
Magnetophotoselection in the Investigation of Excitonically Coupled Chromophores: The Case of the Water-Soluble Chlorophyll Protein

 , , , and
, , , and
Abstract
:1. Introduction

2. Materials and Methods
2.1. Sample Preparation
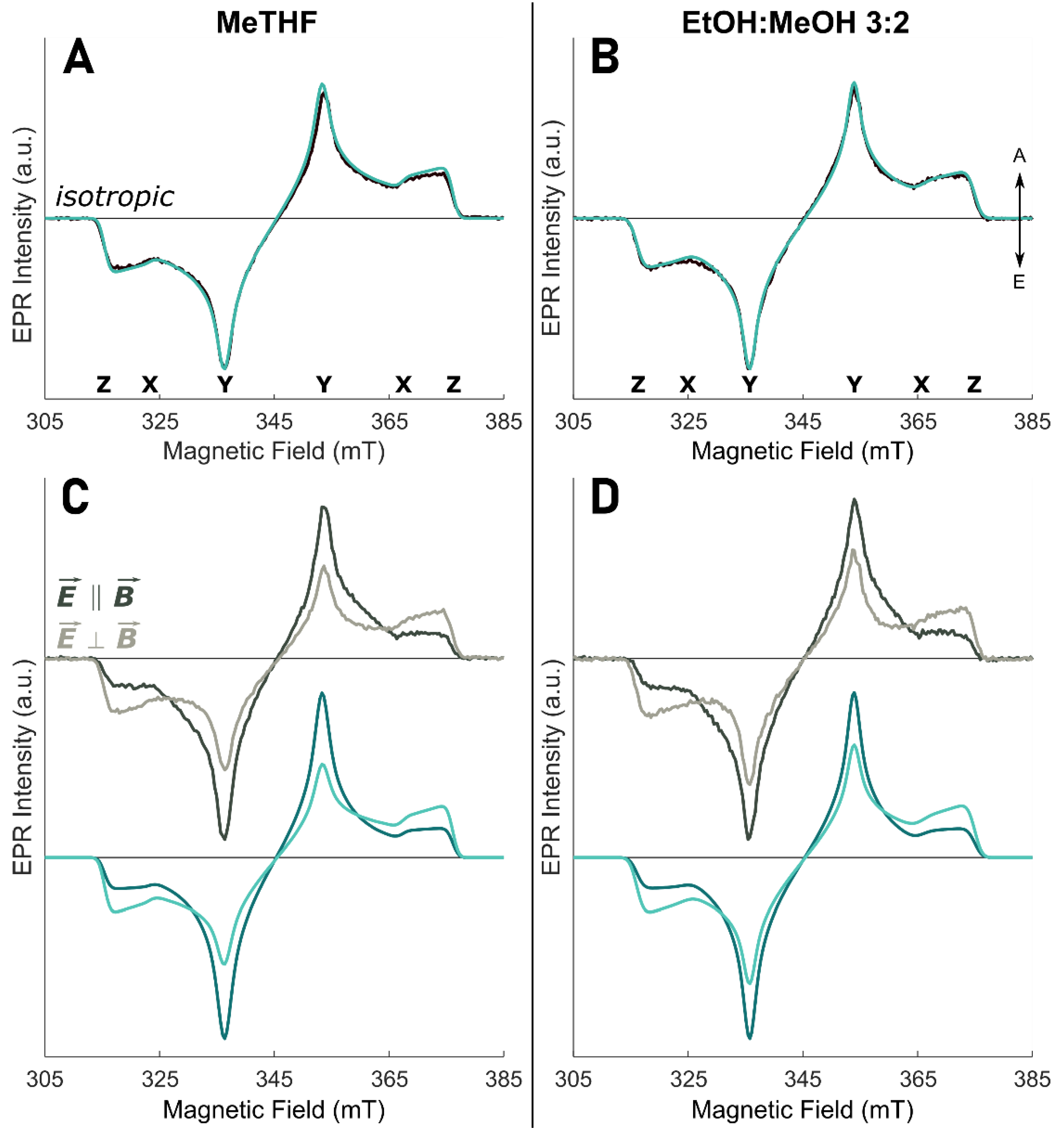
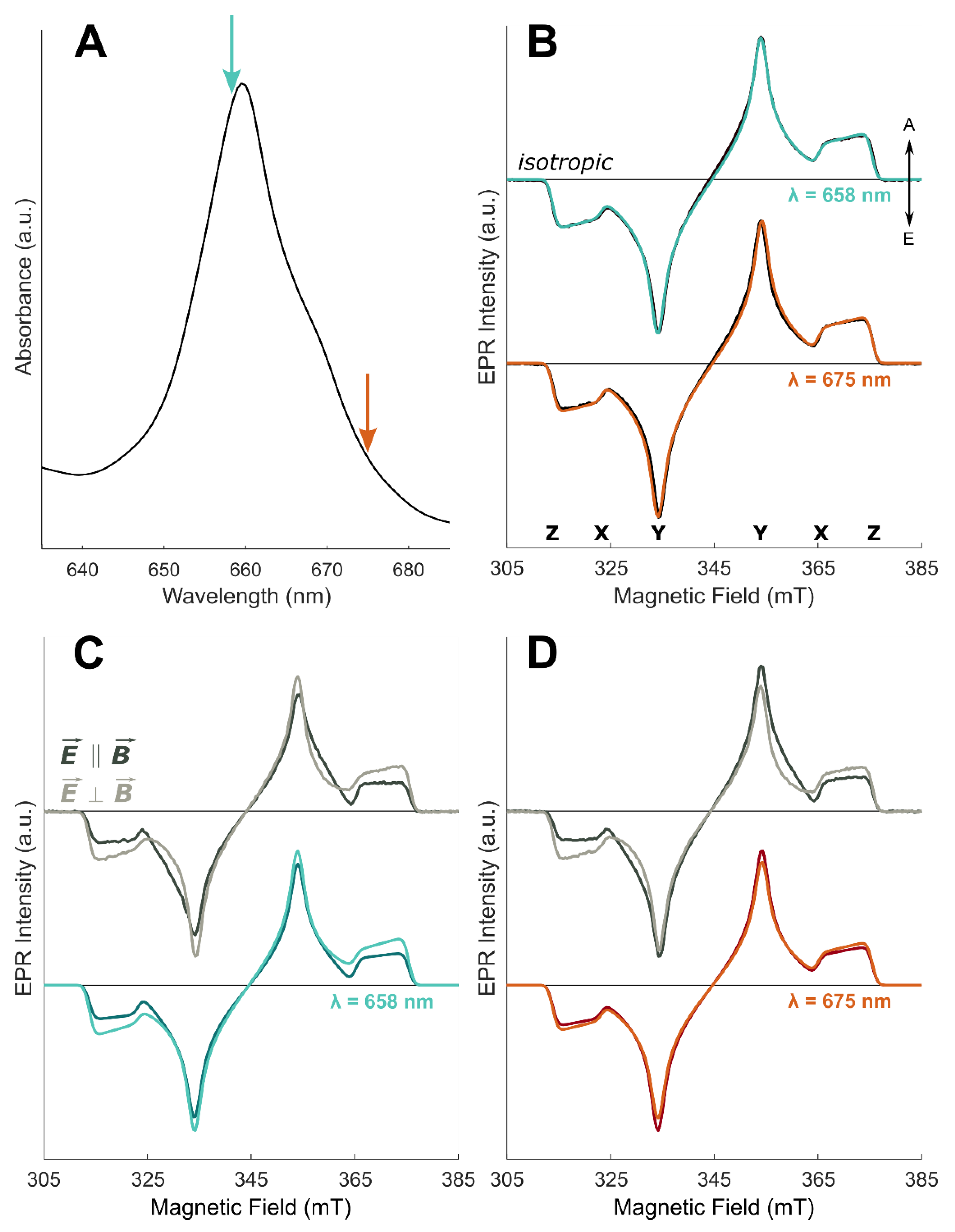
2.2. TR-EPR Experiments
2.3. Spectral Analysis
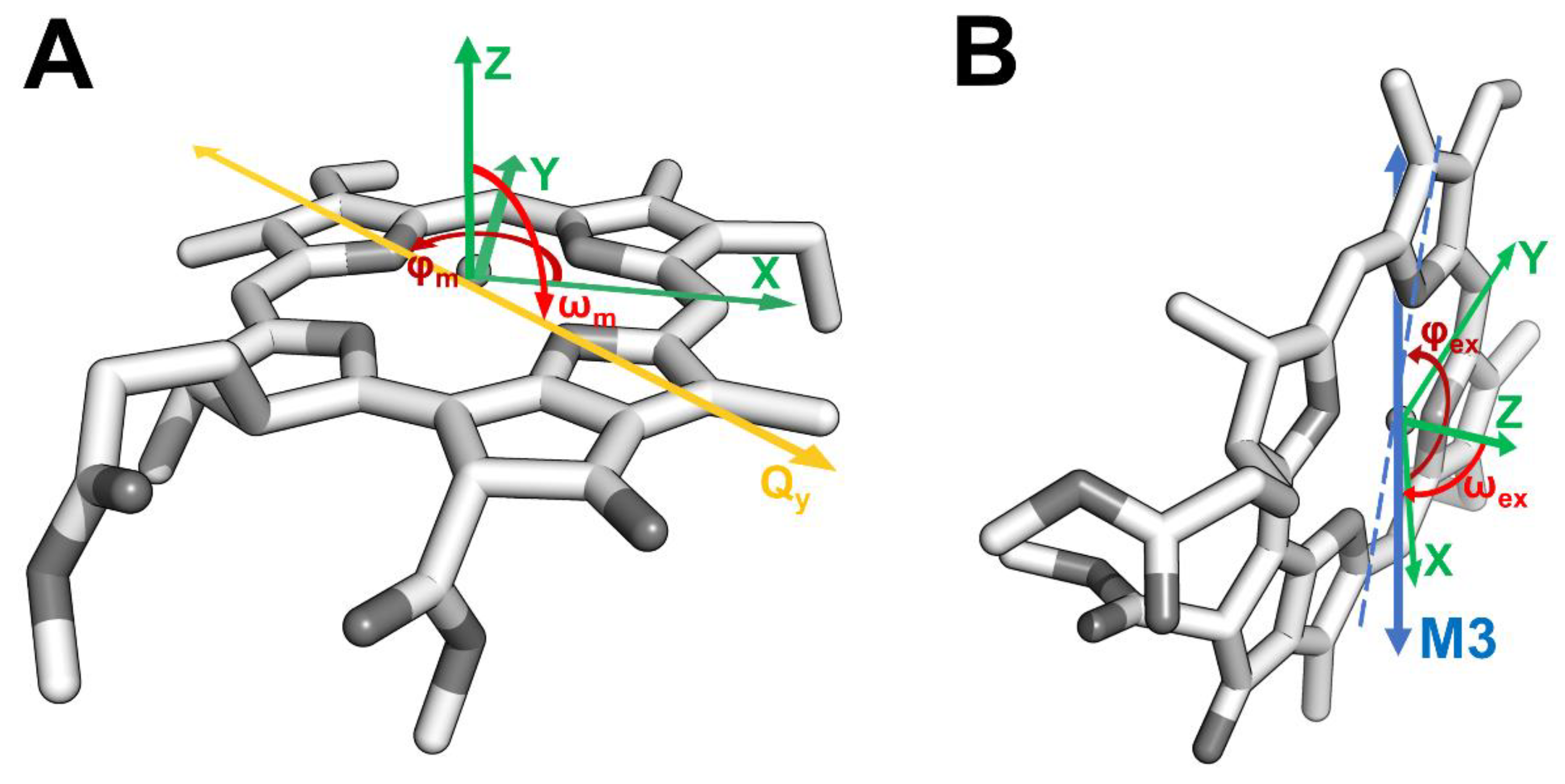
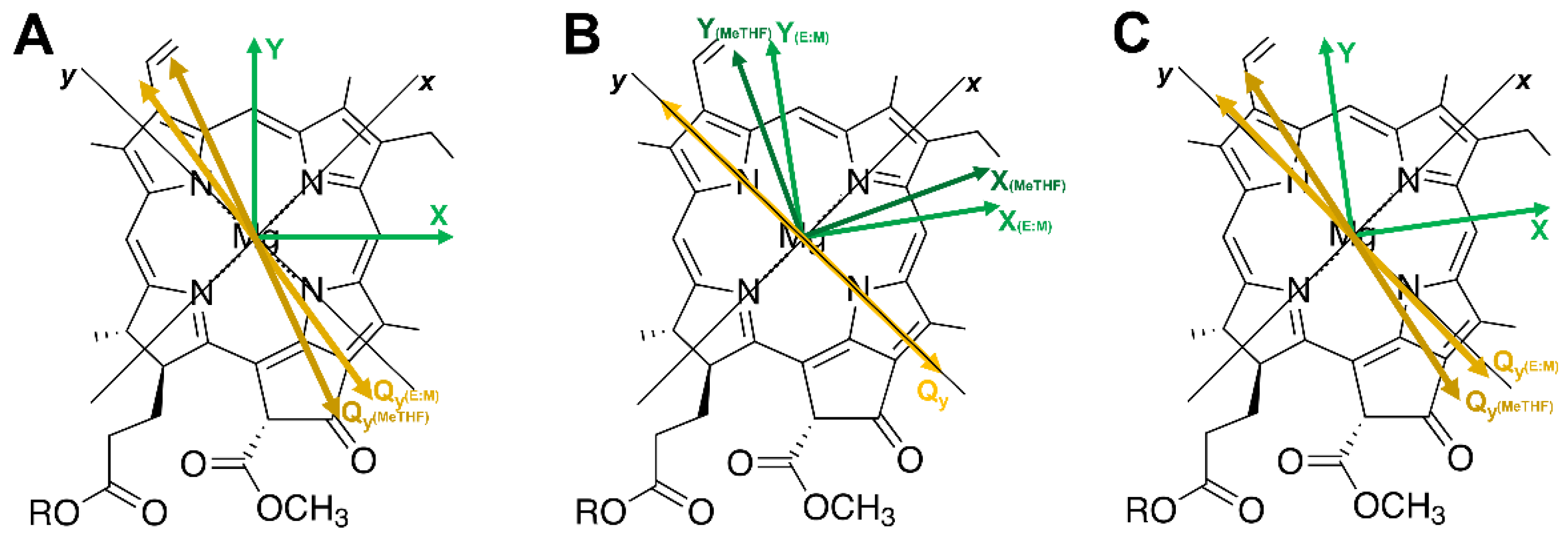
2.4. Excitonic Coupling Calculation
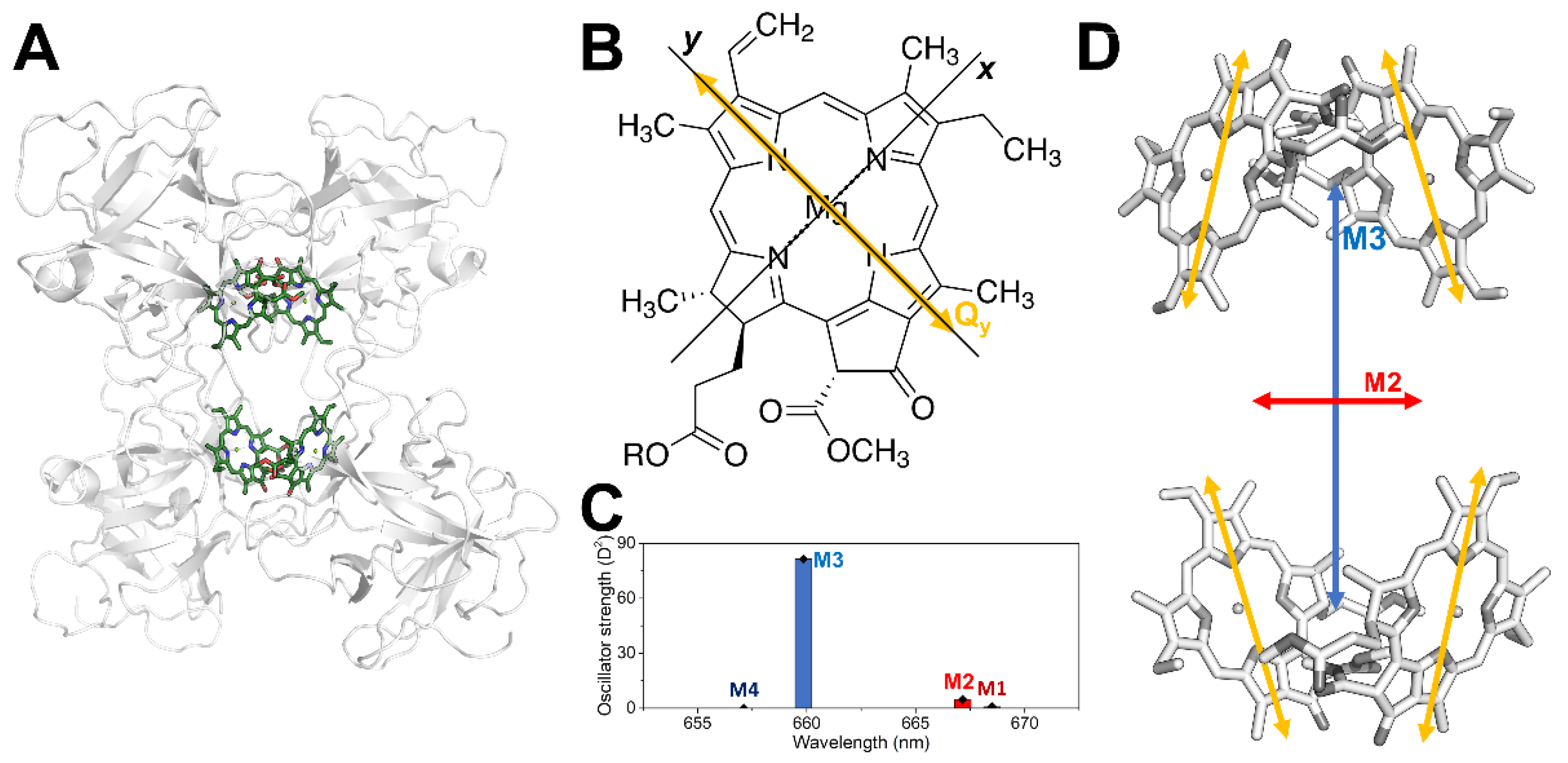
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Scholes, G.D.; Fleming, G.R.; Olaya-Castro, A.; van Grondelle, R. Lessons from Nature about Solar Light Harvesting. Nat. Chem. 2011, 3, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Croce, R.; van Amerongen, H. Natural Strategies for Photosynthetic Light Harvesting. Nat. Chem. Biol. 2014, 10, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Battersby, A.R. Tetrapyrroles: The Pigments of Life. Nat. Prod. Rep. 2000, 17, 507–526. [Google Scholar] [CrossRef]
- Björn, L.O.; Papageorgiou, G.C.; Blankenship, R.E.; Govindjee. A Viewpoint: Why Chlorophyll a? Photosynth. Res. 2009, 99, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Mauzerall, D. Why Chlorophyll? Ann. N. Y. Acad. Sci. 1973, 206, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Gouterman, M. Spectra of Porphyrins. J. Mol. Spectrosc. 1961, 6, 138–163. [Google Scholar] [CrossRef]
- Fragata, M.; Nordén, B.; Kurucsev, T. Linear Dichroism (250–700 nm) of Chlorophyll a and Pheophytin a Oriented in a Lamellar Phase of Glycerylmonooctanoate/H2O. Characterization of Electronic Transitions. Photochem. Photobiol. 1988, 47, 133–143. [Google Scholar] [CrossRef]
- Norden, B.; Fragata, M.; Kurucsev, T. X- and Y-Polarized Spectra of Chlorophyll a and Pheophytin a in the Red Region: Resolution Enhancement and Gaussian Deconvolution. Aust. J. Chem. 1992, 45, 1559. [Google Scholar] [CrossRef]
- Zandvoort, M.V.; Wróbel, D.; Lettinga, P.; Ginkel, G.V.; Levine, Y.K. The Orientation of the Transition Dipole Moments of Chlorophyll a and Pheophytin a in Their Molecular Frame. Photochem. Photobiol. 1995, 62, 299–308. [Google Scholar] [CrossRef]
- Linke, M.; Lauer, A.; von Haimberger, T.; Zacarias, A.; Heyne, K. Three-Dimensional Orientation of the Qy Electronic Transition Dipole Moment within the Chlorophyll a Molecule Determined by Femtosecond Polarization Resolved VIS Pump−IR Probe Spectroscopy. J. Am. Chem. Soc. 2008, 130, 14904–14905. [Google Scholar] [CrossRef]
- Oviedo, M.B.; Sánchez, C.G. Transition Dipole Moments of the Qy Band in Photosynthetic Pigments. J. Phys. Chem. A 2011, 115, 12280–12285. [Google Scholar] [CrossRef]
- Hedayatifar, L.; Irani, E.; Mazarei, M.; Rasti, S.; Azar, Y.T.; Rezakhani, A.T.; Mashaghi, A.; Shayeganfar, F.; Anvari, M.; Heydari, T.; et al. Optical Absorption and Electronic Spectra of Chlorophylls a and b. RSC Adv. 2016, 6, 109778–109785. [Google Scholar] [CrossRef] [Green Version]
- Madjet, M.E.; Abdurahman, A.; Renger, T. Intermolecular Coulomb Couplings from Ab Initio Electrostatic Potentials: Application to Optical Transitions of Strongly Coupled Pigments in Photosynthetic Antennae and Reaction Centers. J. Phys. Chem. B 2006, 110, 17268–17281. [Google Scholar] [CrossRef] [PubMed]
- Friedl, C.; Fedorov, D.G.; Renger, T. Towards a Quantitative Description of Excitonic Couplings in Photosynthetic Pigment–Protein Complexes: Quantum Chemistry Driven Multiscale Approaches. Phys. Chem. Chem. Phys. 2022, 24, 5014–5038. [Google Scholar] [CrossRef] [PubMed]
- Sirohiwal, A.; Berraud-Pache, R.; Neese, F.; Izsák, R.; Pantazis, D.A. Accurate Computation of the Absorption Spectrum of Chlorophyll a with Pair Natural Orbital Coupled Cluster Methods. J. Phys. Chem. B 2020, 124, 8761–8771. [Google Scholar] [CrossRef]
- Agostini, A.; Palm, D.M.; Paulsen, H.; Carbonera, D. Optically Detected Magnetic Resonance of Chlorophyll Triplet States in Water-Soluble Chlorophyll Proteins from Lepidium virginicum: Evidence for Excitonic Interaction among the Four Pigments. J. Phys. Chem. B 2018, 122, 6156–6163. [Google Scholar] [CrossRef] [PubMed]
- Renger, G.; Pieper, J.; Theiss, C.; Trostmann, I.; Paulsen, H.; Renger, T.; Eichler, H.J.; Schmitt, F.-J. Water Soluble Chlorophyll Binding Protein of Higher Plants: A Most Suitable Model System for Basic Analyses of Pigment–Pigment and Pigment–Protein Interactions in Chlorophyll Protein Complexes. J. Plant Physiol. 2011, 168, 1462–1472. [Google Scholar] [CrossRef]
- Satoh, H.; Uchida, A.; Nakayama, K.; Okada, M. Water-Soluble Chlorophyll Protein in Brassicaceae Plants Is a Stress-Induced Chlorophyll-Binding Protein. Plant Cell Physiol. 2001, 42, 906–911. [Google Scholar] [CrossRef]
- Girr, P.; Paulsen, H. How Water-Soluble Chlorophyll Protein Extracts Chlorophyll from Membranes. Biochim. Biophys. Acta—Biomembr. 2021, 1863, 183479. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Yanai, H.; Oka-Takayama, Y.; Zanma-Sohtome, A.; Fujiyama, K.; Uchida, A.; Nakayama, K.; Satoh, H. Molecular Cloning, Characterization and Analysis of the Intracellular Localization of a Water-Soluble Chlorophyll-Binding Protein (WSCP) from Virginia Pepperweed (Lepidium virginicum), a Unique WSCP That Preferentially Binds Chlorophyll b in Vitro. Planta 2013, 238, 1065–1080. [Google Scholar] [CrossRef]
- Takahashi, S.; Yanai, H.; Nakamaru, Y.; Uchida, A.; Nakayama, K.; Satoh, H. Molecular Cloning, Characterization and Analysis of the Intracellular Localization of a Water-Soluble Chl-Binding Protein from Brussels Sprouts (Brassica oleracea Var. Gemmifera). Plant Cell Physiol. 2012, 53, 879–891. [Google Scholar] [CrossRef]
- Boex-Fontvieille, E.; Rustgi, S.; von Wettstein, D.; Pollmann, S.; Reinbothe, S.; Reinbothe, C. An Ethylene-Protected Achilles’ Heel of Etiolated Seedlings for Arthropod Deterrence. Front. Plant Sci. 2016, 7, 1246. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.; Kroymann, J.; Mitchell-Olds, T. Different Transcript Patterns in Response to Specialist and Generalist Herbivores in the Wild Arabidopsis Relative Boechera divaricarpa. PLoS ONE 2007, 2, e1081. [Google Scholar] [CrossRef] [Green Version]
- Lopez, F.; Vansuyt, G.; Fourcroy, P.; Casse-Delbart, F. Accumulation of a 22-KDa Protein and Its MRNA in the Leaves of Raphanus sativus in Response to Salt Stress or Water Deficit. Physiol. Plant. 1994, 91, 605–614. [Google Scholar] [CrossRef]
- Nishio, N.; Satoh, H. A Water-Soluble Chlorophyll Protein in Cauliflower May Be Identical to BnD22, a Drought-Induced, 22-Kilodalton Protein in Rapeseed. Plant Physiol. 1997, 115, 841–846. [Google Scholar] [CrossRef] [Green Version]
- Boex-Fontvieille, E.; Rustgi, S.; Von Wettstein, D.; Reinbothe, S.; Reinbothe, C. Water-Soluble Chlorophyll Protein is Involved in Herbivore Resistance Activation during Greening of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 112, 7303–7308. [Google Scholar] [CrossRef] [Green Version]
- Agostini, A.; Palm, D.M.; Schmitt, F.-J.; Albertini, M.; Di Valentin, M.; Paulsen, H.; Carbonera, D. An Unusual Role for the Phytyl Chains in the Photoprotection of the Chlorophylls Bound to Water-Soluble Chlorophyll-Binding Proteins. Sci. Rep. 2017, 7, 7504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boex-Fontvieille, E.; Rustgi, S.; Reinbothe, S.; Reinbothe, C. A Kunitz-Type Protease Inhibitor Regulates Programmed Cell Death during Flower Development in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 6119–6135. [Google Scholar] [CrossRef] [Green Version]
- Bednarczyk, D.; Dym, O.; Prabahar, V.; Peleg, Y.; Pike, D.H.; Noy, D. Chlorophyll Fine Tuning of Chlorophyll Spectra by Protein-Induced Ring Deformation. Angew. Chem. Int. Ed. 2016, 55, 6901–6905. [Google Scholar] [CrossRef]
- Agostini, A.; Meneghin, E.; Gewehr, L.; Pedron, D.; Palm, D.M.; Carbonera, D.; Paulsen, H.; Jaenicke, E.; Collini, E. How Water-Mediated Hydrogen Bonds Affect Chlorophyll a/b Selectivity in Water-Soluble Chlorophyll Protein. Sci. Rep. 2019, 9, 18255. [Google Scholar] [CrossRef]
- Horigome, D.; Satoh, H.; Itoh, N.; Mitsunaga, K.; Oonishi, I.; Nakagawa, A.; Uchida, A. Structural Mechanism and Photoprotective Function of Water-Soluble Chlorophyll-Binding Protein. J. Biol. Chem. 2007, 282, 6525–6531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renger, T.; Trostmann, I.; Theiss, C.; Madjet, M.E.; Richter, M.; Paulsen, H.; Eichler, H.J.; Knorr, A.; Renger, G. Refinement of a Structural Model of a Pigment-Protein Complex by Accurate Optical Line Shape Theory and Experiments. J. Phys. Chem. B 2007, 111, 10487–10501. [Google Scholar] [CrossRef]
- Hughes, J.L.; Razeghifard, R.; Logue, M.; Oakley, A.; Wydrzynski, T.; Krausz, E. Magneto-Optic Spectroscopy of a Protein Tetramer Binding Two Exciton-Coupled Chlorophylls. J. Am. Chem. Soc. 2006, 128, 3649–3658. [Google Scholar] [CrossRef]
- Rancova, O.; Jakučionis, M.; Valkunas, L.; Abramavicius, D. Origin of Non-Gaussian Site Energy Disorder in Molecular Aggregates. Chem. Phys. Lett. 2017, 674, 120–124. [Google Scholar] [CrossRef]
- Theiss, C.; Trostmann, I.; Andree, S.; Schmitt, F.J.; Renger, T.; Eichler, H.J.; Paulsen, H.; Renger, G. Pigment−Pigment and Pigment−Protein Interactions in Recombinant Water-Soluble Chlorophyll Proteins (WSCP) from Cauliflower. J. Phys. Chem. B 2007, 111, 13325–13335. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, F.-J.; Trostmann, I.; Theiss, C.; Pieper, J.; Renger, T.; Fuesers, J.; Hubrich, E.H.; Paulsen, H.; Eichler, H.J.; Renger, G. Excited State Dynamics in Recombinant Water-Soluble Chlorophyll Proteins (WSCP) from Cauliflower Investigated by Transient Fluorescence Spectroscopy. J. Phys. Chem. B 2008, 112, 13951–13961. [Google Scholar] [CrossRef]
- Renger, T.; Madjet, M.E.; Müh, F.; Trostmann, I.; Schmitt, F.-J.; Theiss, C.; Paulsen, H.; Eichler, H.J.; Knorr, A.; Renger, G. Thermally Activated Superradiance and Intersystem Crossing in the Water-Soluble Chlorophyll Binding Protein. J. Phys. Chem. B 2009, 113, 9948–9957. [Google Scholar] [CrossRef]
- Adolphs, J.; Berrer, M.; Renger, T. Hole-Burning Spectroscopy on Excitonically Coupled Pigments in Proteins: Theory Meets Experiment. J. Am. Chem. Soc. 2016, 138, 2993–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alster, J.; Lokstein, H.; Dostál, J.; Uchida, A.; Zigmantas, D. 2D Spectroscopy Study of Water-Soluble Chlorophyll-Binding Protein from Lepidium virginicum. J. Phys. Chem. B 2014, 118, 3524–3531. [Google Scholar] [CrossRef]
- Pieper, J.; Rätsep, M.; Trostmann, I.; Schmitt, F.-J.; Theiss, C.; Paulsen, H.; Eichler, H.J.; Freiberg, A.; Renger, G. Excitonic Energy Level Structure and Pigment−Protein Interactions in the Recombinant Water-Soluble Chlorophyll Protein. II. Spectral Hole-Burning Experiments. J. Phys. Chem. B 2011, 115, 4053–4065. [Google Scholar] [CrossRef]
- Pieper, J.; Rätsep, M.; Trostmann, I.; Paulsen, H.; Renger, G.; Freiberg, A. Excitonic Energy Level Structure and Pigment−Protein Interactions in the Recombinant Water-Soluble Chlorophyll Protein. I. Difference Fluorescence Line-Narrowing. J. Phys. Chem. B 2011, 115, 4042–4052. [Google Scholar] [CrossRef]
- Kell, A.; Bednarczyk, D.; Acharya, K.; Chen, J.; Noy, D.; Jankowiak, R. New Insight into the Water-Soluble Chlorophyll-Binding Protein from Lepidium virginicum. Photochem. Photobiol. 2016, 92, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Fresch, E.; Meneghin, E.; Agostini, A.; Paulsen, H.; Carbonera, D.; Collini, E. How the Protein Environment Can Tune the Energy, the Coupling, and the Ultrafast Dynamics of Interacting Chlorophylls: The Example of the Water-Soluble Chlorophyll Protein. J. Phys. Chem. Lett. 2020, 11, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Agostini, A.; Palm, D.M.; Paulsen, H.; Carbonera, D. Accessibility of Protein-Bound Chlorophylls Probed by Dynamic Electron Polarization. J. Phys. Chem. Lett. 2018, 9, 672–676. [Google Scholar] [CrossRef]
- Lahav, Y.; Noy, D.; Schapiro, I. Spectral Tuning of Chlorophylls in Proteins—Electrostatics vs. Ring Deformation. Phys. Chem. Chem. Phys. 2021, 23, 6544–6551. [Google Scholar] [CrossRef] [PubMed]
- Agostini, A.; Palm, D.M.; Paulsen, H.; Di Valentin, M.; Carbonera, D. Electron Nuclear Double Resonance of the Chlorophyll Triplet State in the Water-Soluble Chlorophyll Protein from Brassica oleracea: Investigation of the Effect of the Binding Site on the Hyperfine Couplings. Appl. Magn. Reson. 2020, 51, 925–937. [Google Scholar] [CrossRef]
- Srivastava, A.; Ahad, S.; Wat, J.H.; Reppert, M. Accurate Prediction of Mutation-Induced Frequency Shifts in Chlorophyll Proteins with a Simple Electrostatic Model. J. Chem. Phys. 2021, 155, 151102. [Google Scholar] [CrossRef]
- Croce, R. Beyond ‘Seeing Is Believing’: The Antenna Size of the Photosystems in Vivo. New Phytol. 2020, 228, 1214–1218. [Google Scholar] [CrossRef]
- Forbes, M.D.E.; Jarocha, L.E.; Sim, S.; Tarasov, V.F. Time-Resolved Electron Paramagnetic Resonance Spectroscopy. In Advances in Physical Organic Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1–83. [Google Scholar]
- Richert, S.; Tait, C.E.; Timmel, C.R. Delocalisation of Photoexcited Triplet States Probed by Transient EPR and Hyperfine Spectroscopy. J. Magn. Reson. 2017, 280, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Weber, S. Transient EPR. In EPR Spectroscopy: Fundamentals and Methods; Goldfarb, D., Stoll, S., Eds.; Wiley: Weinheim, Germany, 2018; pp. 195–214. ISBN 978-1-119-16298-8. [Google Scholar]
- Kottis, P.; Lefebvre, R. Calculation of the Electron Spin Resonance Line Shape of Randomly Oriented Molecules in a Triplet State. III. Spectra of Photoselected Molecules. J. Chem. Phys. 1964, 41, 3660–3661. [Google Scholar] [CrossRef]
- Ciuti, S.; Barbon, A.; Bortolus, M.; Agostini, A.; Bergantino, E.; Martin, C.; Di Valentin, M.; Carbonera, D. Neuroglobin Provides a Convenient Scaffold to Investigate the Triplet-State Properties of Porphyrins by Time-Resolved EPR Spectroscopy and Magnetophotoselection. Appl. Magn. Reson. 2021. [Google Scholar] [CrossRef]
- Barbon, A.; Dal Farra, M.G.; Ciuti, S.; Albertini, M.; Bolzonello, L.; Orian, L.; Di Valentin, M. Comprehensive Investigation of the Triplet State Electronic Structure of Free-Base 5,10,15,20-Tetrakis(4-Sulfonatophenyl)Porphyrin by a Combined Advanced EPR and Theoretical Approach. J. Chem. Phys. 2020, 152, 034201. [Google Scholar] [CrossRef] [PubMed]
- Redman, A.J.; Moise, G.; Richert, S.; Viere, E.J.; Myers, W.K.; Therien, M.J.; Timmel, C.R. EPR of Photoexcited Triplet-State Acceptor Porphyrins. J. Phys. Chem. C 2021, 125, 11782–11790. [Google Scholar] [CrossRef]
- Tait, C.E.; Neuhaus, P.; Anderson, H.L.; Timmel, C.R. Triplet State Delocalization in a Conjugated Porphyrin Dimer Probed by Transient Electron Paramagnetic Resonance Techniques. J. Am. Chem. Soc. 2015, 137, 6670–6679. [Google Scholar] [CrossRef]
- Thurnauer, M.C.; Norris, J.R. The Ordering of the Zero field Triplet Spin Sublevels in the Chlorophylls. A Magnetophotoselection Study. Chem. Phys. Lett. 1977, 47, 100–105. [Google Scholar] [CrossRef]
- Trosper, T.L.; Frank, H.A.; Norris, J.R.; Thurnauer, M.C. Magnetophotoselection Studies on Rhodopseudomonas viridis Reaction Centers. Biochim. Biophys. Acta—Bioenerg. 1982, 679, 44–50. [Google Scholar] [CrossRef]
- Boxer, S.G.; Roelofs, M.G. Chromophore Organization in Photosynthetic Reaction Centers: High-Resolution Magnetophotoselection. Proc. Natl. Acad. Sci. USA 1979, 76, 5636–5640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, H.A.; Bolt, J.; Friesner, R.; Sauer, K. Magnetophotoselection of the Triplet State of Reaction Centers from Rhodopseudomonas sphaeroides R-26. Biochim. Biophys. Acta Bioenerg. 1979, 547, 502–511. [Google Scholar] [CrossRef] [Green Version]
- Thurnauer, M.C.; Norris, J.R. Magnetophotoselection Applied to the Triplet State Observed by EPR in Photosynthetic Bacteria. Biochem. Biophys. Res. Commun. 1976, 73, 501–506. [Google Scholar] [CrossRef]
- McGann, W.J.; Frank, H.A. Magnetophotoselection of Rhodopseudomonas sphaeroides Wild-Type Reaction Centers. Biochim. Biophys. Acta—Bioenerg. 1985, 807, 101–109. [Google Scholar] [CrossRef]
- Borovykh, I.V.; Proskuryakov, I.I.; Klenina, I.B.; Gast, P.; Hoff, A.J. Magnetophotoselection Study of the Lowest Excited Triplet State of the Primary Donor in Photosynthetic Bacteria. J. Phys. Chem. B 2000, 104, 4222–4228. [Google Scholar] [CrossRef]
- Bosch, M.K.; Proskuryakov, I.I.; Gast, P.; Hoff, A.J. Relative Orientation of the Optical Transition Dipole and Triplet Axes of the Photosystem II Primary Donor. A Magnetophotoselection Study. J. Phys. Chem. 1995, 99, 15310–15316. [Google Scholar] [CrossRef]
- Vrieze, J.; Hoff, A.J. The Orientation of the Triplet Axes with Respect to the Optical Transition Moments in (Bacterio) Chlorophylls. Chem. Phys. Lett. 1995, 237, 493–501. [Google Scholar] [CrossRef]
- Lendzian, F.; Bittl, R.; Telfer, A.; Lubitz, W. Hyperfine Structure of the Photoexcited Triplet State 3P680 in Plant PS II Reaction Centres as Determined by Pulse ENDOR Spectroscopy. Biochim. Biophys. Acta—Bioenerg. 2003, 1605, 35–46. [Google Scholar] [CrossRef]
- Booth, P.J.; Paulsen, H. Assembly of Light-Harvesting Chlorophyll a/b Complex in Vitro. Time-Resolved Fluorescence Measurements. Biochemistry 1996, 35, 5103–5108. [Google Scholar] [CrossRef]
- Palm, D.M.; Agostini, A.; Averesch, V.; Girr, P.; Werwie, M.; Takahashi, S.; Satoh, H.; Jaenicke, E.; Paulsen, H. Chlorophyll a/b Binding-Specificity in Water-Soluble Chlorophyll Protein. Nat. Plants 2018, 4, 920–929. [Google Scholar] [CrossRef]
- Carbonera, D.; Collareta, P.; Giacometti, G. The P700 Triplet State in an Intact Environment Detected by ODMR. A Well Resolved Triplet Minus Singlet Spectrum. Biochim. Biophys. Acta Bioenerg. 1997, 1322, 115–128. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a Comprehensive Software Package for Spectral Simulation and Analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Toffoletti, A.; Wang, Z.; Zhao, J.; Tommasini, M.; Barbon, A. Precise Determination of the Orientation of the Transition Dipole Moment in a Bodipy Derivative by Analysis of the Magnetophotoselection Effect. Phys. Chem. Chem. Phys. 2018, 20, 20497–20503. [Google Scholar] [CrossRef] [PubMed]
- López-Tarifa, P.; Liguori, N.; van den Heuvel, N.; Croce, R.; Visscher, L. Coulomb Couplings in Solubilised Light Harvesting Complex II (LHCII): Challenging the Ideal Dipole Approximation from TDDFT Calculations. Phys. Chem. Chem. Phys. 2017, 19, 18311–18320. [Google Scholar] [CrossRef]
- Frähmcke, J.S.; Walla, P.J. Coulombic Couplings between Pigments in the Major Light-Harvesting Complex LHC II Calculated by the Transition Density Cube Method. Chem. Phys. Lett. 2006, 430, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Parson, W.W. Modern Optical Spectroscopy, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Kay, C.W.M.; Di Valentin, M.; Möbius, K. A Time-Resolved Electron Nuclear Double Resonance (ENDOR) Study of the Photoexcited Triplet State of Free-Base Tetraphenylporphyrin. Sol. Energy Mater. Sol. Cells 1995, 38, 111–118. [Google Scholar] [CrossRef]
- Agostini, A.; Dal Farra, M.G.; Paulsen, H.; Polimeno, A.; Orian, L.; Di Valentin, M.; Carbonera, D. Similarity and Specificity of Chlorophyll b Triplet State in Comparison to Chlorophyll a as Revealed by EPR/ENDOR and DFT Calculations. J. Phys. Chem. B 2019, 123, 8232–8239. [Google Scholar] [CrossRef]
- Bowen, A.M.; Bertran, A.; Henbest, K.B.; Gobbo, M.; Timmel, C.R.; Di Valentin, M. Orientation-Selective and Frequency-Correlated Light-Induced Pulsed Dipolar Spectroscopy. J. Phys. Chem. Lett. 2021, 12, 3819–3826. [Google Scholar] [CrossRef] [PubMed]
- Niklas, J.; Agostini, A.; Carbonera, D.; Di Valentin, M.; Lubitz, W. Primary Donor Triplet States of Photosystem I and II Studied by Q-Band Pulse ENDOR Spectroscopy. Photosynth. Res. 2022. [Google Scholar] [CrossRef]
- Wehner, J.; Baumeier, B. Intermolecular Singlet and Triplet Exciton Transfer Integrals from Many-Body Green’s Functions Theory. J. Chem. Theory Comput. 2017, 13, 1584–1594. [Google Scholar] [CrossRef]
- Ohtsuki, T.; Uchida, A.; Nishigaki, A.; Nagashima, U.; Takahashi, S.; Ohshima, S. Excitonic Energy Level of Homo- and Hetero-Dimers and Their Composition in the Native Water-Soluble Chlorophyll Protein from Lepidium virginicum. Bull. Chem. Soc. Jpn. 2021, 94, 418–426. [Google Scholar] [CrossRef]
- Schmidt, K.; Fufezan, C.; Krieger-Liszkay, A.; Satoh, H.; Paulsen, H. Recombinant Water-Soluble Chlorophyll Protein from Brassica oleracea Var. Botrys Binds Various Chlorophyll Derivatives. Biochemistry 2003, 42, 7427–7433. [Google Scholar] [CrossRef]
- Girr, P.; Kilper, J.; Pohland, A.-C.; Paulsen, H. The Pigment Binding Behaviour of Water-Soluble Chlorophyll Protein (WSCP). Photochem. Photobiol. Sci. 2020, 19, 695–712. [Google Scholar] [CrossRef] [PubMed]
- Palm, D.M.; Agostini, A.; Tenzer, S.; Gloeckle, B.M.; Werwie, M.; Carbonera, D.; Paulsen, H. Water-Soluble Chlorophyll Protein (WSCP) Stably Binds Two or Four Chlorophylls. Biochemistry 2017, 56, 1726–1736. [Google Scholar] [CrossRef]
- Bednarczyk, D.; Tor-Cohen, C.; Das, P.K.; Noy, D. Direct Assembly in Aqueous Solutions of Stable Chlorophyllide Complexes with Type II Water-soluble Chlorophyll Proteins. Photochem. Photobiol. 2021, 97, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Bolzonello, L.; Albertini, M.; Collini, E.; Di Valentin, M. Delocalized Triplet State in Porphyrin J-Aggregates Revealed by EPR Spectroscopy. Phys. Chem. Chem. Phys. 2017, 19, 27173–27177. [Google Scholar] [CrossRef]
- Keijer, T.; Bouwens, T.; Hessels, J.; Reek, J.N.H. Supramolecular Strategies in Artificial Photosynthesis. Chem. Sci. 2021, 12, 50–70. [Google Scholar] [CrossRef] [PubMed]
- Weil, J.A.; Bolton, J.R. Electron Paramagnetic Resonance; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Gonen, O.; Levanon, H. Time-resolved EPR spectroscopy of electron spin polarized ZnTPP triplets oriented in a liquid crystal. J. Phys. Chem. 1985, 89, 1637–1643. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | |D| (mT) | |E| (mT) | px | py | pz | ωm (°) | φm (°) | cP (%) |
|---|---|---|---|---|---|---|---|---|
| MeTHF | 30.4 | 4.3 | 0.31 | 0.56 | 0.13 | 90 | 65 | 63 |
| EtOH:MeOH 3:2 | 29.5 | 3.8 | 0.31 | 0.57 | 0.12 | 90 | 54 | 64 |
| λ (nm) | |D| (mT) | |E| (mT) | px | py | pz | ωex (°) | φex (°) | cP (%) | W (%) |
|---|---|---|---|---|---|---|---|---|---|
| 658 | 30.9 | 3.65 | 0.29 | 0.59 | 0.12 | 76 | 30 | 57 | 100 |
| 675 | 30.9 | 3.65 | 0.29 | 0.59 | 0.12 | 52 | 72 | 57 | 64 |
| 30.9 | 3.65 | 0.29 | 0.59 | 0.12 | 76 | 30 | 57 | 36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciuti, S.; Agostini, A.; Barbon, A.; Bortolus, M.; Paulsen, H.; Di Valentin, M.; Carbonera, D. Magnetophotoselection in the Investigation of Excitonically Coupled Chromophores: The Case of the Water-Soluble Chlorophyll Protein. Molecules 2022, 27, 3654. https://doi.org/10.3390/molecules27123654
Ciuti S, Agostini A, Barbon A, Bortolus M, Paulsen H, Di Valentin M, Carbonera D. Magnetophotoselection in the Investigation of Excitonically Coupled Chromophores: The Case of the Water-Soluble Chlorophyll Protein. Molecules. 2022; 27(12):3654. https://doi.org/10.3390/molecules27123654
Chicago/Turabian StyleCiuti, Susanna, Alessandro Agostini, Antonio Barbon, Marco Bortolus, Harald Paulsen, Marilena Di Valentin, and Donatella Carbonera. 2022. "Magnetophotoselection in the Investigation of Excitonically Coupled Chromophores: The Case of the Water-Soluble Chlorophyll Protein" Molecules 27, no. 12: 3654. https://doi.org/10.3390/molecules27123654