The Differential Effect of Carbon Dots on Gene Expression and DNA Methylation of Human Embryonic Lung Fibroblasts as a Function of Surface Charge and Dose

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
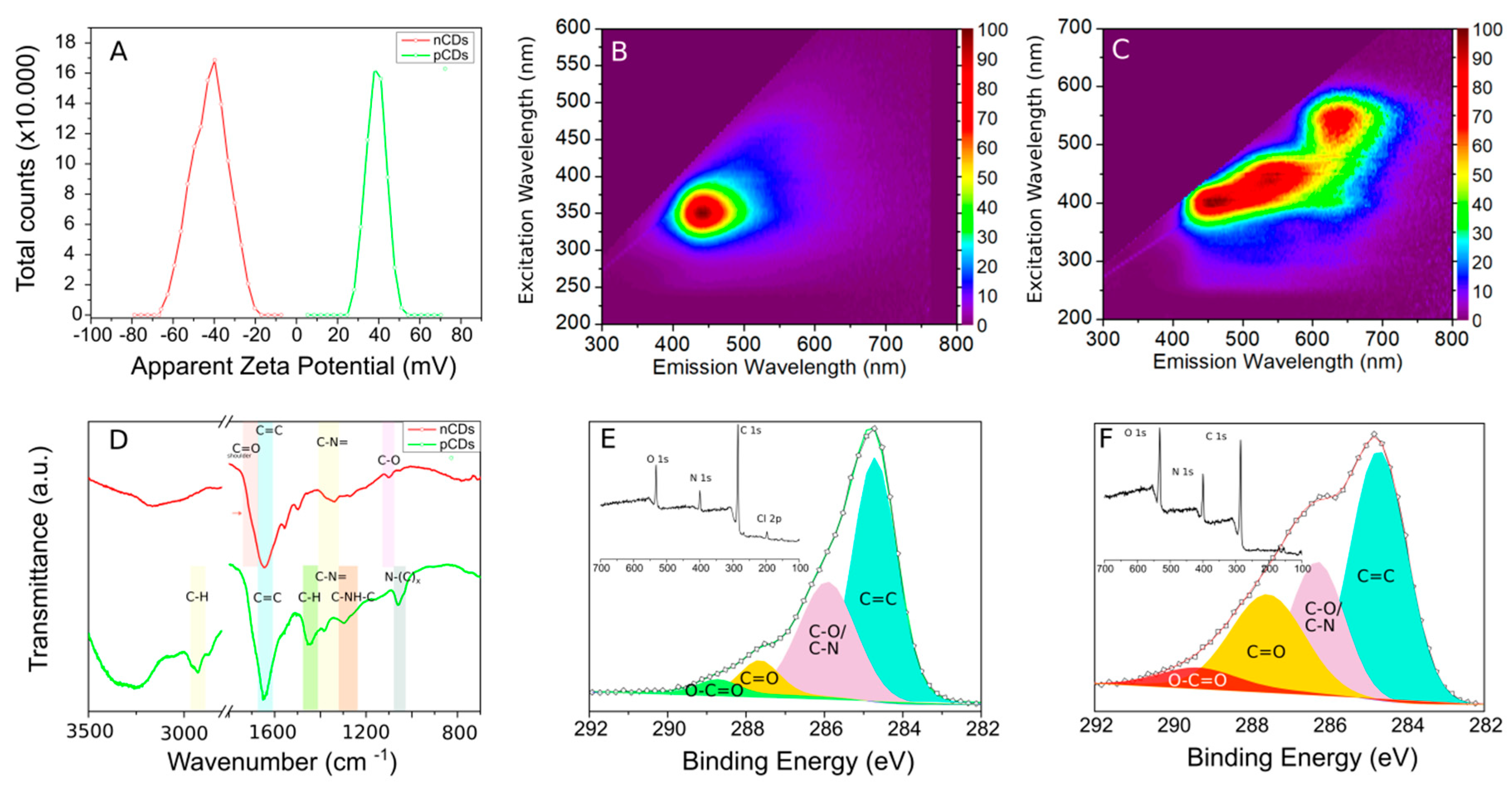
2.1. Physico-Chemical Properties of CDs
2.2. Cell Viability after pCD and nCD Exposure
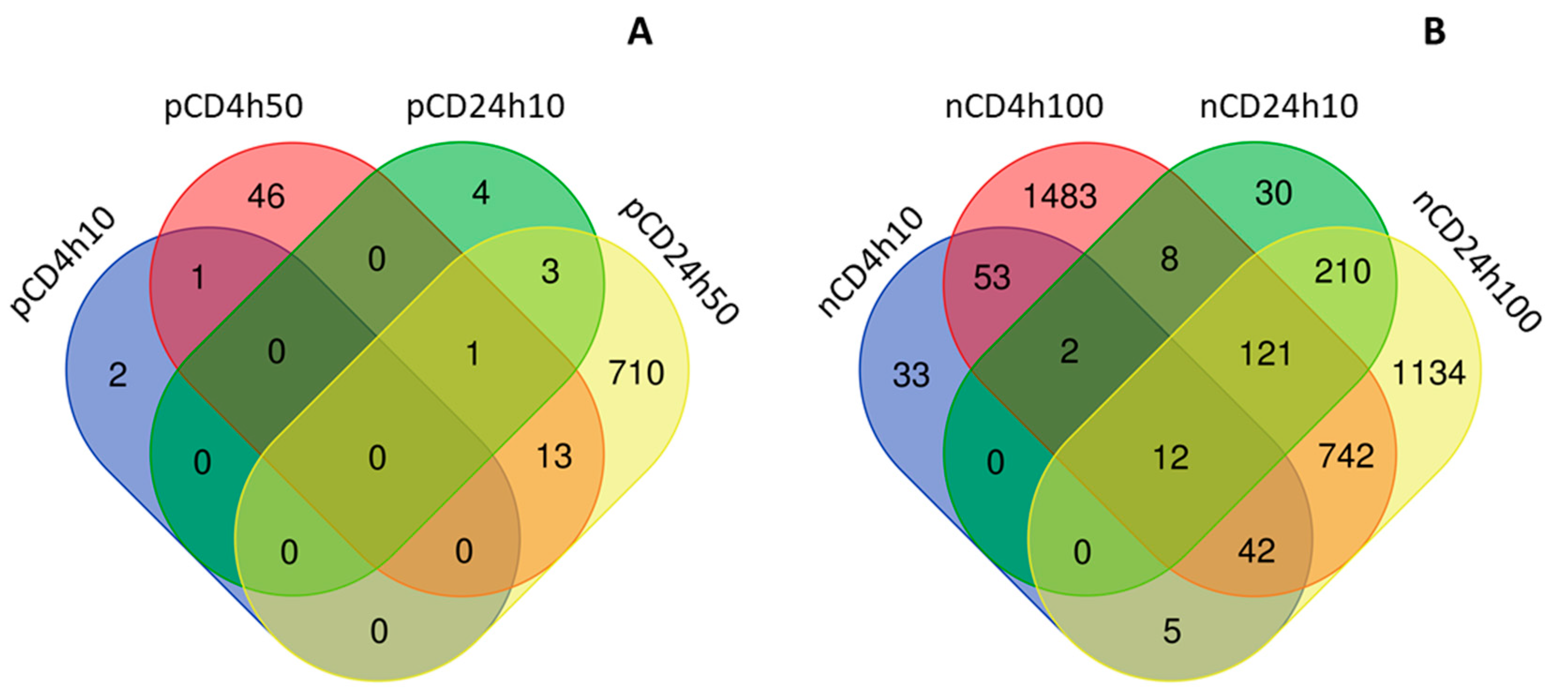
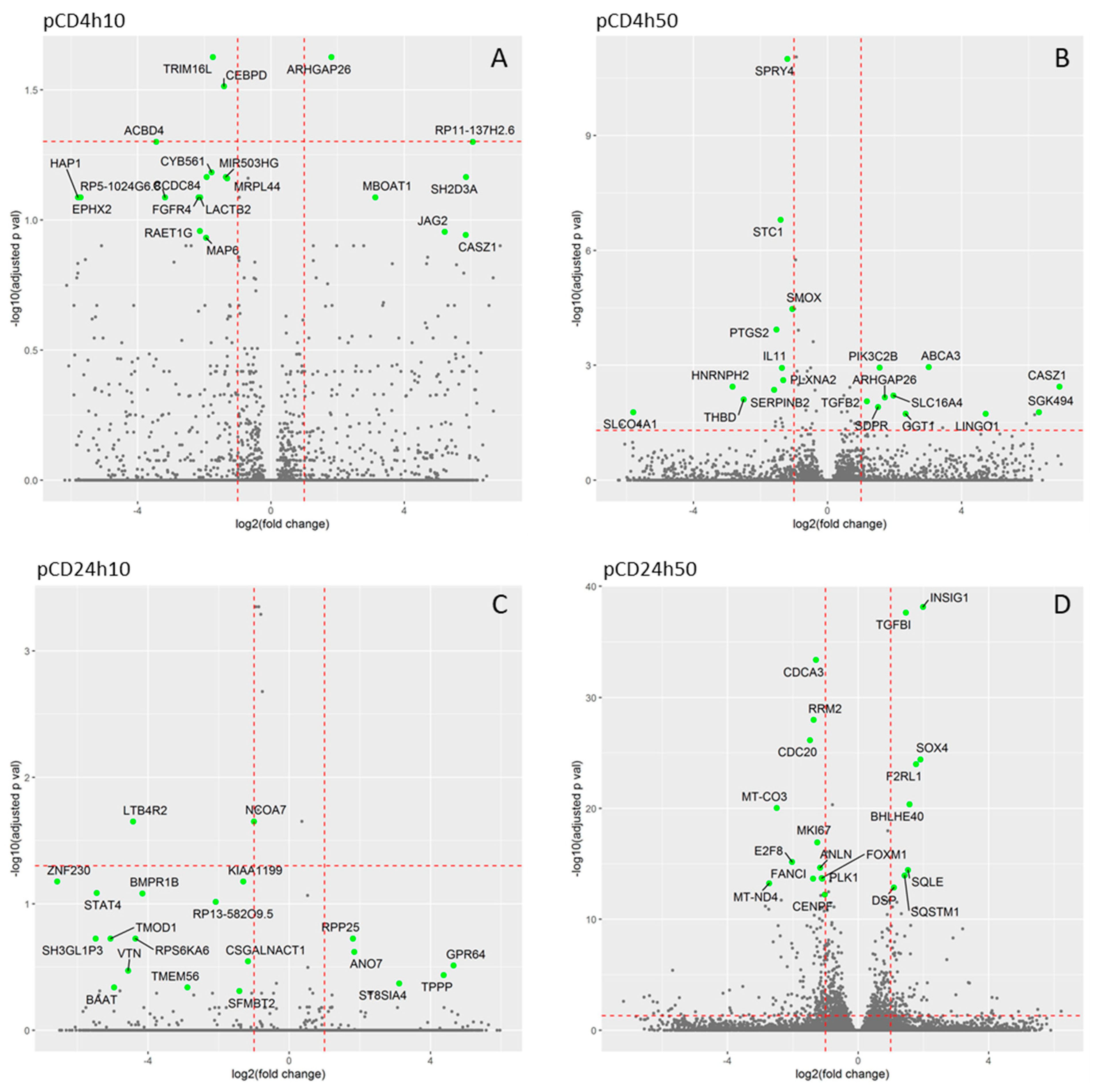
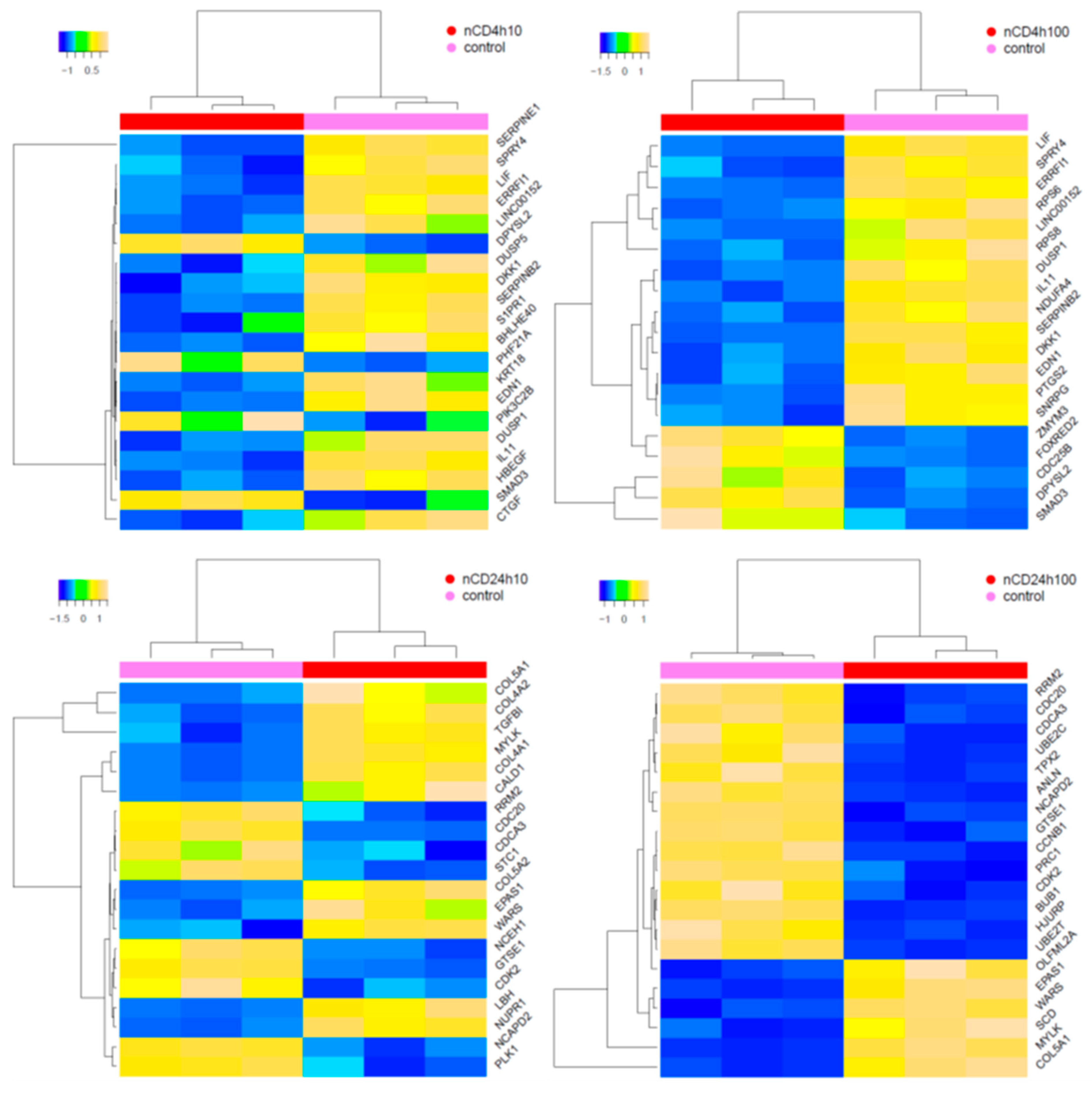
2.3. Expression of mRNAs Induced by pCD and nCD NPs Treatment
2.4. The Expression of miRNAs Affected by Treatment with pCD and nCD NPs
2.5. DNA Methylation Changes after Treatment with pCD and nCD NPs
3. Discussion
4. Materials and Methods
4.1. Carbon Dots and Their Characterization
4.2. Cell Culture Treament and Viability Assay
4.3. DNA, RNA, and miRNA Extraction
4.4. Whole Genome Transcriptome Analysis by Next Generation Sequencing
4.5. miRNA Expression Analysis
4.6. Gene Specific DNA Methylation Analysis
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, X.; Ray, R.; Gu, Y.; Ploehn, H.; Gearheart, L.; Raker, K.; Scrivens, W. Electrophoretic analysis and purification of fluorescent single-walled carbon nanotube fragments. J. Am. Chem. Soc. 2004, 126, 12736–12737. [Google Scholar] [CrossRef] [PubMed]
- Duran, N.; Simoes, M.B.; de Moraes, A.C.M.; Favaro, W.J.; Seabra, A.B. Nanobiotechnology of Carbon Dots: A Review. J. Biomed. Nanotechnol. 2016, 12, 1323–1347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zhao, Y.; Liu, Y.; Chang, X.; Chen, C.; Zhao, Y. Cellular Uptake, Intracellular Trafficking, and Cytotoxicity of Nanomaterials. Small 2011, 7, 1322–1337. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Yang, K.; Ma, Z.; Wan, J.; Zhang, Y.; Kang, Z.; Liu, Z. In Vivo NIR Fluorescence Imaging, Biodistribution, and Toxicology of Photoluminescent Carbon Dots Produced from Carbon Nanotubes and Graphite. Small 2012, 8, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-T.; Wang, X.; Wang, H.; Lu, F.; Luo, P.G.; Cao, L.; Meziani, M.J.; Liu, J.-H.; Liu, Y.; Chen, M.; et al. Carbon Dots as Nontoxic and High-Performance Fluorescence Imaging Agents. J. Phys. Chem. 2009, 113, 18110–18114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Anilkumar, P.; Cao, L.; Liu, J.-H.; Luo, P.G.; Tackett, K.N., II; Sahu, S.; Wang, P.; Wang, X.; Sun, Y.-P. Carbon dots of different composition and surface functionalization: Cytotoxicity issues relevant to fluorescence cell imaging. Exp. Biol. Med. 2011, 236, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Havrdova, M.; Hola, K.; Skopalik, J.; Tomankova, K.; Martin, P.A.; Cepe, K.; Polakova, K.; Tucek, J.; Bourlinos, A.B.; Zboril, R. Toxicity of carbon dots—Effect of surface functionalization on the cell viability, reactive oxygen species generation and cell cycle. Carbon 2016, 99, 238–248. [Google Scholar] [CrossRef]
- Hsu, P.-C.; Chen, P.-C.; Ou, C.-M.; Chang, H.-Y.; Chang, H.-T. Extremely high inhibition activity of photoluminescent carbon nanodots toward cancer cells. J. Mater. Chem. B 2013, 1, 1774–1781. [Google Scholar] [CrossRef]
- Li, C.-L.; Ou, C.-M.; Huang, C.-C.; Wu, W.-C.; Chen, Y.-P.; Lin, T.-E.; Ho, L.-C.; Wang, C.-W.; Shih, C.-C.; Zhou, H.-C.; et al. Carbon dots prepared from ginger exhibiting efficient inhibition of human hepatocellular carcinoma cells. J. Mater. Chem. B 2014, 2, 4564–4571. [Google Scholar] [CrossRef]
- Das, B.; Pal, P.; Dadhich, P.; Dutta, J.; Dhara, S. In Vivo Cell Tracking, Reactive Oxygen Species Scavenging, and Antioxidative Gene Down Regulation by Long-Term Exposure of Biomass-Derived Carbon Dots. ACS Biomater-Sci. Eng. 2019, 5, 346–356. [Google Scholar] [CrossRef]
- Li, M.; Gu, M.-M.; Tian, X.; Xiao, B.-B.; Lu, S.; Zhu, W.; Yu, L.; Shang, Z.-F. Hydroxylated-Graphene Quantum Dots Induce DNA Damage and Disrupt Microtubule Structure in Human Esophageal Epithelial Cells. Toxicol. Sci. 2018, 164, 339–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Panwar, V.; Chopra, V.; Thomas, J.; Kaushik, S.; Ghosh, D. Interaction of Carbon Dots with Endothelial Cells: Implications for Biomedical Applications. ACS Appl. Nano Mater. 2019, 2, 5483–5491. [Google Scholar] [CrossRef]
- Chen, J.; Liu, B.; Yang, Z.; Qu, J.; Xun, H.; Dou, R.; Gao, X.; Wang, L. Phenotypic, transcriptional, physiological and metabolic responses to carbon nanodot exposure in Arabidopsis thaliana (L.). Environ. Sci. Nano 2018, 5, 2672–2685. [Google Scholar] [CrossRef]
- Maiti, S.; Das, K.; Das, P.K. Label-free fluorimetric detection of histone using quaternized carbon dot-DNA nanobiohybrid. Chem. Commun. 2013, 49, 8851–8853. [Google Scholar] [CrossRef]
- Datta, K.K.R.; Kozak, O.; Ranc, V.; Havrdova, M.; Bourlinos, A.B.; Safarova, K.; Hola, K.; Tomankova, K.; Zoppellaro, G.; Otyepka, M.; et al. Quaternized carbon dot-modified graphene oxide for selective cell labelling—Controlled nucleus and cytoplasm imaging. Chem. Commun. 2014, 50, 10782–10785. [Google Scholar] [CrossRef]
- Malina, T.; Polakova, K.; Skopalik, J.; Milotova, V.; Hola, K.; Havrdova, M.; Tomankova, K.B.; Cmiel, V.; Sefc, L.; Zboril, R. Carbon dots for in vivo fluorescence imaging of adipose tissue-derived mesenchymal stromal cells. Carbon 2019, 152, 434–443. [Google Scholar] [CrossRef]
- Bourlinos, A.B.; Zboril, R.; Petr, J.; Bakandritsos, A.; Krysmann, M.; Giannelis, E.P. Luminescent Surface Quaternized Carbon Dots. Chem. Mater. 2012, 24, 6–8. [Google Scholar] [CrossRef]
- Hola, K.; Sudolska, M.; Kalytchuk, S.; Nachtigallova, D.; Rogach, A.L.; Otyepka, M.; Zboril, R. Graphitic Nitrogen Triggers Red Fluorescence in Carbon Dots. ACS Nano 2017, 11, 12402–12410. [Google Scholar] [CrossRef]
- Lin-Vien, D.; Colthup, N.B.; Fateley, W.G.; Grasselli, J.G. The Handbook of Infrared and Raman Characteristic Frequencies of Organic Molecules; Elsevier: Amsterdam, The Netherlands, 1991; p. 503. [Google Scholar]
- Alley, D.F.; Langley-Turnbaugh, S.; Gordon, N.R.; Wise, J.P.; Van Eps, G.; Jalbert, A. The effect of PM10 on human lung fibroblasts. Toxicol. Ind. Health 2009, 25, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, P.; Hu, C.; Wang, K.; Chang, Q.; Liu, L.; Han, Z.; Shao, Y.; Zhai, Y.; Zuo, Z.; et al. Protein corona of airborne nanoscale PM2.5 induces aberrant proliferation of human lung fibroblasts based on a 3D organotypic culture. Sci. Rep. 2018, 8, 1939. [Google Scholar] [CrossRef]
- Durantie, E.; Barosova, H.; Drasler, B.; Rodriguez-Lorenzo, L.; Urban, D.A.; Vanhecke, D.; Septiadi, D.; Hirschi-Ackermann, L.; Petri-Fink, A.; Rothen-Rutishauser, B. Carbon nanodots: Opportunities and limitations to study their biodistribution at the human lung epithelial tissue barrier. Biointerphases 2018, 13, 06D404. [Google Scholar] [CrossRef] [PubMed]
- Tennis, M.A.; Van Scoyk, M.M.; Freeman, S.V.; Vandervest, K.M.; Nemenoff, R.A.; Winn, R.A. Sprouty-4 Inhibits Transformed Cell Growth, Migration and Invasion, and Epithelial-Mesenchymal Transition, and Is Regulated by Wnt7A through PPAR in Non-Small Cell Lung Cancer. Mol. Cancer Res. 2010, 8, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beers, M.F.; Mulugeta, S. The biology of the ABCA3 lipid transporter in lung health and disease. Cell Tissue Res. 2017, 367, 481–493. [Google Scholar] [CrossRef]
- Irigoyen, M.; Pajares, M.J.; Agorreta, J.; Ponz-Sarvisé, M.; Salvo, E.; Lozano, M.D.; Pío, R.; Gil-Bazo, I.; Rouzaut, A. TGFBI expression is associated with a better response to chemotherapy in NSCLC. Mol. Cancer 2010, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Ballard, P.L.; Lee, J.W.; Fang, X.; Chapin, C.; Allen, L.; Segal, M.R.; Fischer, H.; Illek, B.; Gonzales, L.W.; Kolla, V.; et al. Regulated gene expression in cultured type II cells of adult human lung. Am. J. Physiol-Lung. C 2010, 299, L36–L50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, J.-H.; Chen, Z.-H.; Shao, C.-L.; Pei, W.-W.; Zhang, J.; Zhang, S.-Y.; Jiao, Y.; Tong, J. Analysis of the miRNA-mRNA networks in malignant transformation BEAS-2B cells induced by alpha-particles. J. Toxicol. Environ. Health A 2016, 79, 427–435. [Google Scholar] [CrossRef]
- Du, Y.-Z.; Gu, X.-H.; Cheng, S.-F.; Li, L.; Liu, H.; Hu, L.-P.; Gao, F. The oncogenetic role of stanniocalcin 1 in lung adenocarcinoma: A promising serum candidate biomarker for tracking lung adenocarcinoma progression. Tumour Biol. 2016, 37, 5633–5644. [Google Scholar] [CrossRef]
- Adams, M.N.; Burgess, J.T.; He, Y.; Gately, K.; Snell, C.; Zhang, S.-D.; Hooper, J.D.; Richard, D.J.; O’Byrne, K.J. Expression of CDCA3 Is a Prognostic Biomarker and Potential Therapeutic Target in Non-Small Cell Lung Cancer. J. Thorac. Oncol. 2017, 12, 1071–1084. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.-Y.; Liu, X.-X.; Li, H.; Li, R.; Liu, X.; Qu, Y.-Q. Elevated mRNA Levels of AURKA, CDC20 and TPX2 are associated with poor prognosis of smoking related lung adenocarcinoma using bioinformatics analysis. Int. J. Med. Sci. 2018, 15, 1676–1685. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wu, L.-Y.; Yang, W.-X. Nanoparticles induce apoptosis via mediating diverse cellular pathways. Nanomedicine 2018, 13, 2939–2955. [Google Scholar] [CrossRef]
- Li, C.; Park, S.; Zhang, X.; Eisenberg, L.M.; Zhao, H.; Darzynkiewicz, Z.; Xu, D. Nuclear Gene 33/Mig6 regulates the DNA damage response through an ATM serine/threonine kinase-dependent mechanism. J. Biol. Chem. 2017, 292, 16746–16759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, D. Epithelium-fibroblast interactions in response to airway inflammation. Immunol. Cell Biol. 2001, 79, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.-Y.; Wang, W.-J.; Mei, J.-H.; Wang, C.-L. Mitogen-Activated Protein Kinase Signal Transduction in Solid Tumors. Asian Pac. J. Cancer Prev. 2014, 15, 8539–8548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eymin, B.; Gazzeri, S. Role of cell cycle regulators in lung carcinogenesis. Cell Adh. Migr. 2010, 4, 114–123. [Google Scholar] [CrossRef]
- Jin, H.; Kang, G.-Y.; Jeon, S.; Kim, J.-M.; Park, Y.N.; Cho, J.; Lee, Y.-S. Identification of molecular signatures involved in radiation-induced lung fibrosis. J. Mol. Med. 2019, 97, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhang, H.; Du, Y.; Gu, X. Prognostic significance of BRCA1, ERCC1, RRM1, and RRM2 in patients with advanced non-small cell lung cancer receiving chemotherapy. Tumour Biol. 2014, 35, 12679–12688. [Google Scholar] [CrossRef]
- Mok, M.T.; Zhou, J.; Tang, W.; Zeng, X.; Oliver, A.W.; Ward, S.E.; Cheng, A.S. CCRK is a novel signalling hub exploitable in cancer immunotherapy. Pharmacol. Therapeut. 2018, 186, 138–151. [Google Scholar] [CrossRef]
- Ko, J.-W.; Shin, N.-R.; Park, J.-W.; Park, S.-H.; Lee, I.-C.; Kim, J.-S.; Kim, J.-C.; Ahn, K.-S.; Shin, I.-S. Copper oxide nanoparticles induce collagen deposition via TGF-beta 1/Smad3 signaling in human airway epithelial cells. Nanotoxicology 2018, 12, 239–250. [Google Scholar] [CrossRef]
- Hu, J.; Lin, W.; Lin, B.; Wu, K.; Fan, H.; Yu, Y. Persistent DNA methylation changes in zebrafish following graphene quantum dots exposure in surface chemistry-dependent manner. Ecotoxicol. Environ. Saf. 2019, 169, 370–375. [Google Scholar] [CrossRef]
- Rossner, P., Jr.; Vrbova, K.; Rossnerova, A.; Zavodna, T.; Milcova, A.; Klema, J.; Vecera, Z.; Mikuska, P.; Coufalik, P.; Capka, L.; et al. Gene Expression and Epigenetic Changes in Mice Following Inhalation of Copper(II) Oxide Nanoparticles. Nanomaterials 2020, 10, 550. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Bardes, E.E.; Aronow, B.J.; Jegga, A.G. ToppGene Suite for gene list enrichment analysis and candidate gene prioritization. Nucleic Acids Res. 2009, 37, W305–W311. [Google Scholar] [CrossRef] [PubMed]
- Vila-Casadesus, M.; Gironella, M.; Jose Lozano, J. MiRComb: An R Package to Analyse miRNA-mRNA Interactions. Examples across Five Digestive Cancers. PLoS ONE 2016, 11, e0151127. [Google Scholar] [CrossRef] [PubMed]
- Maksimovic, J.; Phipson, B.; Oshlack, A. A cross-package Bioconductor workflow for analysing methylation array data. F1000Research 2016, 5, 1281. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
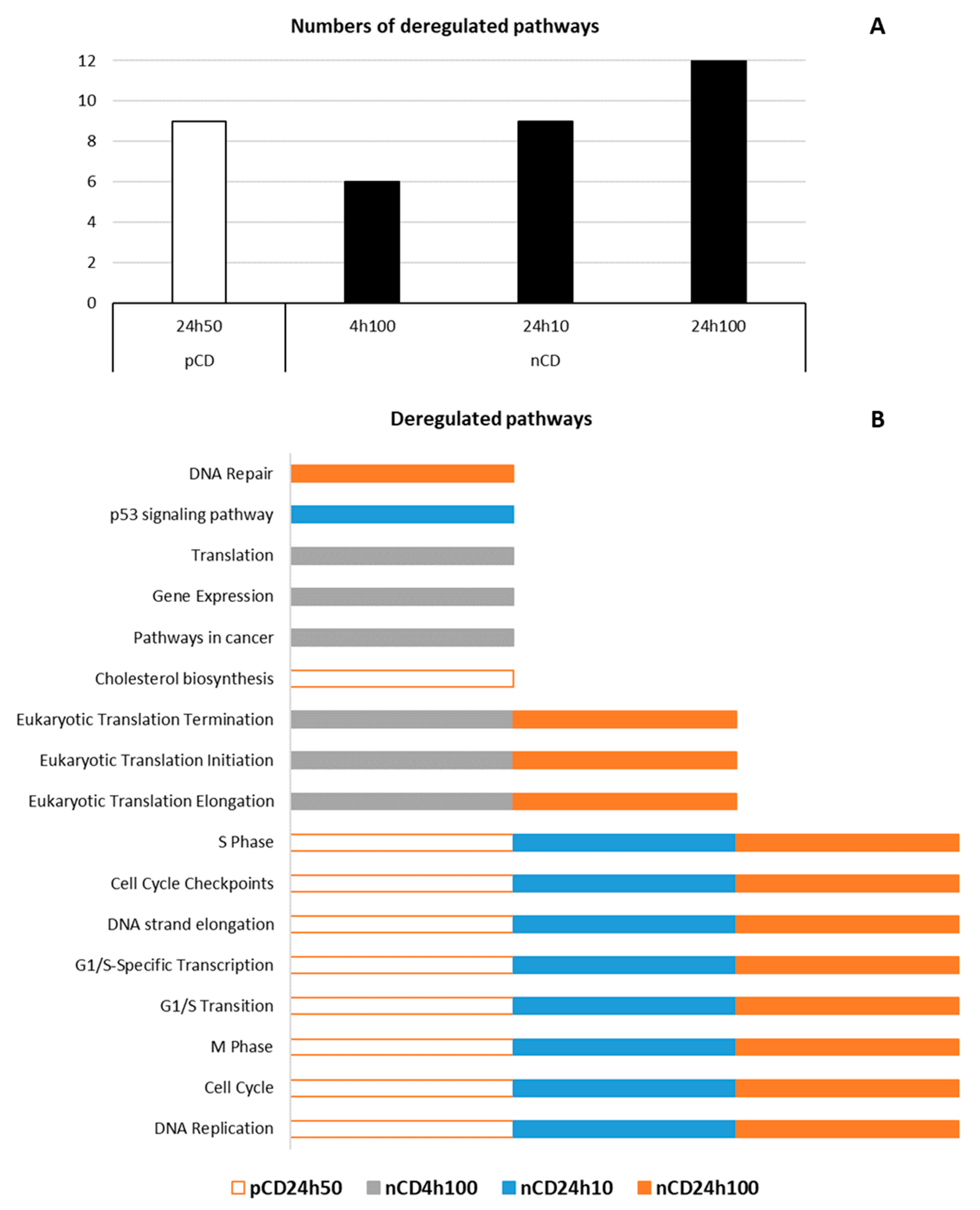
| ID | Source | Deregulated Pathway | q-Value | Deregulated Genes (N) | Genes in Pathway (N) |
|---|---|---|---|---|---|
| pCD 24 h 50 µg/mL | |||||
| 83039 | KEGG | DNA Replication | <0.001 | 14 | 36 |
| 1269741 | REACTOME | Cell Cycle | <0.001 | 106 | 624 |
| 1269810 | REACTOME | M Phase | <0.001 | 48 | 311 |
| 1269768 | REACTOME | G1/S Transition | <0.001 | 25 | 121 |
| 1269772 | REACTOME | G1/S-Specific Transcription | <0.001 | 12 | 18 |
| 1269784 | REACTOME | DNA strand elongation | <0.001 | 17 | 32 |
| 1269742 | REACTOME | Cell Cycle Checkpoints | <0.001 | 29 | 204 |
| 1269777 | REACTOME | S Phase | <0.001 | 29 | 132 |
| 1270037 | REACTOME | Cholesterol biosynthesis | <0.001 | 9 | 24 |
| ID | Source | Deregulated Pathway | q-Value | Deregulated Genes (N) | Genes in Pathway (N) |
|---|---|---|---|---|---|
| nCD 4 h 100 µg/mL | |||||
| 83105 | KEGG | Pathways in cancer | <0.001 | 82 | 395 |
| 1268690 | REACTOME | Eukaryotic Translation Elongation | <0.001 | 49 | 98 |
| 1268679 | REACTOME | Eukaryotic Translation Initiation | <0.001 | 55 | 127 |
| 1268692 | REACTOME | Eukaryotic Translation Termination | <0.001 | 47 | 97 |
| 1269649 | REACTOME | Gene Expression | <0.001 | 345 | 1844 |
| 1268678 | REACTOME | Translation | <0.001 | 68 | 165 |
| nCD 24 h 10 µg/mL | |||||
| 83039 | KEGG | DNA Replication | <0.001 | 8 | 36 |
| 83055 | KEGG | p53 signaling pathway | <0.001 | 9 | 69 |
| 1269741 | REACTOME | Cell Cycle | <0.001 | 63 | 624 |
| 1269810 | REACTOME | M Phase | <0.001 | 26 | 311 |
| 1269768 | REACTOME | G1/S Transition | <0.001 | 18 | 121 |
| 1269772 | REACTOME | G1/S-Specific Transcription | <0.001 | 5 | 18 |
| 1269784 | REACTOME | DNA strand elongation | <0.001 | 11 | 32 |
| 1269742 | REACTOME | Cell Cycle Checkpoints | <0.001 | 16 | 204 |
| 1269777 | REACTOME | S Phase | <0.001 | 21 | 132 |
| nCD 24 h 100 µg/mL | |||||
| 83039 | KEGG | DNA Replication | <0.001 | 24 | 36 |
| 1269741 | REACTOME | Cell Cycle | <0.001 | 216 | 624 |
| 1269810 | REACTOME | M Phase | <0.001 | 104 | 311 |
| 1269768 | REACTOME | G1/S Transition | <0.001 | 49 | 121 |
| 1269772 | REACTOME | G1/S-Specific Transcription | <0.001 | 15 | 18 |
| 1269784 | REACTOME | DNA strand elongation | <0.001 | 25 | 32 |
| 1269742 | REACTOME | Cell Cycle Checkpoints | <0.001 | 66 | 204 |
| 1269777 | REACTOME | S Phase | <0.001 | 52 | 132 |
| 1268690 | REACTOME | Eukaryotic Translation Elongation | <0.001 | 38 | 98 |
| 1268679 | REACTOME | Eukaryotic Translation Initiation | <0.001 | 43 | 127 |
| 1268692 | REACTOME | Eukaryotic Translation Termination | <0.001 | 36 | 97 |
| 1270350 | REACTOME | DNA Repair | <0.001 | 80 | 319 |
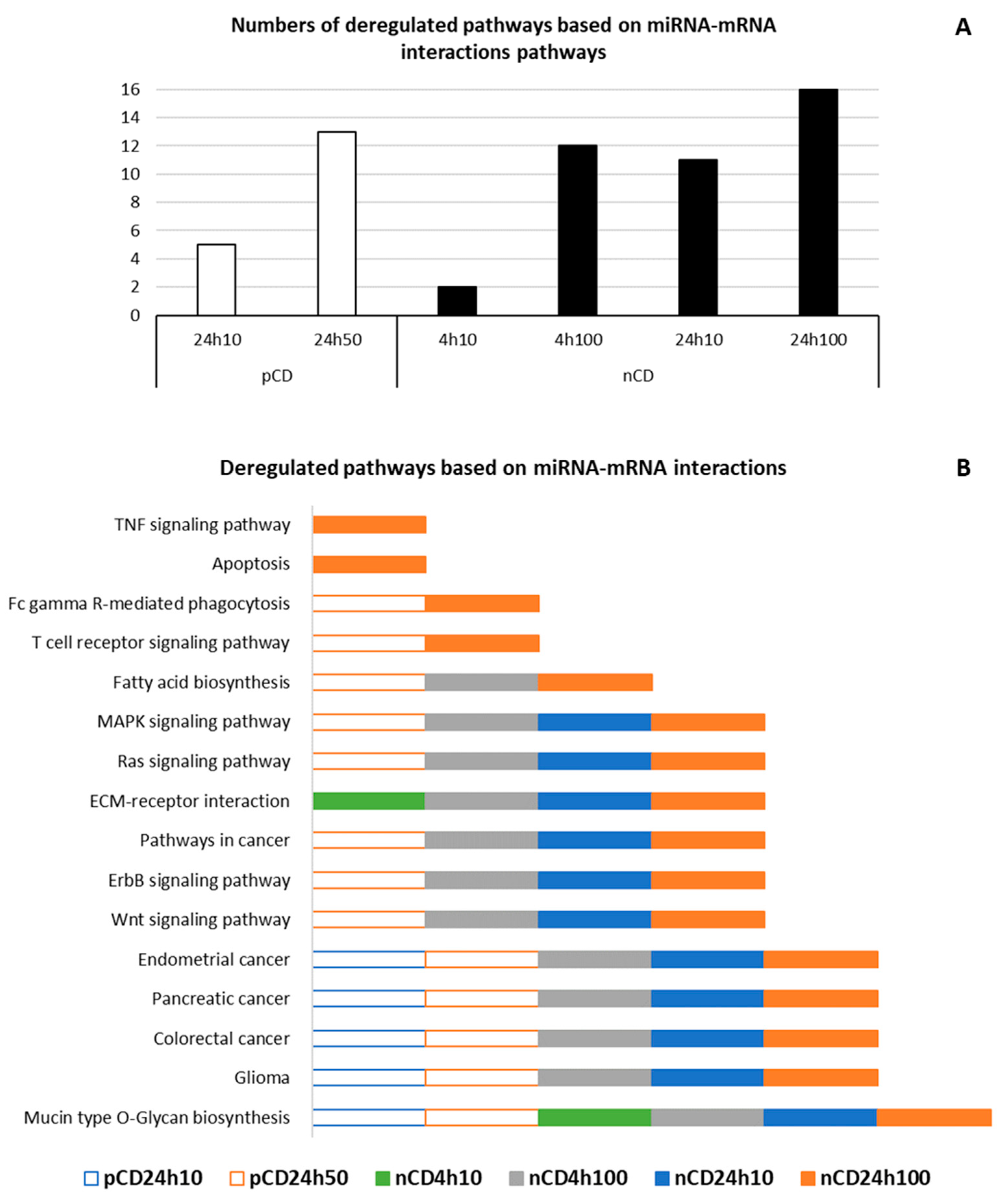
| ID | Deregulated Pathway | q-Value | miRNA (N) | Genes (N) |
|---|---|---|---|---|
| pCD 24 h 10 µg/mL | ||||
| hsa00512 | Mucin type O-Glycan biosynthesis | <0.001 | 5 | 11 |
| hsa05214 | Glioma | 0.049 | 5 | 15 |
| hsa05210 | Colorectal cancer | 0.003 | 5 | 20 |
| hsa05212 | Pancreatic cancer | 0.013 | 5 | 19 |
| hsa05213 | Endometrial cancer | 0.028 | 6 | 17 |
| pCD 24 h 50 µg/mL | ||||
| hsa00512 | Mucin type O-Glycan biosynthesis | <0.001 | 24 | 26 |
| hsa05214 | Glioma | 0.005 | 41 | 45 |
| hsa05210 | Colorectal cancer | <0.001 | 41 | 48 |
| hsa05212 | Pancreatic cancer | <0.001 | 41 | 52 |
| hsa05213 | Endometrial cancer | <0.001 | 41 | 42 |
| hsa04310 | Wnt signaling pathway | <0.001 | 44 | 99 |
| hsa04012 | ErbB signaling pathway | <0.001 | 41 | 66 |
| hsa05200 | Pathways in cancer | <0.001 | 47 | 266 |
| hsa04014 | Ras signaling pathway | <0.001 | 44 | 156 |
| hsa04010 | MAPK signaling pathway | 0.040 | 44 | 160 |
| hsa00061 | Fatty acid biosynthesis | <0.001 | 15 | 8 |
| hsa04660 | T cell receptor signaling pathway | 0.024 | 41 | 71 |
| hsa04666 | Fc gamma R-mediated phagocytosis | 0.007 | 41 | 64 |
| ID | Deregulated Pathway | q-Value | miRNA (N) | Genes (N) |
|---|---|---|---|---|
| nCD 4 h 10 µg/mL | ||||
| hsa00512 | Mucin type O-Glycan biosynthesis | 0.009 | 9 | 7 |
| hsa04512 | ECM-receptor interaction | <0.001 | 10 | 17 |
| nCD 4 h 100 µg/mL | ||||
| hsa00512 | Mucin type O-Glycan biosynthesis | <0.001 | 46 | 23 |
| hsa05214 | Glioma | <0.001 | 78 | 52 |
| hsa05210 | Colorectal cancer | 0.013 | 73 | 50 |
| hsa05212 | Pancreatic cancer | 0.003 | 79 | 53 |
| hsa05213 | Endometrial cancer | 0.022 | 72 | 42 |
| hsa04310 | Wnt signaling pathway | 0.003 | 89 | 108 |
| hsa04012 | ErbB signaling pathway | <0.001 | 86 | 73 |
| hsa05200 | Pathways in cancer | <0.001 | 94 | 303 |
| hsa04014 | Ras signaling pathway | <0.001 | 91 | 175 |
| hsa04010 | MAPK signaling pathway | 0.018 | 93 | 184 |
| hsa04512 | ECM-receptor interaction | <0.001 | 21 | 27 |
| hsa00061 | Fatty acid biosynthesis | <0.001 | 34 | 10 |
| nCD 24 h 10 µg/mL | ||||
| hsa00512 | Mucin type O-Glycan biosynthesis | <0.001 | 26 | 20 |
| hsa05214 | Glioma | <0.001 | 46 | 49 |
| hsa05210 | Colorectal cancer | 0.001 | 40 | 46 |
| hsa05212 | Pancreatic cancer | <0.001 | 45 | 50 |
| hsa05213 | Endometrial cancer | 0.006 | 40 | 40 |
| hsa04310 | Wnt signaling pathway | 0.007 | 52 | 93 |
| hsa04012 | ErbB signaling pathway | <0.001 | 51 | 66 |
| hsa05200 | Pathways in cancer | <0.001 | 59 | 260 |
| hsa04014 | Ras signaling pathway | 0.002 | 55 | 145 |
| hsa04010 | MAPK signaling pathway | 0.004 | 53 | 164 |
| hsa04512 | ECM-receptor interaction | <0.001 | 49 | 55 |
| nCD 24 h 100 µg/mL | ||||
| hsa00512 | Mucin type O-Glycan biosynthesis | <0.001 | 51 | 23 |
| hsa05214 | Glioma | <0.001 | 69 | 55 |
| hsa05210 | Colorectal cancer | <0.001 | 71 | 57 |
| hsa05212 | Pancreatic cancer | <0.001 | 72 | 58 |
| hsa05213 | Endometrial cancer | <0.001 | 68 | 45 |
| hsa04310 | Wnt signaling pathway | <0.001 | 84 | 108 |
| hsa04012 | ErbB signaling pathway | <0.001 | 80 | 80 |
| hsa05200 | Pathways in cancer | <0.001 | 91 | 288 |
| hsa04014 | Ras signaling pathway | <0.001 | 87 | 171 |
| hsa04010 | MAPK signaling pathway | 0.002 | 87 | 191 |
| hsa04512 | ECM-receptor interaction | 0.002 | 72 | 61 |
| hsa00061 | Fatty acid biosynthesis | <0.001 | 34 | 9 |
| hsa04660 | T cell receptor signaling pathway | <0.001 | 72 | 85 |
| hsa04666 | Fc gamma R-mediated phagocytosis | 0.024 | 77 | 66 |
| hsa04210 | Apoptosis | 0.036 | 70 | 66 |
| hsa04668 | TNF signaling pathway | 0.016 | 76 | 84 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sima, M.; Vrbova, K.; Zavodna, T.; Honkova, K.; Chvojkova, I.; Ambroz, A.; Klema, J.; Rossnerova, A.; Polakova, K.; Malina, T.; et al. The Differential Effect of Carbon Dots on Gene Expression and DNA Methylation of Human Embryonic Lung Fibroblasts as a Function of Surface Charge and Dose. Int. J. Mol. Sci. 2020, 21, 4763. https://doi.org/10.3390/ijms21134763
Sima M, Vrbova K, Zavodna T, Honkova K, Chvojkova I, Ambroz A, Klema J, Rossnerova A, Polakova K, Malina T, et al. The Differential Effect of Carbon Dots on Gene Expression and DNA Methylation of Human Embryonic Lung Fibroblasts as a Function of Surface Charge and Dose. International Journal of Molecular Sciences. 2020; 21(13):4763. https://doi.org/10.3390/ijms21134763
Chicago/Turabian StyleSima, Michal, Kristyna Vrbova, Tana Zavodna, Katerina Honkova, Irena Chvojkova, Antonin Ambroz, Jiri Klema, Andrea Rossnerova, Katerina Polakova, Tomas Malina, and et al. 2020. "The Differential Effect of Carbon Dots on Gene Expression and DNA Methylation of Human Embryonic Lung Fibroblasts as a Function of Surface Charge and Dose" International Journal of Molecular Sciences 21, no. 13: 4763. https://doi.org/10.3390/ijms21134763