We Are What We Eat: A Stoichiometric and Ecometabolomic Study of Caterpillars Feeding on Two Pine Subspecies of Pinus sylvestris

 , , , and
, , , and
Abstract
:1. Introduction
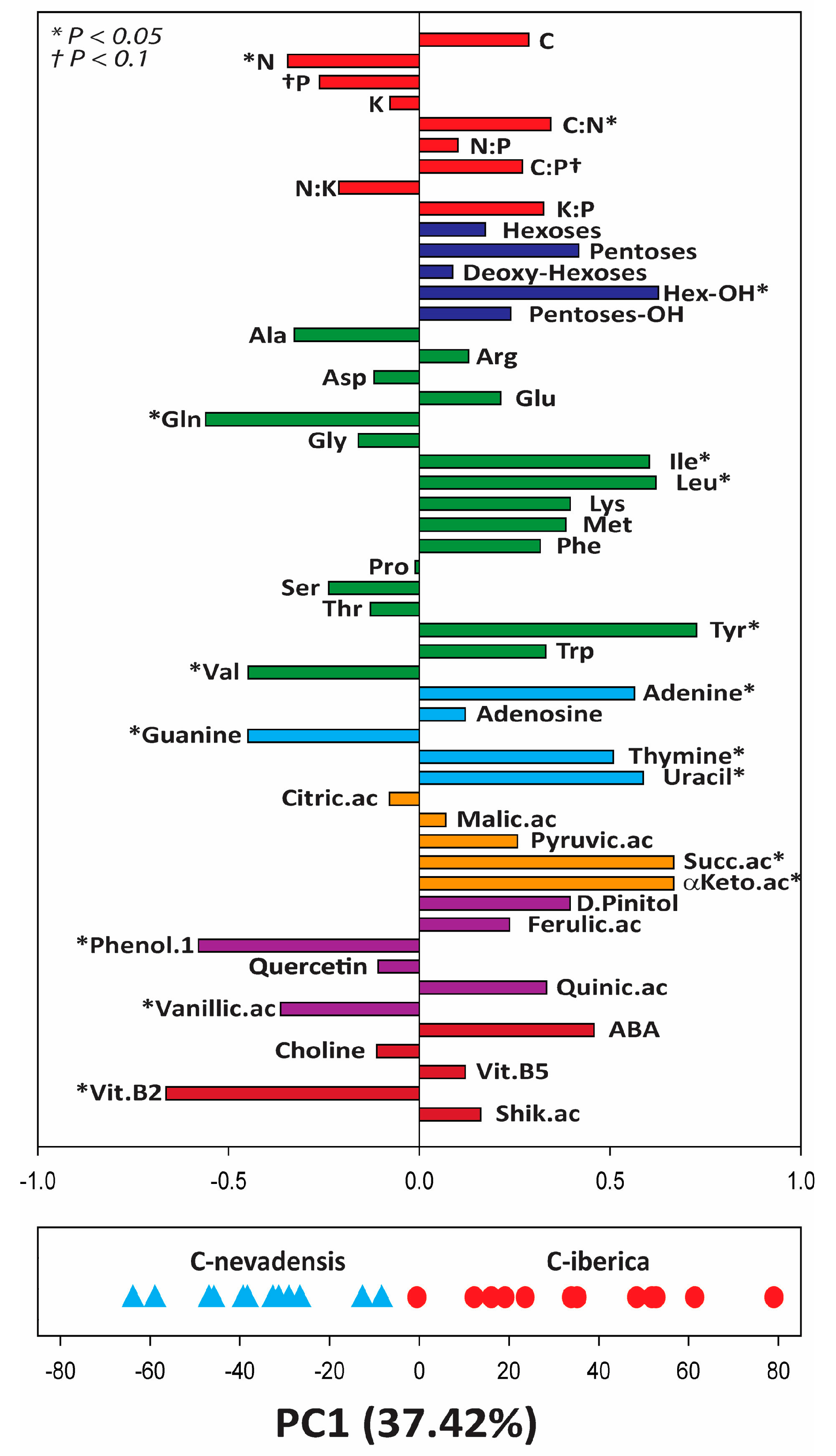
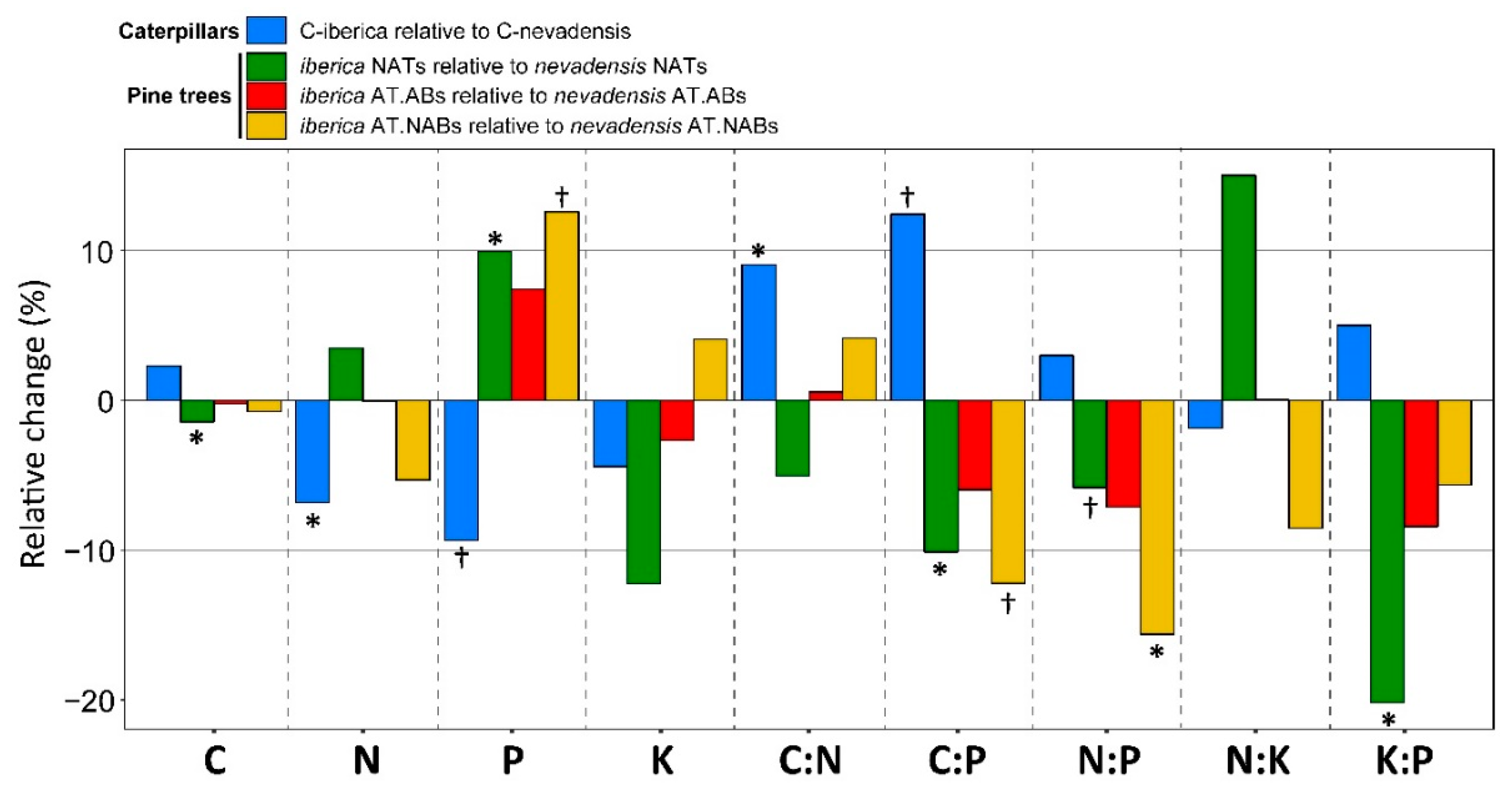
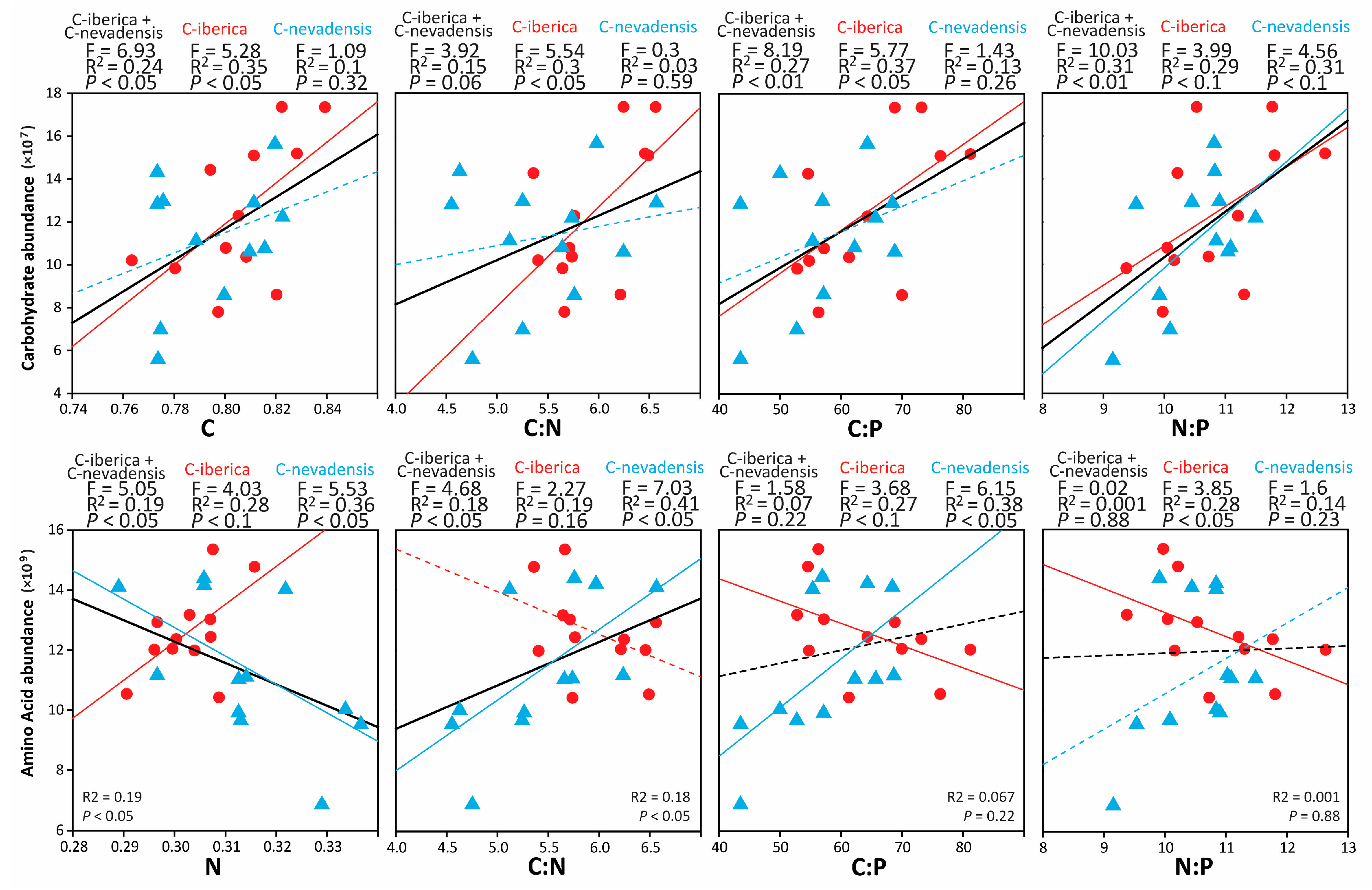
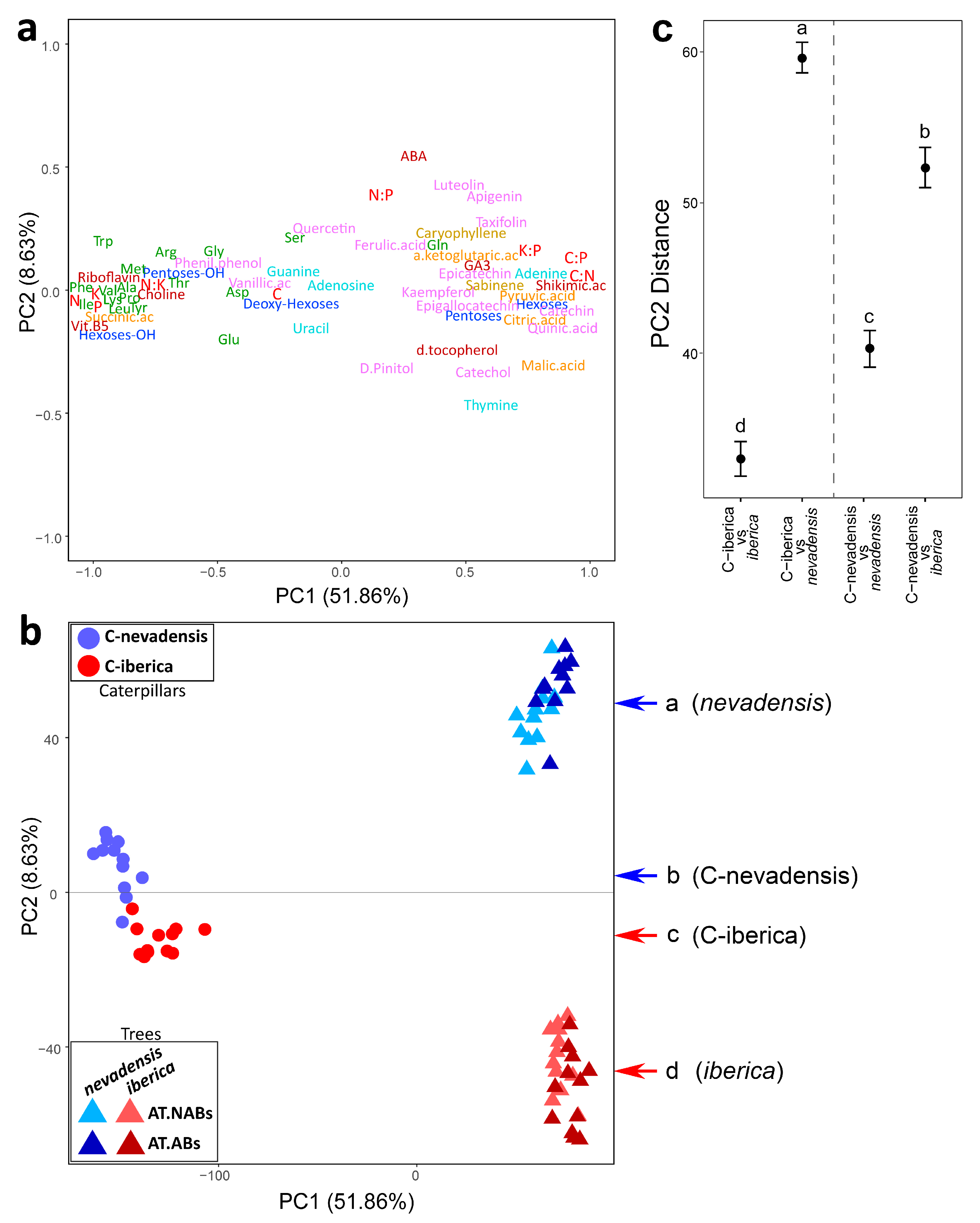
2. Results
3. Discussion
- Do insect herbivores have the ability to accumulate specific plant secondary metabolites to repeal parasitoids? If this ability is found in multiple insect groups, is it consequence of common ancestry or is consequence of multiple independent acquisitions (phenotypic convergence)?
- Why nutrient concentration in insect herbivore tissues is not necessarily correlated with the nutrient abundance of their food source? Are there molecular compounds in the food source that can potentially impact the nutrient transfer across different trophic levels by forcing changes in herbivore physiology?
- How flexible are specialist insects feeding in diverse plant species? Which are the key plant molecular factors constraining the food diversity of specialist herbivores?
4. Material and Methods
4.1. Study Site
4.2. Experiment Design and Sampling of Needles and Caterpillars
4.3. Caterpillar and Foliar Processing for Elemental and Metabolomics Analyses
4.4. Elemental Analysis
4.5. Extraction of Metabolites for Liquid Chromatography-Mass Spectrometry (LC-MS) Analyses
4.6. LC-MS Analysis
4.7. Processing of LC-MS Chromatograms
4.8. Statistical Analyses
- (1)
- All zero values of the dataset were replaced for missing data (NA).
- (2)
- For each variable (metabolite feature, element and stoichiometric ratio), outlier values were determined for each cell individually. A cell corresponds to each group of samples defined by the combination of each factor and level. Therefore, our study is mainly composed by 6 cells: Caterpillars feeding on iberica (C-iberica), caterpillars feeding on nevadensis (C-nevadensis), AT.ABs of iberica, AT.NABs of iberica, AT.ABs of nevadensis and AT.NABs of nevadensis. Since only stoichiometric and elemental data was used from NATs of both pine subspecies, this dataset was filtered separately. Detected outliers were replaced for NA and were defined as:where Q75 represents the third quartile, Q25 represents the first quartile and IQR is the interquartile range (IQR = Q75–Q25) of each variable and cell.
- (3)
- Variables with less than 60% of data within all cells were removed from the dataset.
- (4)
- Variables with signal to noise lower than 15 (determined by the signal from blanks analyzed during the sequence) were removed from the dataset.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Behmer, S.T. Insect Herbivore Nutrient Regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef] [PubMed]
- Bernays, E.A. Evolution of Feeding Behavior in Insect Herbivores. Bioscience 1998, 48, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Pauw, A.; Stofberg, J.; Waterman, R.J. Flies and Flowers in Darwin’s Race. Evolution 2009, 63, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Toxic cardenolides: Chemical ecology and coevolution of specialized plant-herbivore interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Toju, H. Weevils and camellias in a Darwin’s race: Model system for the study of eco-evolutionary interactions between species. Ecol. Res. 2011, 26, 239–251. [Google Scholar] [CrossRef]
- Heimpel, G.E.; Rosenheim, J.A.; Kattari, D. Adult feeding and lifetime reproductive success in the parasitoid Aphytis melinus. Entomol. Exp. Appl. 1997, 83, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Andersen, L.H.; Kristensen, T.N.; Loeschcke, V.; Toft, S.; Mayntz, D. Protein and carbohydrate composition of larval food affects tolerance to thermal stress and desiccation in adult Drosophila melanogaster. J. Insect Physiol. 2010, 56, 336–340. [Google Scholar] [CrossRef]
- Bauerfeind, S.S.; Fischer, K. Testing the plant stress hypothesis: Stressed plants offer better food to an insect herbivore. Entomol. Exp. Appl. 2013, 149, 148–158. [Google Scholar] [CrossRef]
- Kasumovic, M.M. The multidimensional consequences of the juvenile environment: Towards an integrative view of the adult phenotype. Anim. Behav. 2013, 85, 1049–1059. [Google Scholar] [CrossRef]
- Waldbauer, G.P.; Cohen, R.W.; Friedman, S. Self-Selection of an Optimal Nutrient Mix from Defined Diets by Larvae of the Corn Earworm, Heliothis zea (Boddie). Physiol. Zool. 1984, 57, 590–597. [Google Scholar] [CrossRef]
- Simpson, S.J.; Sibly, R.M.; Lee, K.P.; Behmer, S.T.; Raubenheimer, D. Optimal foraging when regulating intake of multiple nutrients. Anim. Behav. 2004, 68, 1299–1311. [Google Scholar] [CrossRef]
- Le Gall, M.; Behmer, S.T. Effects of Protein and Carbohydrate on an Insect Herbivore: The Vista from a Fitness Landscape. Integr. Comp. Biol. 2014, 54, 942–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattson, W.J. Herbivory in Relation to Plant Nitrogen Content. Annu. Rev. Ecol. Syst. 1980, 11, 119–161. [Google Scholar] [CrossRef]
- Sterner, R.; Elser, J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princetion University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Ngai, J.T.; Jefferies, R.L. Nutrient limitation of plant growth and forage quality in Arctic coastal marshes. J. Ecol. 2004, 92, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Chen, Y.; Coleman, T.W.; Jones, M.I.; Flint, M.L.; Seybold, S.J. Foliar nutrients explain goldspotted oak borer, Agrilus auroguttatus, adult feeding preference among four California oak species. Entomol. Exp. Appl. 2013, 149, 57–66. [Google Scholar] [CrossRef]
- Hódar, J.A.; Zamora, R.; Castro, J. Host utilisation by moth and larval survival of pine processionary caterpillar Thaumetopoea pityocampa in relation to food quality in three Pinus species. Ecol. Entomol. 2002, 27, 292–301. [Google Scholar] [CrossRef]
- Metspalu, L.; Kruus, E.; Jögar, K.; Kuusik, A.; Williams, I.H.; Veromann, E.; Luik, A.; Ploomi, A.; Hiiesaar, K.; Kivimägi, I.; Mänd, M. Larval food plants can regulate the cabbage moth, Mamestra brassicae population. Bull. Insectol. 2013, 66, 93–101. [Google Scholar]
- Hódar, J.A.; Castro, J.; Zamora, R. Pine processionary caterpillar Thaumetopoea pityocampa as a new threat for relict Mediterranean Scots pine forests under climatic warming. Biol. Conserv. 2003, 110, 123–129. [Google Scholar] [CrossRef]
- Loaiza, V.; Jonas, J.L.; Joern, A. Grasshoppers (Orthoptera: Acrididae) select vegetation patches in local-scale responses to foliar nitrogen but not phosphorus in native grassland. Insect Sci. 2011, 18, 533–540. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Gargallo-Garriga, A.; Sardans, J.; Oravec, M.; Mateu-Castell, L.; Pérez-Trujillo, M.; Parella, T.; Ogaya, R.; Urban, O.; Peñuelas, J. Drought enhances folivory by shifting foliar metabolomes in Quercus ilex trees. New Phytol. 2014, 202, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Ubach, A.; Sardans, J.; Hódar, J.A.; Garcia-Porta, J.; Guenther, A.; Oravec, M.; Urban, O.; Peñuelas, J. Similar local but different systemic metabolomic responses of closely related pine subspecies to folivory by caterpillars of the processionary moth. Plant Biol. 2016, 18, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Colasurdo, N.; Gélinas, Y.; Despland, E. Larval nutrition affects life history traits in a capital breeding moth. J. Exp. Biol. 2009, 212, 1794–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, J.; Benrey, B.; Thiery, D. Assessing larval food quality for phytophagous insects: Are the facts as simple as they appear? Funct. Ecol. 2006, 20, 92–600. [Google Scholar] [CrossRef]
- Fagan, W.F.; Siemann, E.; Mitter, C.; Denno, R.F.; Huberty, A.F.; Woods, H.A.; Elser, J.J. Nitrogen in insects: Implications for trophic complexity and species diversification. Am. Nat. 2002, 160, 784–802. [Google Scholar] [CrossRef]
- Denno, R.F.; Fagan, W.F. Might nitrogen limitation promote omnivory among carnivorous arthropods? Ecology 2003, 84, 2522–2531. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J. Ecological metabolomics. Chem. Ecol. 2009, 25, 305–309. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Sardans, J.; Peŕez-Trujillo, M.; Estiarte, M.; Penũelas, J. Strong relationship between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 4181–4186. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, C.; George, R.M.; Tattini, M.; Field, K.; Davey, M.P. Metabolomics in plant environmental physiology. J. Exp. Bot. 2013, 64, 4011–4020. [Google Scholar] [CrossRef] [Green Version]
- Sardans, J.; Peñuelas, J.; Rivas-Ubach, A. Ecological metabolomics: Overview of current developments and future challenges. Chemoecology 2011, 21, 191–225. [Google Scholar] [CrossRef]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2008, 5, 3–21. [Google Scholar] [CrossRef]
- Macel, M.; Van Dam, N.M.; Keurentjes, J.J.B. Metabolomics: The chemistry between ecology and genetics. Mol. Ecol. Resour. 2010, 10, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-Ubach, A.; Barbeta, A.; Sardans, J.; Guenther, A.; Ogaya, R.; Oravec, M.; Urban, O.; Peñuelas, J. Topsoil depth substantially influences the responses to drought of the foliar metabolomes of Mediterranean forests. Perspect. Plant Ecol. Evol. Syst. 2016, 21, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Ubach, A.; Sardans, J.; Hódar, J.A.; Garcia-Porta, J.; Guenther, A.; Paša-Tolić, L.; Oravec, M.; Urban, O.; Peñuelas, J. Close and distant: Contrasting the metabolism of two closely related subspecies of Scots pine under the effects of folivory and summer drought. Ecol. Evol. 2017, 7, 8976–8988. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Oravec, M.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; Parella, T.; Peñuelas, J. Warming differentially influences the effects of drought on stoichiometry and metabolomics in shoots and roots. New Phytol. 2015, 207, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Mirnezhad, M.; Romero-González, R.R.; Leiss, K.A.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. Metabolomic analysis of host plant resistance to thrips in wild and cultivated tomatoes. Phytochem. Anal. 2010, 21, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.J.; Allwood, J.W.; Marsden-Edwards, E.; van der Putten, W.H.; Goodacre, R.; van Dam, N.M. Metabolomic analysis of the interaction between plants and herbivores. Metabolomics 2009, 5, 150–161. [Google Scholar] [CrossRef]
- Gómez, S.; Steinbrenner, A.D.; Osorio, S.; Schueller, M.; Ferrieri, R.A.; Fernie, A.R.; Orians, C.M. From shoots to roots: Transport and metabolic changes in tomato after simulated feeding by a specialist lepidopteran. Entomol. Exp. Appl. 2012, 144, 101–111. [Google Scholar] [CrossRef]
- Glauser, G.; Marti, G.; Villard, N.; Doyen, G.A.; Wolfender, J.-L.; Turlings, T.C.J.; Erb, M. Induction and detoxification of maize 1,4-benzoxazin-3-ones by insect herbivores. Plant J. 2011, 68, 901–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-Ubach, A.; Liu, Y.; Bianchi, T.S.; Toli, N.; Jansson, C.; Paša-Tolić, L. Moving beyond the van Krevelen diagram: A new stoichiometric approach for compound classification in organisms. Anal. Chem. 2018, 90, 6152–6160. [Google Scholar] [CrossRef] [PubMed]
- Avtzis, N. Development of Thaumetopoea pityocampa schiff.(Lepidoptera: Thaumetopoeidae) in relation to food consumption. For. Ecol. Manag. 1986, 15, 65–68. [Google Scholar] [CrossRef]
- Battisti, A. Host-plant relationships and population dynamics of the Pine Processionary Caterpillar Thaumetopoea pityocampa (Denis & Schiffermuller). J. Appl. Entomol. 1988, 105, 393–402. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Robledo-Arnuncio, J.J.; Navascués, M.; González-Martínez, S.C.; Gil, L. Estimating gametic introgression rates in a risk assessment context: A case study with Scots pine relicts. Heredity 2009, 103, 385–393. [Google Scholar] [CrossRef]
- Johnson, S.N.; McNicol, J.W. Elevated CO2 and aboveground-belowground herbivory by the clover root weevil. Oecologia 2010, 162, 209–216. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C:N:P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Andersen, T.; Hessen, D.O. Carbon, nitrogen, and phosphorus content of freshwater zooplankton. Limnol. Oceanogr. 1991, 36, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Mayntz, D.; Toft, S. Nutrient composition of the prey’s diet affects growth and survivorship of a generalist predator. Oecologia 2001, 127, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ashhurst, D.E. The connective tissues of insects. Annu. Rev. Entomol. 1968, 13, 45–74. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef]
- Clarkson, D.T.; Hanson, J.B. The Mineral Nutrition of Higher Plants. Annu. Rev. Plant Physiol. 1980, 31, 239–298. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Coll, M.; Vayreda, J.; Rivas-Ubach, A. Stoichiometry of potassium is largely determined by water availability and growth in Catalonian forests. Funct. Ecol. 2012, 26, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Acquisti, C.; Elser, J.J.; Kumar, S. Ecological Nitrogen Limitation Shapes the DNA Composition of Plant Genomes. Mol. Biol. Evol. 2009, 26, 953–956. [Google Scholar] [CrossRef] [Green Version]
- Mathavan, S.; Muthukrishnan, J. Use of larval weight and length as indices of gut content weight in some Lepidopterous larvae. Oecologia 1980, 44, 317–318. [Google Scholar] [CrossRef]
- Hoch, G.; Toffolo, E.P.; Netherer, S.; Battisti, A.; Schopf, A. Survival at low temperature of larvae of the pine processionary moth Thaumetopoea pityocampa from an area of range expansion. Agric. For. Entomol. 2009, 11, 313–320. [Google Scholar] [CrossRef]
- Robinet, C.; Baier, P.; Pennerstorfer, J.; Schopf, A.; Roques, A. Modelling the effects of climate change on the potential feeding activity of Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Notodontidae) in France. Glob. Ecol. Biol. 2007, 16, 460–471. [Google Scholar]
- Després, L.; David, J.-P.; Gallet, C. The evolutionary ecology of insect resistance to plant chemicals. Trends Ecol. Evol. 2007, 22, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R. Sequestration of Defensive Substances from Plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, M.G. Ecological Implications of Minilivestock; CRC Press: Boca Raton, FL, USA, 2005; ISBN 9781482294439. [Google Scholar]
- Banjo, A.; Lawal, O.; Songonuga, E. The nutritional value of fourteen species of edible insects in southwestern Nigeria. Afr. J. Biotechnol. 2006, 5, 298–301. [Google Scholar]
- Sulanc, M.; Emre, I. Effects of B group vitamins and choline chloride on the development and protein synthesis in the male larvae of Pimpla turionellae L. (Hym., Ichneumonidae). J. Appl. Entomol. 2000, 124, 151–153. [Google Scholar] [CrossRef]
- Liu, F.; Wei, F.; Wang, L.; Liu, H.; Zhu, X.; Liang, Y. Riboflavin activates defense responses in tobacco and induces resistance against Phytophthora parasitica and Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2010, 74, 330–336. [Google Scholar] [CrossRef]
- Taheri, P.; Tarighi, S. Riboflavin induces resistance in rice against Rhizoctonia solani via jasmonate-mediated priming of phenylpropanoid pathway. J. Plant Physiol. 2010, 167, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Verdrengh, M.; Tarkowski, A. Riboflavin in innate and acquired immune responses. Inflamm. Res. 2005, 54, 390–393. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Stefanescu, C.; Parella, T.; Filella, I. Lonicera Implexa Leaves Bearing Naturally Laid Eggs of the Specialist Herbivore Euphydryas Aurinia have Dramatically Greater Concentrations of Iridoid Glycosides than other Leaves. J. Chem. Ecol. 2006, 32, 1925–1933. [Google Scholar] [CrossRef]
- Gols, R.; Harvey, J.A. Plant-mediated effects in the Brassicaceae on the performance and behaviour of parasitoids. Phytochem. Rev. 2009, 8, 187–206. [Google Scholar] [CrossRef]
- Achotegui-Castells, A.; Llusia, J.; Hódar, J.A.; Peñuelas, J. Needle terpene concentrations and emissions of two coexisting subspecies of Scots pine attacked by the pine processionary moth (Thaumetopoea pityocampa). Acta Physiol. Plant. 2013, 35, 3047–3058. [Google Scholar] [CrossRef]
- Rajsnerová, P.; Klem, K.; Holub, P.; Novotná, K.; Večeřová, K.; Kozáčiková, M.; Rivas-Ubach, A.; Sardans, J.; Marek, M.V.; Peñuelas, J.; et al. Morphological, biochemical and physiological traits of upper and lower canopy leaves of European beech tend to converge with increasing altitude. Tree Phys. 2015, 1, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Montes, F.; Peñuelas, J. Determination of As, Cd, Cu, Hg and Pb in biological samples by modern electrothermal atomic absorption spectrometry. Spectrochim. Acta Part B At. Spectrosc. 2010, 65, 97–112. [Google Scholar] [CrossRef]
- t’Kindt, R.; De Veylder, L.; Storme, M.; Deforce, D.; Van Bocxlaer, J. LC-MS metabolic profiling of Arabidopsis thaliana plant leaves and cell cultures: Optimization of pre-LC-MS procedure parameters. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2008, 871, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Ubach, A.; Hódar, J.A.; Sardans, J.; Kyle, J.E.; Kim, Y.-M.; Oravec, M.; Urban, O.; Guenther, A.; Peñuelas, J. Are the metabolomic responses to folivory of closely related plant species linked to macroevolutionary and plant-folivore coevolutionary processes? Ecol. Evol. 2016, 6, 4372–4386. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based metabolomic analysis of plants. Nat. Protoc. 2010, 5, 536–549. [Google Scholar] [CrossRef]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Pérez-Trujillo, M.; Sardans, J.; Gargallo-Garriga, A.; Parella, T.; Peñuelas, J. Ecometabolomics: Optimized NMR-based method. Methods Ecol. Evol. 2013, 4, 464–473. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria, 2013. Available online: http://www.R-project.org (accessed on 22 December 2018).
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R package Version 1.2-3. 2015. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 22 December 2018).
- Harrell, F.E. Hmisc: Harrell Miscellaneous. R Package Version 4.1-1. 2018. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 22 December 2018).
- Oksanen, J.; Guillaume-Blanchet, F.; Fierndly, M.; Kindt, R.; Legendre, P.; McGlinnd, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.1-1. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 22 December 2018).
- Husson, F.; Josse, J.; Le, S.; Mazet, J. FactoMineR: Multivariate Exploratory Data Analysis and Data Mining. R Package Version 1.32. 2016. Available online: https://CRAN.R-project.org/package=FactoMineR (accessed on 22 December 2018).
- Husson, F.; Josse, J. missMDA: Handling Missing Values with Multivariate Data Analysis. R Package Version 1.9. 2015. Available online: https://CRAN.R-project.org/package=missMDA (accessed on 22 December 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Class | Compound Name | Averaged Relative Abundance (Deconvoluted Ion Chromatogram Peak Area) | |
|---|---|---|---|
| Caterpillars | Pine Needles | ||
| Carbohydrates | Hexoses (Glucose, Fructose, Mannose, Galactose, …) | 37,234,258 | 75,422,781 |
| Pentoses (Arabinose, Ribose, Xylose, …) | 9,983,894 | 12,228,327 | |
| Deoxy-Hexoses (Deoxy-glucose, Deoxy-galactose, Fucose, …) | 592,288.1 | 52,9697.9 | |
| Hexoses-OH (Sorbitol, Mannitol, …) | 3,303,206 | 69,191.52 | |
| Pentoses-OH (Xylitol, Arabitol, …) | 64,255,464 | 29,779,498 | |
| Amino acids | Alanine | 2.55 × 108 | 14,910,060 |
| Arginine | 73,078,590 | 10,791,686 | |
| Aspartic acid | 1,060,014 | 603,470.4 | |
| Glutamic acid | 21,812,174 | 9,902,287 | |
| Glutamine | 18,562,019 | 35,001,363 | |
| Glycine | 9,044,821 | 3,829,374 | |
| Isoleucine | 4.38 × 109 | 4.98 × 108 | |
| Leucine | 55,187,379 | 1,556,441 | |
| Lysine | 3.12 × 108 | 5,959,588 | |
| Methionine | 1.76 × 108 | 65,552,885 | |
| Phenylalanine | 2.24 × 109 | 2 × 108 | |
| Proline | 1.5 × 109 | 53,596,908 | |
| Serine | 10,026,643 | 7,239,244 | |
| Threonine | 18,231,142 | 8,067,142 | |
| Tyrosine | 2.14 × 108 | 24,445,586 | |
| Tryptophan | 2.03 × 109 | 4.8 × 108 | |
| Valine | 4.39 × 108 | 47,193,674 | |
| Nucleobases | Adenine | 1,422,770 | 41,186,385 |
| Guanine | 428,695.9 | 281,113.5 | |
| Thymine | 2,393,343 | 19,099,857 | |
| Uracil | 5,393,886 | 3,712,866 | |
| Nucleosides | Adenosine | 1,733,495 | 1,297,581 |
| Krebs cycle related organic acids | Citric acid | 4,984,835 | 76,318,569 |
| Malic acid | 6,556,566 | 93,520,587 | |
| Pyruvic ac | 2,293,721 | 8,504,367 | |
| Succinic ac | 1.68 × 108 | 740,971.5 | |
| α-Ketoglutaric acid | 37,118.36 | 235,423.5 | |
| Terpenes | Caryophyllene | 13,334,699 | |
| Sabinene | 2,857,821 | ||
| Phenolics | Apigenin | 21,878,949 | |
| Catechin | 1.54 × 108 | ||
| Catechol | 39,448,216 | ||
| d-Pinitol | 34,467,901 | 30,806,094 | |
| Epicatechin | 219,812.1 | ||
| Epigallocatechin | 376,194.4 | ||
| Ferulic acid | 754,250.3 | 972,992.9 | |
| Kaempferol | 1,199,232 | ||
| Luteolin | 442,780.3 | ||
| Phenil-phenol | 883,635.4 | 110,703.6 | |
| Quercetin | 1,089,097 | 805,365.7 | |
| Quinic acid | 52,693,061 | 4.26 × 108 | |
| Taxifolin | 8,041,666 | ||
| Vanillic acid | 917,952.3 | 457,177.3 | |
| Other | Abscisic acid | 502,407.6 | 3,853,610 |
| Choline | 2.44 × 108 | 67,441,330 | |
| δ-tocopherol | 3,376,661 | ||
| Gibberellic acid 3 | 1,050,073 | ||
| Vitamin B5 (Panthotenic acid) | 38,952,234 | 1,464,870 | |
| Vitamin B2 (Riboflavin) | 6,437,087 | 94,010.85 | |
| Shikimic acid | 26,206,952 | 1.27 × 108 | |
| Source of Variation | Degrees of Freedom | Sums of Squares | Mean Square | Pseudo-F | R2 | p Value |
|---|---|---|---|---|---|---|
| Caterpillar group | 1 | 1.30 × 1019 | 1.30 × 1019 | 2.524 | 0.103 | 0.0223 |
| Residuals | 22 | 1.13 × 1020 | 5.14 × 1018 | 0.897 | ||
| Total | 23 | 1.26 × 1020 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivas-Ubach, A.; Peñuelas, J.; Hódar, J.A.; Oravec, M.; Paša-Tolić, L.; Urban, O.; Sardans, J. We Are What We Eat: A Stoichiometric and Ecometabolomic Study of Caterpillars Feeding on Two Pine Subspecies of Pinus sylvestris. Int. J. Mol. Sci. 2019, 20, 59. https://doi.org/10.3390/ijms20010059
Rivas-Ubach A, Peñuelas J, Hódar JA, Oravec M, Paša-Tolić L, Urban O, Sardans J. We Are What We Eat: A Stoichiometric and Ecometabolomic Study of Caterpillars Feeding on Two Pine Subspecies of Pinus sylvestris. International Journal of Molecular Sciences. 2019; 20(1):59. https://doi.org/10.3390/ijms20010059
Chicago/Turabian StyleRivas-Ubach, Albert, Josep Peñuelas, José Antonio Hódar, Michal Oravec, Ljiljana Paša-Tolić, Otmar Urban, and Jordi Sardans. 2019. "We Are What We Eat: A Stoichiometric and Ecometabolomic Study of Caterpillars Feeding on Two Pine Subspecies of Pinus sylvestris" International Journal of Molecular Sciences 20, no. 1: 59. https://doi.org/10.3390/ijms20010059