CAIX-Mediated Control of LIN28/let-7 Axis Contributes to Metabolic Adaptation of Breast Cancer Cells to Hypoxia

 , ,
, , 
Abstract
:1. Introduction
2. Results
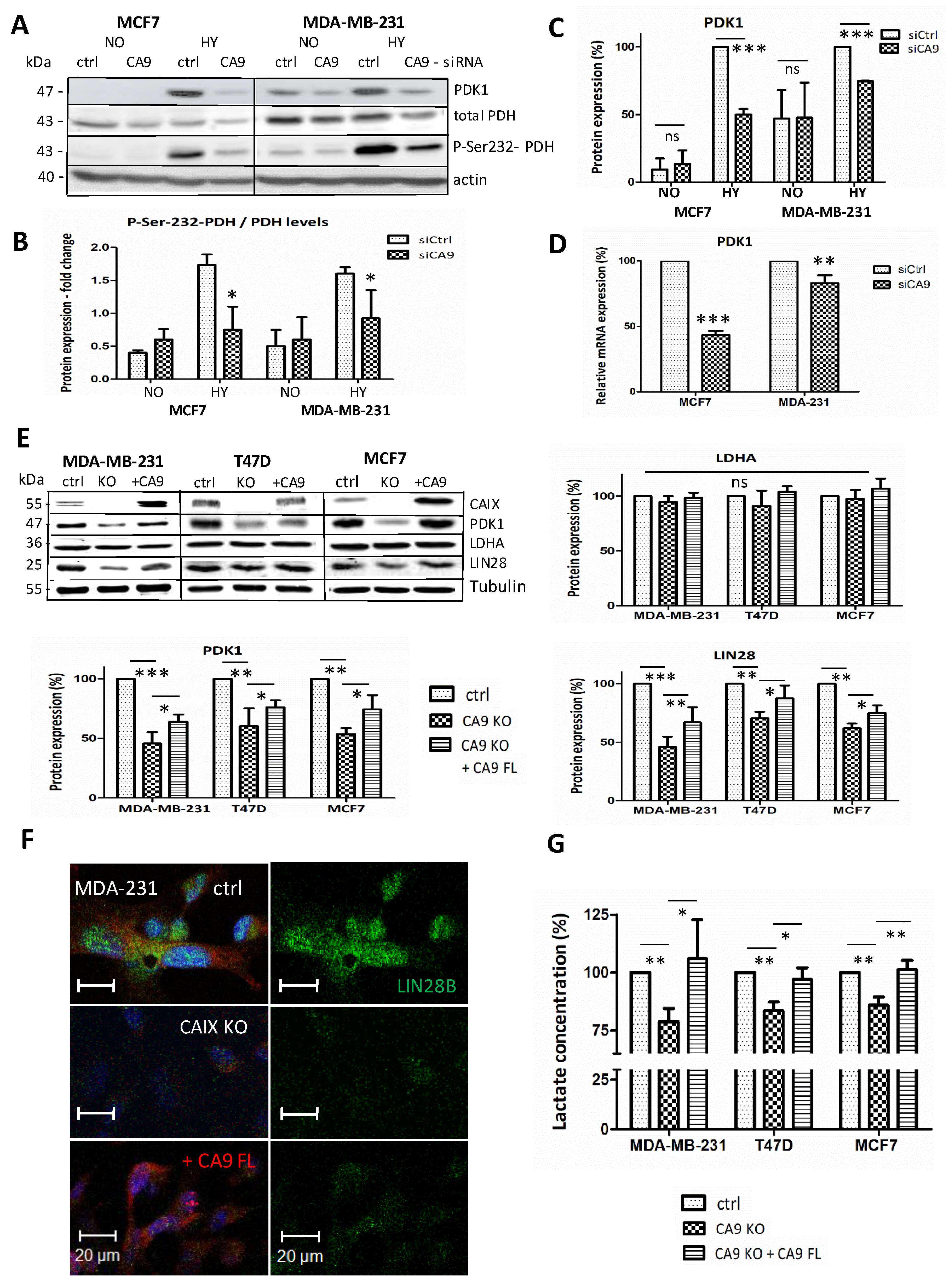
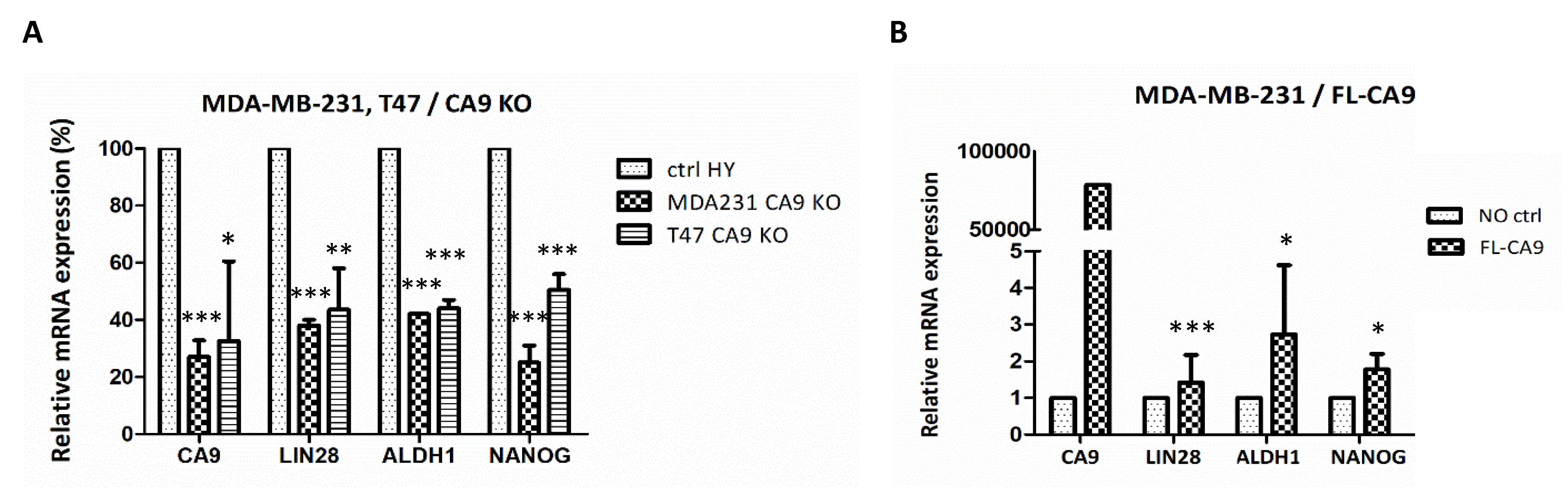
2.1. Suppression of CAIX Affects the Let-7/LIN28 Axis
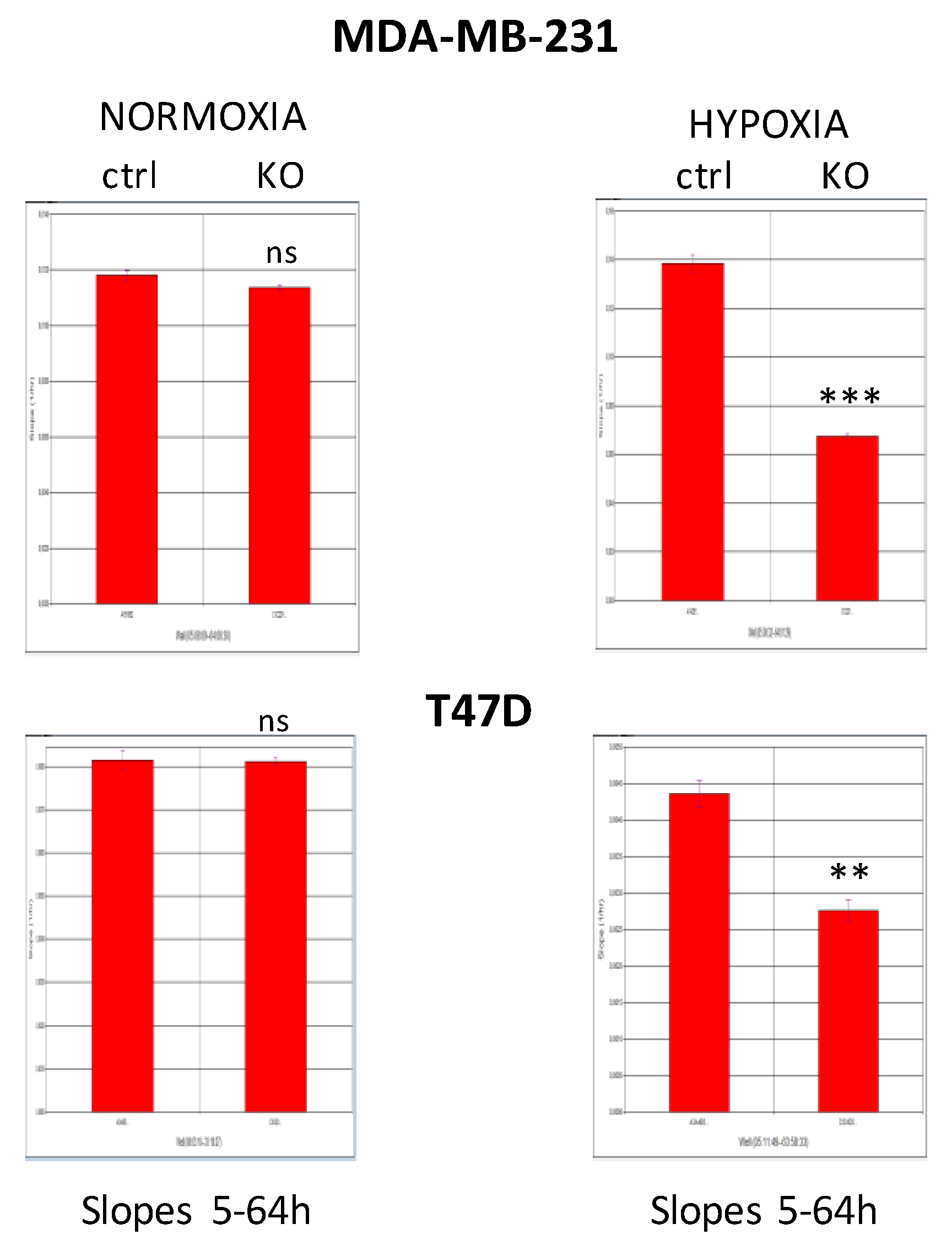
2.2. CAIX Suppression Reduces Glycolysis by Targeting PDK1
2.3. CAIX Depletion Reduces NF-κB Expression and Transactivation
2.4. Extracellular Acidosis and Inhibition of CAIX Enzymatic Activity Affects NF-κB, LIN28B and PDK1 Expression
2.5. CAIX Knockout Decreases Cell Proliferation
2.6. CAIX Expression Influences LIN28 Together with Stem Cell Markers
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Microarray Analysis
4.3. Transient Silencing
4.4. Generation of Knockout Cell Line with CRISPR/Cas9 System
4.5. Immunoblotting
4.6. Real-Time Quantitative PCR (qPCR)
4.7. Lactate Measurement
4.8. xCELLigence Real-Time Cell Assay (RTCA)
4.9. Reporter Assay for NF-κB
4.10. Immunofluorescence
4.11. Statistical Analyzes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CAIX | carbonic anhydrase IX protein |
| CA9 | corresponding gene |
| PDK1 | pyruvate dehydrogenase kinase |
| PDH | pyruvate dehydrogenase |
| LDHA | lactate dehydrogenase A |
| ALDH1 | aldehyde dehydrogenase isoform 1 |
| CSCs | cancer stem cells |
| pHi | intracellular pH |
| pHe | extracellular pH |
| HIF-1 | hypoxia inducible factor 1 |
| NO | normoxia |
| HY | hypoxia |
| NF-κB | nuclear factor-κB |
| mTORC1 | mammalian target of rapamycin complex 1 |
| HSFA | 4-Homosulfanilamide |
References
- Pastorek, J.; Pastorekova, S. Hypoxia-induced carbonic anhydrase IX as a target for cancer therapy: From biology to clinical use. Semin. Cancer Biol. 2015, 31, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Svastova, E.; Witarski, W.; Csaderova, L.; Kosik, I.; Skvarkova, L.; Hulikova, A.; Zatovicova, M.; Barathova, M.; Kopacek, J.; Pastorek, J.; et al. Carbonic anhydrase IX interacts with bicarbonate transporters in lamellipodia and increases cell migration via its catalytic domain. J. Biol. Chem. 2012, 287, 3392–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debreova, M.; Csaderova, L.; Burikova, M.; Lukacikova, L.; Kajanova, I.; Sedlakova, O.; Kery, M.; Kopacek, J.; Zatovicova, M.; Bizik, J.; et al. CAIX regulates invadopodia formation through both a pH-dependent mechanism and interplay with actin regulatory proteins. Int. J. Mol. Sci. 2019, 20, 2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatenby, R.A.; Smallbone, K.; Maini, P.K.; Rose, F.; Averill, J.; Nagle, R.B.; Worrall, L.; Gillies, R.J. Cellular adaptations to hypoxia and acidosis during somatic evolution of breast cancer. Br. J. Cancer 2007, 97, 646–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, B.A.; Chimenti, M.; Jacobson, M.P.; Barber, D.L. Dysregulated pH: A perfect storm for cancer progression. Nat. Rev. Cancer 2011, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Kulshreshtha, R.; Davuluri, R.V.; Calin, G.A.; Ivan, M. A microRNA component of the hypoxic response. Cell Death Differ. 2008, 15, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Hebert, C.; Norris, K.; Scheper, M.A.; Nikitakis, N.; Sauk, J.J. High mobility group A2 is a target for miRNA-98 in head and neck squamous cell carcinoma. Mol. Cancer 2007, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Hua, Z.; Lv, Q.; Ye, W.; Wong, C.K.; Cai, G.; Gu, D.; Ji, Y.; Zhao, C.; Wang, J.; Yang, B.B.; et al. MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS ONE 2006, 1, e116. [Google Scholar] [CrossRef] [Green Version]
- Heo, I.; Joo, C.; Cho, J.; Ha, M.; Han, J.; Kim, V.N. Lin28 mediates the terminal uridylation of let-7 precursor MicroRNA. Mol. Cell 2008, 32, 276–284. [Google Scholar] [CrossRef]
- Ambros, V.; Horvitz, H.R. Heterochronic mutants of the nematode Caenorhabditis elegans. Science 1984, 226, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An epigenetic switch involving NF-kappaB, Lin28, Let-7 MicroRNA, and IL6 links inflammation to cell transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, S.R.; Daley, G.Q.; Gregory, R.I. Selective blockade of microRNA processing by Lin28. Science 2008, 320, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; He, Y.; Zhu, Y.; Chen, M.; Weng, M.; Yang, C.; Zhang, Y.; Ning, N.; Zhao, R.; Yang, W.; et al. Comparison of the expression and function of Lin28A and Lin28B in colon cancer. Oncotarget 2016, 7, 79605–79616. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, C.; Sun, L.; Huang, D.; Li, T.; He, X.; Wu, G.; Yang, Z.; Zhong, X.; Song, L.; et al. Lin28/let-7 axis regulates aerobic glycolysis and cancer progression via PDK1. Nat. Commun. 2014, 5, 5212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Shyh-Chang, N.; Segre, A.V.; Shinoda, G.; Shah, S.P.; Einhorn, W.S.; Takeuchi, A.; Engreitz, J.M.; Hagan, J.P.; Kharas, M.G.; et al. The Lin28/let-7 axis regulates glucose metabolism. Cell 2011, 147, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.H.; Zhu, H. Lin28 and let-7 in cell metabolism and cancer. Transl. Pediatr. 2015, 4, 4–11. [Google Scholar]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/let-7 pathway in cancer. Front. Genet. 2017, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Kaelin, W.G.; Thompson, C.B. Q and A: Cancer: Clues from cell metabolism. Nature 2010, 465, 562–564. [Google Scholar] [CrossRef]
- Velpula, K.K.; Bhasin, A.; Asuthkar, S.; Tsung, A.J. Combined targeting of PDK1 and EGFR triggers regression of glioblastoma by reversing the warburg effect. Cancer Res. 2013, 73, 7277–7289. [Google Scholar] [CrossRef] [Green Version]
- Shyh-Chang, N.; Daley, G.Q. Lin28: Primal regulator of growth and metabolism in stem cells. Cell Stem Cell 2013, 12, 395–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Li, N.; Liang, S.; Huang, Q.; Coukos, G.; Zhang, L. Identification of microRNAs regulating reprogramming factor LIN28 in embryonic stem cells and cancer cells. J. Biol. Chem. 2010, 285, 41961–41971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chafe, S.C.; Lou, Y.; Sceneay, J.; Vallejo, M.; Hamilton, M.J.; McDonald, P.C.; Bennewith, K.L.; Moller, A.; Dedhar, S. Carbonic anhydrase IX promotes myeloid-derived suppressor cell mobilization and establishment of a metastatic niche by stimulating G-CSF production. Cancer Res. 2015, 75, 996–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peppicelli, S.; Bianchini, F.; Contena, C.; Tombaccini, D.; Calorini, L. Acidic pH via NF-κB favours VEGF-C expression in human melanoma cells. Clin. Exp. Metastasis 2013, 30, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Fidler, I.J. Acidic pH-induced elevation in interleukin 8 expression by human ovarian carcinoma cells. Cancer Res. 2000, 60, 4610–4616. [Google Scholar]
- Xiong, Q.; Shi, Q.; Le, X.; Wang, B.; Xie, K. Regulation of interleukin-8 expression by nitric oxide in human pancreatic adenocarcinoma. J. Interf. Cytokine Res. 2001, 21, 529–537. [Google Scholar] [CrossRef]
- Fukumura, D.; Xu, L.; Chen, Y.; Gohongi, T.; Seed, B.; Jain, R.K. Hypoxia and acidosis independently up-regulate vascular endothelial growth factor transcription in brain tumors in vivo. Cancer Res. 2001, 61, 6020–6024. [Google Scholar]
- Zhou, J.; Ng, S.B.; Chng, W.J. LIN28/LIN28B: An emerging oncogenic driver in cancer stem cells. Int. J. Biochem. Cell Biol. 2013, 45, 973–978. [Google Scholar] [CrossRef]
- Yang, X.; Lin, X.; Zhong, X.; Kaur, S.; Li, N.; Liang, S.; Lassus, H.; Wang, L.; Katsaros, D.; Montone, K.; et al. Double-negative feedback loop between reprogramming factor LIN28 and microRNA let-7 regulates aldehyde dehydrogenase 1-positive cancer stem cells. Cancer Res. 2010, 70, 9463–9472. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Yao, H.; Zhu, P.; Zhang, X.; Pan, Q.; Gong, C.; Huang, Y.; Hu, X.; Su, F.; Lieberman, J.; et al. let-7 regulates self renewal and tumorigenicity of breast cancer cells. Cell 2007, 131, 1109–1123. [Google Scholar] [CrossRef] [Green Version]
- Reinhart, B.J.; Slack, F.J.; Basson, M.; Pasquinelli, A.E.; Bettinger, J.C.; Rougvie, A.E.; Horvitz, H.R.; Ruvkun, G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 2000, 403, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Thornton, J.E.; Gregory, R.I. How does Lin28 let-7 control development and disease? Trends Cell Biol. 2012, 22, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L. HIF-1: Upstream and downstream of cancer metabolism. Curr. Opin. Genet. Dev. 2010, 20, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.W.; Tchernyshyov, I.; Semenza, G.L.; Dang, C. V HIF-1-mediated expression of pyruvate dehydrogenase kinase: A metabolic switch required for cellular adaptation to hypoxia. Cell Metab. 2006, 3, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab. 2006, 3, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golias, T.; Papandreou, I.; Sun, R.; Kumar, B.; Brown, N.V.; Swanson, B.J.; Pai, R.; Jaitin, D.; Le, Q.T.; Teknos, T.N.; et al. Hypoxic repression of pyruvate dehydrogenase activity is necessary for metabolic reprogramming and growth of model tumours. Sci. Rep. 2016, 6, 31146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlowski, A.; De Giusti, V.C.; Morgan, P.E.; Aiello, E.A.; Alvarez, B. V Binding of carbonic anhydrase IX to extracellular loop 4 of the NBCe1 Na+/HCO3- cotransporter enhances NBCe1-mediated HCO3- influx in the rat heart. Am. J. Physiol. Cell Physiol. 2012, 303, C69–C80. [Google Scholar] [CrossRef] [Green Version]
- Swietach, P.; Patiar, S.; Supuran, C.T.; Harris, A.L.; Vaughan-Jones, R.D. The role of carbonic anhydrase 9 in regulating extracellular and intracellular ph in three-dimensional tumor cell growths. J. Biol. Chem. 2009, 284, 20299–20310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svastova, E.; Hulikova, A.; Rafajova, M.; Zat’ovicova, M.; Gibadulinova, A.; Casini, A.; Cecchi, A.; Scozzafava, A.; Supuran, C.T.; Pastorek, J.; et al. Hypoxia activates the capacity of tumor-associated carbonic anhydrase IX to acidify extracellular pH. FEBS Lett. 2004, 577, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.C.; Singh, R.; Pochampally, R.; Watabe, K.; Mo, Y.Y. Acidosis promotes invasiveness of breast cancer cells through ROS-AKT-NF-kappaB pathway. Oncotarget 2014, 5, 12070–12082. [Google Scholar] [CrossRef] [Green Version]
- Balgi, A.D.; Diering, G.H.; Donohue, E.; Lam, K.K.; Fonseca, B.D.; Zimmerman, C.; Numata, M.; Roberge, M. Regulation of mTORC1 signaling by pH. PLoS ONE 2011, 6, e21549. [Google Scholar] [CrossRef]
- Boyd, N.H.; Walker, K.; Fried, J.; Hackney, J.R.; McDonald, P.C.; Benavides, G.A.; Spina, R.; Audia, A.; Scott, S.E.; Libby, C.J.; et al. Addition of carbonic anhydrase 9 inhibitor SLC-0111 to temozolomide treatment delays glioblastoma growth in vivo. JCI Insight 2017, 2, 1–16. [Google Scholar] [CrossRef]
- Hedlund, E.M.E.; McDonald, P.C.; Nemirovsky, O.; Awrey, S.; Jensen, L.D.E.; Dedhar, S. Harnessing induced essentiality: Targeting carbonic anhydrase IX and angiogenesis reduces lung metastasis of triple negative breast cancer xenografts. Cancers 2019, 11, 1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faes, S.; Planche, A.; Uldry, E.; Santoro, T.; Pythoud, C.; Stehle, J.C.; Horlbeck, J.; Letovanec, I.; Riggi, N.; Datta, D.; et al. Targeting carbonic anhydrase IX improves the anti-cancer efficacy of mTOR inhibitors. Oncotarget 2016, 7, 36666–36680. [Google Scholar] [CrossRef]
- McDonald, P.C.; Chafe, S.C.; Brown, W.S.; Saberi, S.; Swayampakula, M.; Venkateswaran, G.; Nemirovsky, O.; Gillespie, J.A.; Karasinska, J.M.; Kalloger, S.E.; et al. Regulation of pH by Carbonic Anhydrase 9 Mediates Survival of Pancreatic Cancer Cells With Activated KRAS in Response to Hypoxia. Gastroenterology 2019, 157, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Hashim, A.A.I.; Abrahams, D.; Xu, L.; Centeno, B.; Sunassee, E.; Abddelgader, R.; Dubois, L.; Lambin, P.; Gatenby, R.A.; Gillies, R.J. Targeting tumor acidity with the LDHA inhibitor (FX11) and CAIX inhibitor (DH348) overcomes resistance to PD-1 blockade and inhibits metastasis in a pancreatic cancer model [abstract]. Proc. Annu. Meet. Am. Assoc. Cancer Res. 2017, 77, 5932. [Google Scholar] [CrossRef]
- Deev, I.E.; Vasilenko, K.P.; Kurmangaliev, E.; Serova, O.V.; Popova, N.V.; Galagan, Y.S.; Burova, E.B.; Zozulya, S.A.; Nikol’skii, N.N.; Petrenko, A.G. Effect of changes in ambient pH on phosphorylation of cellular proteins. Dokl. Biochem. Biophys. 2006, 408, 184–187. [Google Scholar] [CrossRef]
- Tsanov, K.M.; Pearson, D.S.; Wu, Z.; Han, A.; Triboulet, R.; Seligson, M.T.; Powers, J.T.; Osborne, J.K.; Kane, S.; Gygi, S.P.; et al. LIN28 phosphorylation by MAPK/ERK couples signalling to the post-transcriptional control of pluripotency. Nat. Cell Biol. 2017, 19, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Lock, F.E.; McDonald, P.C.; Lou, Y.; Serrano, I.; Chafe, S.C.; Ostlund, C.; Aparicio, S.; Winum, J.Y.; Supuran, C.T.; Dedhar, S. Targeting carbonic anhydrase IX depletes breast cancer stem cells within the hypoxic niche. Oncogene 2012, 32, 5210–5219. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Shin, H.J.; Kim, J.Y.; Byun, H.J.; Lee, J.H.; Sung, Y.K.; Rho, S.B. Dickkopf-1 (DKK-1) interrupts FAK/PI3K/mTOR pathway by interaction of carbonic anhydrase IX (CA9) in tumorigenesis. Cell. Signal. 2012, 24, 1406–1413. [Google Scholar] [CrossRef]
- Parks, S.K.; Chiche, J.; Pouysségur, J. Disrupting proton dynamics and energy metabolism for cancer therapy. Nat. Rev. Cancer 2013, 13, 611–623. [Google Scholar] [CrossRef]
- Ledaki, I.; Mcintyre, A.; Wigfield, S.; Buffa, F.; Baban, D.; Li, J.; Harris, A.L. Carbonic anhydrase IX induction defines a heterogeneous cancer cell response to hypoxia and mediates stem cell-like properties and sensitivity to HDAC inhibition. Oncotargetvol 2015, 6, 19423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pore, N.; Jalla, S.; Liu, Z.; Higgs, B.; Sorio, C.; Scarpa, A.; Hollingsworth, R.; Tice, D.A.; Michelotti, E. In vivo loss of function screening reveals carbonic anhydrase IX as a key modulator of tumor initiating potential in primary pancreatic tumors. Neoplasia 2015, 17, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpern, H.J.; LaManna, J.C.; Harrison, D.K.; Epel, B. Oxygen Transport to Tissue XXXIX; Springer International Publishing AG: Cham, Switzerland, 2017; Chapter 16; pp. 119–124. [Google Scholar] [CrossRef]
- Peng, F.; Wang, J.H.; Fan, W.J.; Meng, Y.T.; Li, M.M.; Li, T.T.; Cui, B.; Wang, H.F.; Zhao, Y.; An, F.; et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene 2018, 37, 1119. [Google Scholar] [CrossRef] [Green Version]
- García-Venzor, A.; Mandujano-Tinoco, E.A.; Lizarraga, F.; Zampedri, C.; Krötzsch, E.; Salgado, R.M.; Dávila-Borja, V.M.; Encarnación-Guevara, S.; Melendez-Zajgla, J.; Maldonado, V. Microenvironment-regulated lncRNA-HAL is able to promote stemness in breast cancer cells. Biochim. Biophys. Acta-Mol. Cell Res. 2019, 1866, 118523. [Google Scholar]
- Radvak, P.; Repic, M.; Svastova, E.; Takacova, M.; Csaderova, L.; Strnad, H.; Pastorek, J.; Pastorekova, S.; Kopacek, J. Suppression of carbonic anhydrase IX leads to aberrant focal adhesion and decreased invasion of tumor cells. Oncol. Rep. 2013, 29, 1147–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kery, M.; Oravcova, N.; Radenkovic, S.; Iuliano, F.; Tomaskova, J.; Golias, T. Pyruvate dehydrogenase kinase 1 and carbonic anhydrase IX targeting in hypoxic tumors. Neoplasma 2018, 66, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibadulinova, A.; Pastorek, M.; Filipcik, P.; Radvak, P.; Csaderova, L.; Vojtesek, B.; Pastorekova, S. Cancer-associated S100P protein binds and inactivates p53, permits therapy-induced senescence and supports chemoresistance. Oncotarget 2016, 7, 22508–22522. [Google Scholar] [CrossRef]
- Pastorekova, S.; Zavadova, Z.; Kostal, M.; Babusikova, O.; Zavada, J. A novel quasi-viral agent, MaTu, is a two-component system. Virology 1992, 187, 620–626. [Google Scholar] [CrossRef]
- Jurkovicova, D.; Lukackova, R.; Magyerkova, M.; Kulcsar, L.; Krivjanska, M.; Krivjansky, V.; Chovanec, M. microRNA expression profiling as supportive diagnostic and therapy prediction tool in chronic myeloid leukemia. Neoplasma 2015, 62, 949–957. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene Name | logFC | p-Value | fdr |
|---|---|---|---|---|
| MIRLET7D | microRNA let-7d | 0.51 | 0.0015 | 0.34 |
| MIRLET7C | microRNA let-7c | 0.5 | 0.017 | 0.36 |
| MIR125B2 | microRNA 125b-2 | 0.43 | 0.04 | 0.4 |
| MIRLET7F1 | microRNA let-7f-1 | 0.43 | 0.045 | 0.4 |
| NFKB1 | nuclear factor kappa B subunit 1 | –0.83 | 0.039 | 0.4 |
| Antigen | Host | Dilution | Company |
|---|---|---|---|
| actin | M | 1:1000 | Cell Signaling 8H10D10 |
| tubulin | Rb | 1:2000 | Abcam 4074 |
| CAIX | M | 1:4 | in-house M75 [60] |
| total PDH E1α | M | 1:6000 | MitoSciences |
| pSer232-E1α | Rb | 1:3000 | EMD Chemicals |
| PDK1 | Rb | 1:1000 | ENZO |
| LIN28A | Rb | 1:1000 | Cell Signaling A177 |
| LIN28B | Rb | 1:500 | Abcam 71415 |
| B-Actin S | CCAACCGCGAGAAGATGACC | B-actin A | GATCTTCATGAGGTAGTCAGT |
| CA9 S | CCGAGCGACGCAGCCTTTGA | CA9 A | GGCTCCAGTCTCGGCTACCT |
| LIN28A F | GAGTGAGAGGCGGCCAAAA | LIN28A R | TGATGATCTAGACCTCCACAGTTGTAG |
| LIN28B F | TGATAAACCGAGAGGGAAGC | LIN28B R | TGTGAATTCCACTGGTTCTCC |
| PDK1 S | ATTGGAAGCATAAATCCAAACTG | PDK1 A | CGGTCACTCATCTTCACAGTC |
| LDHA F | TGGCAGCCTTTTCCTTAGAA | LDHA R | ACTTGCAGTTCGGGCTGTAT |
| ALDH1 F | CGGGAAAAGCAATCTGAAGAGGG | ALDH1 R | GATGCGGCTATACAACACTGGC |
| NANOG S | GCAAATGTCTTCTGCTGAGATGC | NANOG A | AGCTGGGTGGAAGAGAACACAG |
| NF-κB/p65 F | GACCTGAATGCTGTGCGGC | NF-κB/p65 R | ATCTTGAGCTCGGCAGTGTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibadulinova, A.; Bullova, P.; Strnad, H.; Pohlodek, K.; Jurkovicova, D.; Takacova, M.; Pastorekova, S.; Svastova, E. CAIX-Mediated Control of LIN28/let-7 Axis Contributes to Metabolic Adaptation of Breast Cancer Cells to Hypoxia. Int. J. Mol. Sci. 2020, 21, 4299. https://doi.org/10.3390/ijms21124299
Gibadulinova A, Bullova P, Strnad H, Pohlodek K, Jurkovicova D, Takacova M, Pastorekova S, Svastova E. CAIX-Mediated Control of LIN28/let-7 Axis Contributes to Metabolic Adaptation of Breast Cancer Cells to Hypoxia. International Journal of Molecular Sciences. 2020; 21(12):4299. https://doi.org/10.3390/ijms21124299
Chicago/Turabian StyleGibadulinova, Adriana, Petra Bullova, Hynek Strnad, Kamil Pohlodek, Dana Jurkovicova, Martina Takacova, Silvia Pastorekova, and Eliska Svastova. 2020. "CAIX-Mediated Control of LIN28/let-7 Axis Contributes to Metabolic Adaptation of Breast Cancer Cells to Hypoxia" International Journal of Molecular Sciences 21, no. 12: 4299. https://doi.org/10.3390/ijms21124299