88] | China | tissue | 40 CRC 16 adenomas | miR-31 miR-135b | ↑ expression in adenoma carcinoma sequence |
| stool | 104 CRC 169 adenomas 109 controls | ↑ expression in stool miR-135 level across the adenoma carcinoma sequence | |||
| [82] | Japan | tissue | 870 CRC 637 adenomas | miR-31 | ↑ miR-31 expression was associated with CIMP status |
| [92] | USA | tissue | 113 adenomas | miR-320a | ↑ expression in adenoma carcinoma sequence |
| miR-145 miR-192 | ↓ expression across adenoma carcinoma sequence | ||||
| [93] | China | serum | 307 CRC 164 adenomas 266 controls | miR19a miR-92a miR-223a | ↑ expression in adenoma carcinoma sequence |
| miR-422 | ↓ expression across adenoma carcinoma sequence | ||||
| [80] | UK | tissue | 13 CRC 55 adenomas 10 controls | miR-135b | ↑ expression across adenoma carcinoma sequence |
| [94] | Netherlands | tissue | 52 CRC 48 adenomas | miR-15a | ↑ expression in adenoma carcinoma sequence |
| [95] | France | tissue | 41 CRC 51 adenomas 34 controls | miR-15b miR-16b miR-21 miR-24 miR-145 miR-150 miR-378 | ↓ expression in adenomas compared to controls |
| [96] | Japan | tissue | 151 CRC 21 adenomas | miR-148a | ↓ expression across adenoma carcinoma sequence |
| [97] | USA | serum | 11 CRC 20 adenomas 10 controls | miR-30b miR-30c miR-146a miR-30d | ↑ expression in adenoma carcinoma sequence |
| [98] | Japan | tissue | 18 CRC with adenomas 3 CRC without adenomas 21 normal mucosa | miR-320 family | ↓ expression in adenomas and early CRC tissue vs. controls |
| [78] | USA | tissue | 109 adenomas | miR-145 miR-143 miR-107a miR-194 miR-26a miR-663b miR-1268 miR-320a miR-1275 | ability to distinguish high risk adenomas from low risk adenomas |
| [86] | Japan | serum (+exosomes) | 26 adenomas 47 controls | miR-21 miR-29a miR-92a miR-135b | ability to distinguish adenomas vs. controls discriminate patients with ↑ risk adenoma |
| [85] | Hungary | tissueplasma | 20 CRC 20 adenomas 20 controls | miR-31 miR-10b miR-183 miR-196a | expression of miRNAs in plasma correlated with matched tissue expression level ability to distinguish adenomas vs. controls and CRC |
| [99] | China | serum | 20 CRC 20 adenomas 20 controls | miR-4463 miR-5704 miR-371b miR-1247 miR-1293 miR-548a miR-107 miR-139 | ability to distinguish CRC vs. adenomas vs. controls |
| [100] | USA | serum | 34 CRC 33 adenomas 35 controls | Ratios of: let-7b/miR-367 miR-130a/miR-409 miR-148/miR-27 miR-148/miR-409 miR-21-miR367 | ability to distinguish adenoma vs. controls |
| miR-17/miR-135b miR-92a/miR-135b miR-451a/miR-491 | ability to distinguish CRC vs. adenomas | ||||
| [84] | Colombia | tissue serum | 45 CRC 25 adenomas 45 controls | miR-141 miR-200c | ↑ expression in adenomas compared to CRC and controls |
| [79] | USA | tissue | 26 adenomas 30 controls | miR-31 miR-135b miR-378a | predictors of serrated neoplasia |
| [101] | Ireland | plasma | 16 CRC 24 adenomas 8 controls | miR-34 miR-150 | ability to distinguish CRC vs. adenomas |
| [76] | China | tissue | 6 CRC 6 adenomas 6 controls | miR-135b miR-18a miR-29b | ↑ expression in adenoma carcinoma sequence |
| miR-1 miR-338 miR-218 | ↓ expression level across adenoma carcinoma sequence |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siskova, A.; Cervena, K.; Kral, J.; Hucl, T.; Vodicka, P.; Vymetalkova, V. Colorectal Adenomas—Genetics and Searching for New Molecular Screening Biomarkers. Int. J. Mol. Sci. 2020, 21, 3260. https://doi.org/10.3390/ijms21093260
Siskova A, Cervena K, Kral J, Hucl T, Vodicka P, Vymetalkova V. Colorectal Adenomas—Genetics and Searching for New Molecular Screening Biomarkers. International Journal of Molecular Sciences. 2020; 21(9):3260. https://doi.org/10.3390/ijms21093260
Chicago/Turabian StyleSiskova, Anna, Klara Cervena, Jan Kral, Tomas Hucl, Pavel Vodicka, and Veronika Vymetalkova. 2020. "Colorectal Adenomas—Genetics and Searching for New Molecular Screening Biomarkers" International Journal of Molecular Sciences 21, no. 9: 3260. https://doi.org/10.3390/ijms21093260