

Epichloë Endophytes Shape the Foliar Endophytic Fungal Microbiome and Alter the Auxin and Salicylic Acid Phytohormone Levels in Two Meadow Fescue Cultivars

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Setup
2.2. Plant Performance Parameters
2.3. Sample Collection
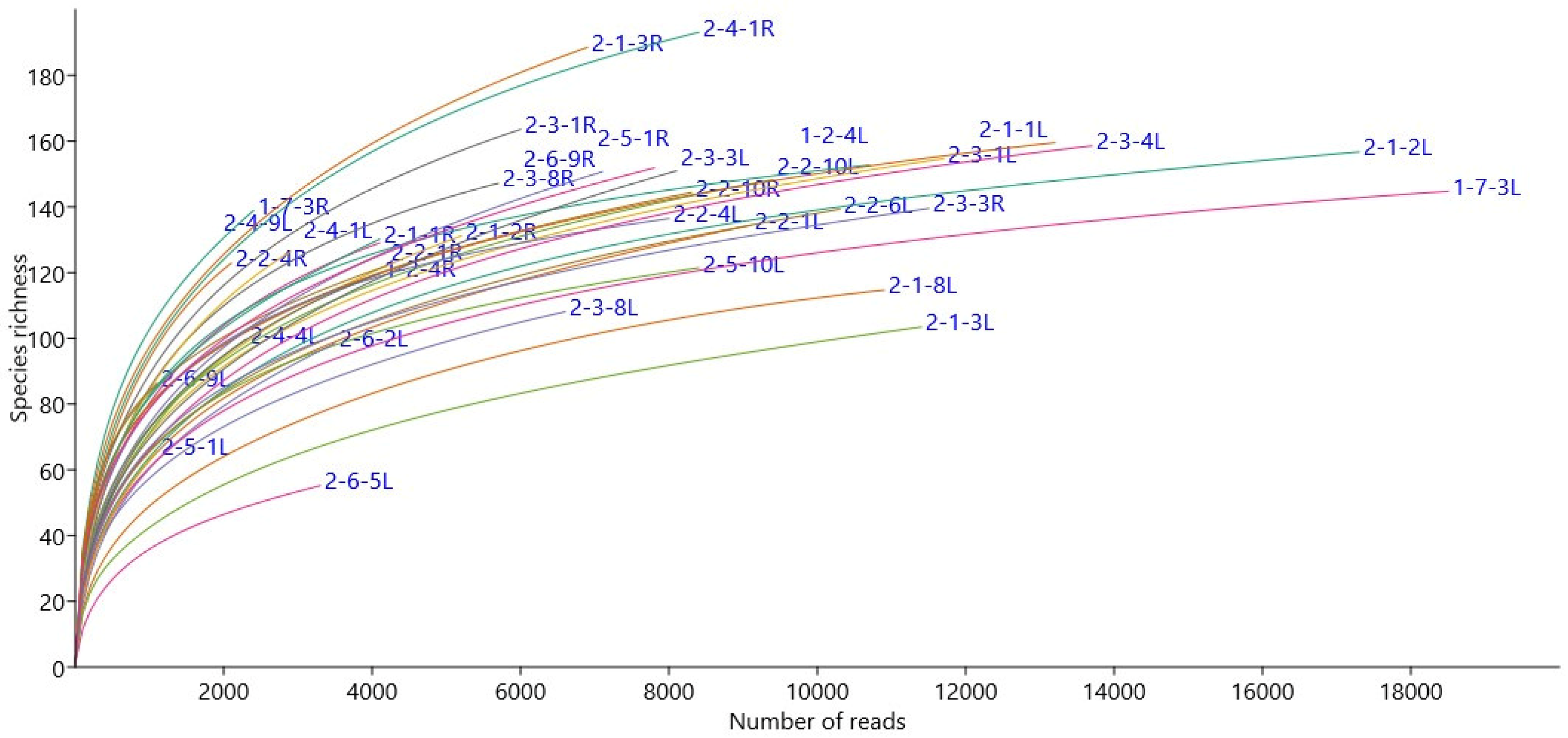
2.4. Microbiome Analysis
2.5. Plant Hormone Extraction and Quantification
2.6. Bioinformatics and Statistical Analys
3. Results
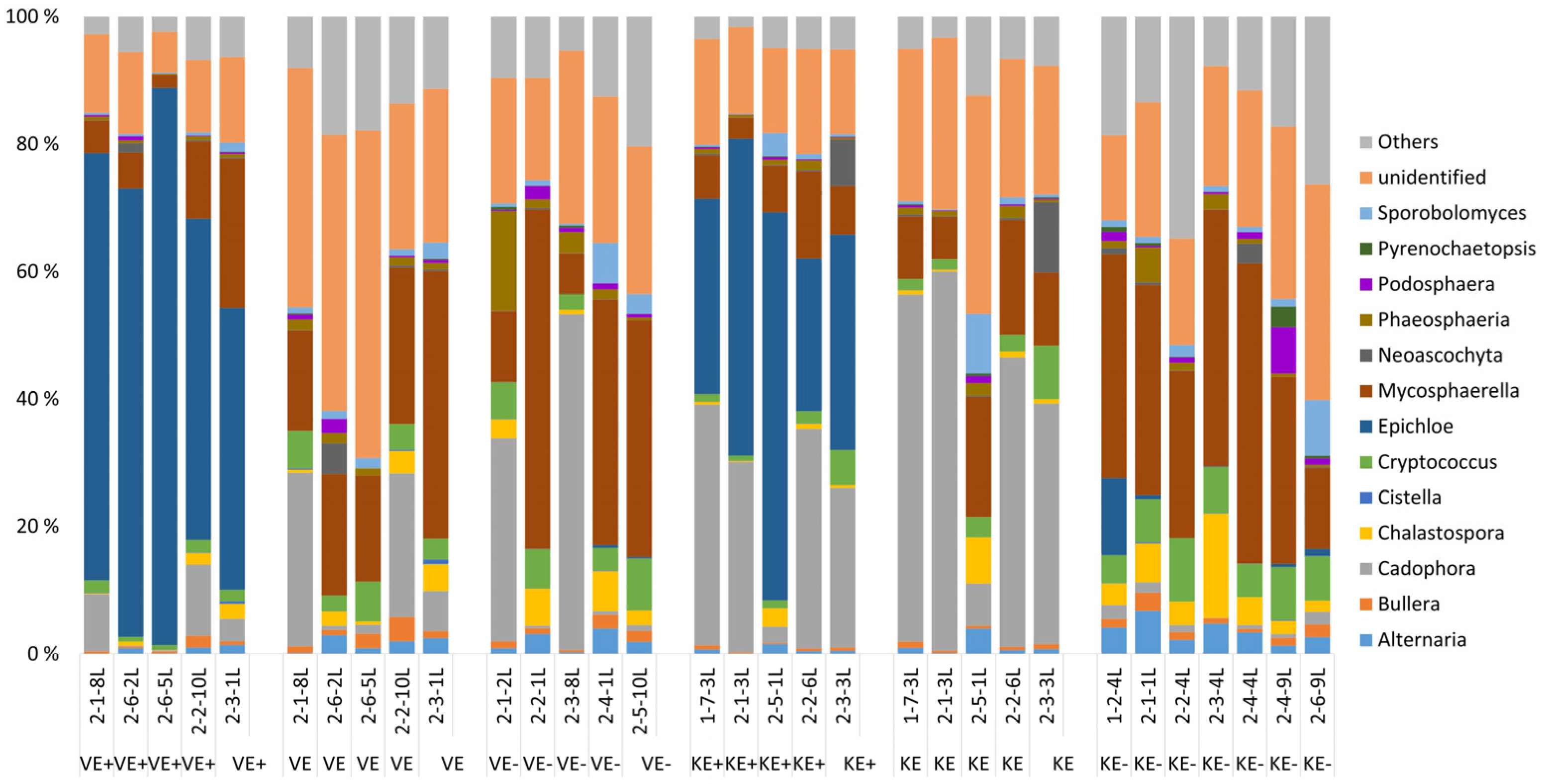
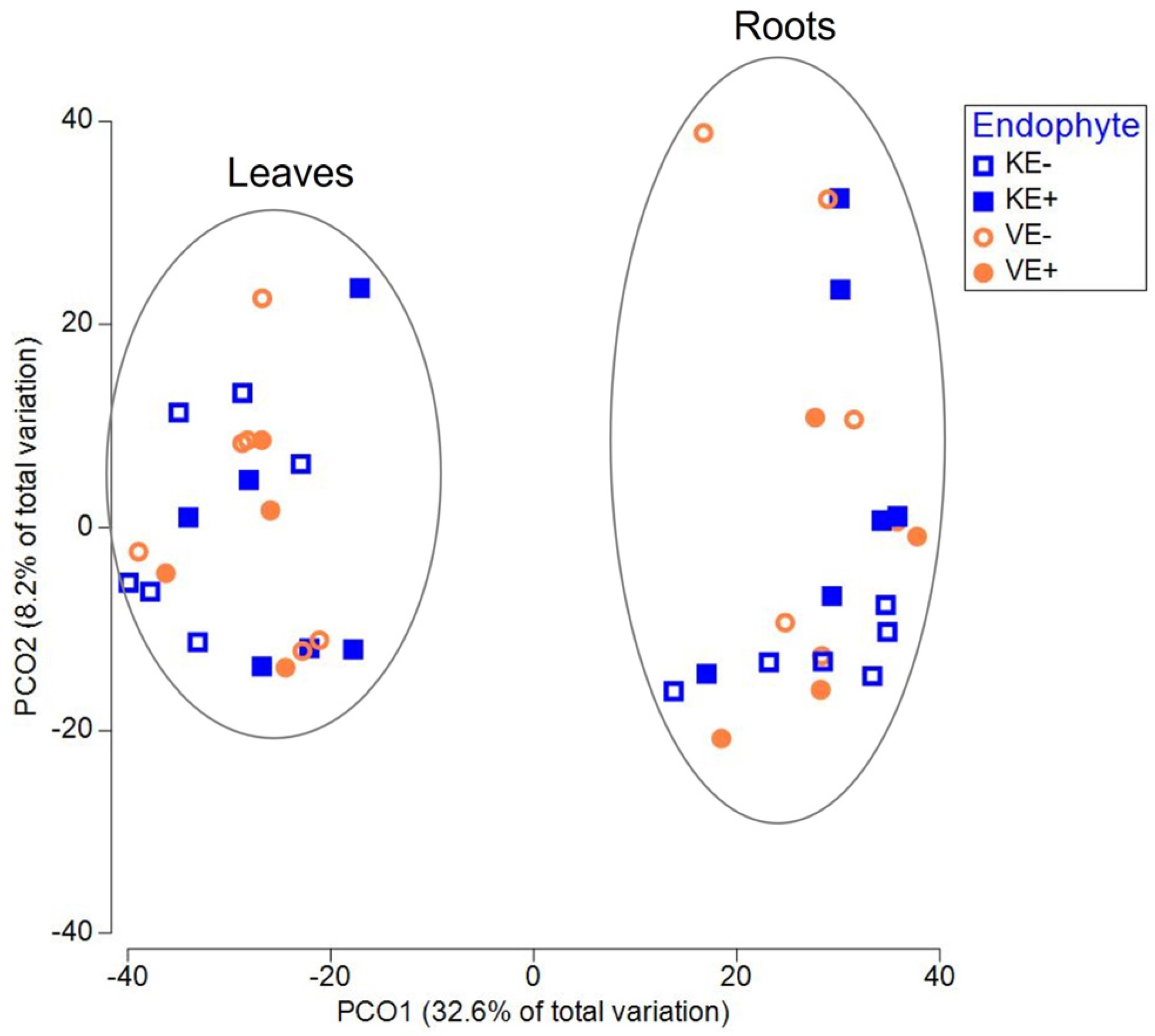
3.1. Epichloë Shapes Endophytic Fungal Community Composition in Leaves and Not in Roots
3.2. Epichloë Does Not Impact Endophytic Bacterial Community Structure
3.3. Epichloë Symbiosis Alters Plant Parameters
3.4. Epichloë Symbiosis Alters Plant Hormone Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kowalchuk, G.A.; Jones, S.E.; Blackall, L.L. Microbes Orchestrate Life on Earth. ISME J. 2008, 2, 795–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, J.A.; Neufeld, J.D. Life in a World without Microbes. PLoS Biol. 2014, 12, e1002020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemen, A.C.; Agler, M.T.; Kemen, E. Host–Microbe and Microbe–Microbe Interactions in the Evolution of Obligate Plant Parasitism. New Phytol. 2015, 206, 1207–1228. [Google Scholar] [CrossRef] [PubMed]
- Hassani, M.A.; Durán, P.; Hacquard, S. Microbial Interactions within the Plant Holobiont. Microbiome 2018, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Berg, G. Plant-Microbe Interactions Promoting Plant Growth and Health: Perspectives for Controlled Use of Microorganisms in Agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef]
- Elmer, G.W.; Mcfarland, L.V.; Mcfarland, M.; Russo, E.B. The Power of Probiotics: Improving Your Health with Beneficial Microbes; Routledge: Abingdon, UK, 2013; ISBN 978-1-317-82496-1. [Google Scholar]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–Microbiome Interactions under a Changing World: Responses, Consequences and Perspectives. New Phytol. 2022, 234, 1951–1959. [Google Scholar] [CrossRef]
- Saikkonen, K.; Nissinen, R.; Helander, M. Toward Comprehensive Plant Microbiome Research. Front. Ecol. Evol. 2020, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The Importance of the Microbiome of the Plant Holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Carvalhais, L.C.; Schenk, P.M.; Dennis, P.G. Jasmonic Acid Signalling and the Plant Holobiont. Curr. Opin. Microbiol. 2017, 37, 42–47. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Raaijmakers, J.M. Embracing Community Ecology in Plant Microbiome Research. Trends Plant Sci. 2018, 23, 467–469. [Google Scholar] [CrossRef]
- Sánchez Márquez, S.; Bills, G.F.; Herrero, N.; Zabalgogeazcoa, Í. Non-Systemic Fungal Endophytes of Grasses. Fungal Ecol. 2012, 5, 289–297. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Faeth, S.H.; Helander, M.; Sullivan, T.J. Fungal Endophytes: A Continuum of Interactions with Host Plants. Annu. Rev. Ecol. Syst. 1998, 29, 319–343. [Google Scholar] [CrossRef]
- Van Wees, S.C.M.; Van der Ent, S.; Pieterse, C.M.J. Plant Immune Responses Triggered by Beneficial Microbes. Curr. Opin. Plant Biol. 2008, 11, 443–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants 2022, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Jones, J.D.G.; Ding, P. Plant Immune Networks. Trends Plant Sci. 2022, 27, 255–273. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Gundel, P.E.; Helander, M.; Saikkonen, K. Endophytic Mediation of Reactive Oxygen Species and Antioxidant Activity in Plants: A Review. Fungal Divers. 2012, 54, 1–10. [Google Scholar] [CrossRef]
- Eichmann, R.; Richards, L.; Schäfer, P. Hormones as Go-Betweens in Plant Microbiome Assembly. Plant J. 2021, 105, 518–541. [Google Scholar] [CrossRef]
- Xu, G.; Yang, S.; Meng, L.; Wang, B.-G. The Plant Hormone Abscisic Acid Regulates the Growth and Metabolism of Endophytic Fungus Aspergillus nidulans. Sci. Rep. 2018, 8, 6504. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone Crosstalk in Plant Disease and Defense: More than Just Jasmonate-Salicylate Antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Foo, E.; Plett, J.M.; Lopez-Raez, J.A.; Reid, D. Editorial: The Role of Plant Hormones in Plant-Microbe Symbioses. Front. Plant Sci. 2019, 10, 1391. [Google Scholar] [CrossRef] [PubMed]
- Carvalhais, L.C.; Dennis, P.G.; Badri, D.V.; Tyson, G.W.; Vivanco, J.M.; Schenk, P.M. Activation of the Jasmonic Acid Plant Defence Pathway Alters the Composition of Rhizosphere Bacterial Communities. PLoS ONE 2013, 8, e56457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bürger, M.; Chory, J. Stressed out about hormones: How plants orchestrate immunity. Cell Host Microbe 2019, 26, 163–172. [Google Scholar] [CrossRef]
- Fitzpatrick, C.R.; Salas-González, I.; Conway, J.M.; Finkel, O.M.; Gilbert, S.; Russ, D.; Teixeira, P.J.P.L.; Dangl, J.L. The plant microbiome: From ecology to reductionism and beyond. Annu. Rev. Microbiol. 2020, 74, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Elkabetz, D.; Leibman-Markus, M.; Sayas, T.; Schneider, A.; Jami, E.; Kleiman, M.; Bar, M. Cytokinin drives assembly of the phyllosphere microbiome and promotes disease resistance through structural and chemical cues. ISME J. 2022, 16, 122–137. [Google Scholar] [CrossRef]
- Saikkonen, K.; Lehtonen, P.; Helander, M.; Koricheva, J.; Faeth, S.H. Model Systems in Ecology: Dissecting the Endophyte–Grass Literature. Trends Plant Sci. 2006, 11, 428–433. [Google Scholar] [CrossRef]
- Müller, C.B.; Krauss, J. Symbiosis between Grasses and Asexual Fungal Endophytes. Curr. Opin. Plant. Biol. 2005, 8, 450–456. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtmann, A.; Spiering, M.J. Symbioses of Grasses with Seedborne Fungal Endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef]
- Saikkonen, K.; Saari, S.; Helander, M. Defensive Mutualism between Plants and Endophytic Fungi? Fungal Divers. 2010, 41, 101–113. [Google Scholar] [CrossRef]
- Saikkonen, K.; Gundel, P.E.; Helander, M. Chemical Ecology Mediated by Fungal Endophytes in Grasses. J. Chem. Ecol. 2013, 39, 962–968. [Google Scholar] [CrossRef]
- Bastías, D.A.; Alejandra Martínez-Ghersa, M.; Newman, J.A.; Card, S.D.; Mace, W.J.; Gundel, P.E. The Plant Hormone Salicylic Acid Interacts with the Mechanism of Anti-Herbivory Conferred by Fungal Endophytes in Grasses. Plant Cell Environ. 2018, 41, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Herbivore-Specific Induction of Defence Metabolites in a Grass–Endophyte Association. Funct. Ecol. 2017, 31, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Laihonen, M.; Saikkonen, K.; Helander, M.; Vázquez de Aldana, B.R.; Zabalgogeazcoa, I.; Fuchs, B. Epichloë Endophyte-Promoted Seed Pathogen Increases Host Grass Resistance Against Insect Herbivory. Front. Microbiol. 2022, 12, 786619. [Google Scholar] [CrossRef]
- Pérez, L.I.; Gundel, P.E.; Ghersa, C.M.; Omacini, M. Family Issues: Fungal Endophyte Protects Host Grass from the Closely Related Pathogen Claviceps purpurea. Fungal Ecol. 2013, 6, 379–386. [Google Scholar] [CrossRef]
- Bastías, D.A.; Bustos, L.B.; Jáuregui, R.; Barrera, A.; Acuña-Rodríguez, I.S.; Molina-Montenegro, M.A.; Gundel, P.E. Epichloë Fungal Endophytes Influence Seed-Associated Bacterial Communities. Front. Microbiol. 2022, 12, 795354. [Google Scholar] [CrossRef]
- König, J.; Guerreiro, M.A.; Peršoh, D.; Begerow, D.; Krauss, J. Knowing Your Neighbourhood—The Effects of Epichloë Endophytes on Foliar Fungal Assemblages in Perennial Ryegrass in Dependence of Season and Land-Use Intensity. PeerJ 2018, 6, e4660. [Google Scholar] [CrossRef] [Green Version]
- Nissinen, R.; Helander, M.; Kumar, M.; Saikkonen, K. Heritable Epichloë Symbiosis Shapes Fungal but Not Bacterial Communities of Plant Leaves. Sci. Rep. 2019, 9, 5253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, J.C.M.; Newman, J.A. A First Draft of the Core Fungal Microbiome of Schedonorus arundinaceus with and without Its Fungal Mutualist Epichloë coenophiala. J. Fungi 2022, 8, 1026. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; Gundel, P.E.; Helander, M.; Saikkonen, K. Non-Systemic Fungal Endophytes in Festuca rubra Plants Infected by Epichloë festucae in Subarctic Habitats. Fungal Divers. 2013, 60, 25–32. [Google Scholar] [CrossRef]
- Ju, Y.; Zhong, R.; Christensen, M.J.; Zhang, X. Effects of Epichloë gansuensis Endophyte on the Root and Rhizosphere Soil Bacteria of Achnatherum inebrians Under Different Moisture Conditions. Front. Microbiol. 2020, 11, 747. [Google Scholar] [CrossRef]
- Liu, B.; Ju, Y.; Xia, C.; Zhong, R.; Christensen, M.J.; Zhang, X.; Nan, Z. The Effect of Epichloë Endophyte on Phyllosphere Microbes and Leaf Metabolites in Achnatherum inebrians. iScience 2022, 25, 104144. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.C.; Jackson, M.A.; Johnson-Cicalese, J.M. A Rapid Staining Method for Detection of Endophytic Fungi in Turf and Forage Grasses. Phytopathology 1988, 78, 237–239. [Google Scholar] [CrossRef]
- Chelius, M.K.; Triplett, E.W. The Diversity of Archaea and Bacteria in Association with the Roots of Zea mays L. Microb Ecol. 2001, 41, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Ghyselinck, J.; Pfeiffer, S.; Heylen, K.; Sessitsch, A.; De Vos, P. The Effect of Primer Choice and Short Read Sequences on the Outcome of 16S rRNA Gene Based Diversity Studies (J Ravel, Ed.). PLoS ONE 2013, 8, e71360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mäki, A.; Rissanen, A.J.; Tiirola, M. A practical method for barcoding and size-trimming PCR templates for amplicon sequencing. BioTechniques 2016, 60, 88–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, D.; Alm, E.W.; Stahl, D.A.; Raskin, L. Characterization of universal small-subunit rRNA hybridization probes for quantitative molecular microbial ecology studies. Appl. Environ. Microbiol. 1996, 62, 4504–4513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region - evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef]
- Dobrev, P.I.; Vankova, R. Quantification of Abscisic Acid, Cytokinin, and Auxin Content in Salt-Stressed Plant Tissues. In Plant Salt Tolerance: Methods and Protocols; Shabala, S., Cuin, T.A., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; pp. 251–261. ISBN 978-1-61779-986-0. [Google Scholar]
- Fuchs, B.; Laihonen, M.; Muola, A.; Saikkonen, K.; Dobrev, P.I.; Vankova, R.; Helander, M. A Glyphosate-Based Herbicide in Soil Differentially Affects Hormonal Homeostasis and Performance of Non-Target Crop Plants. Front. Plant Sci. 2022, 12, 787958. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal Endophytes: Diversity and Functional Roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Fuchs, B.; Kuhnert, E.; Krauss, J. Contrasting Effects of Grass–Endophyte Chemotypes on a Tri-Trophic Cascade. J. Chem. Ecol. 2020, 46, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikkonen, K.; Young, C.A.; Helander, M.; Schardl, C.L. Endophytic Epichloë Species and Their Grass Hosts: From Evolution to Applications. Plant Mol. Biol. 2016, 90, 665–675. [Google Scholar] [CrossRef] [Green Version]
- König, J.; Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Hide and Seek—Infection Rates and Alkaloid Concentrations of Epichloë festucae var. lolii in Lolium perenne along a Land-Use Gradient in Germany. Grass Forage Sci. 2018, 73, 510–516. [Google Scholar] [CrossRef]
- Fuchs, B.; Krischke, M.; Mueller, M.J.; Krauss, J. Plant Age and Seasonal Timing Determine Endophyte Growth and Alkaloid Biosynthesis. Fungal Ecol. 2017, 29, 52–58. [Google Scholar] [CrossRef]
- Vikuk, V.; Fuchs, B.; Krischke, M.; Mueller, M.J.; Rueb, S.; Krauss, J. Alkaloid Concentrations of Lolium perenne Infected with Epichloë festucae var. J. Fungi 2020, 6, 177. [Google Scholar] [CrossRef] [PubMed]
- Saari, S.; Lehtonen, P.; Helander, M.; Saikkonen, K. High Variation in Frequency of Infection by Endophytes in Cultivars of Meadow Fescue in Finland. Grass Forage Sci. 2009, 64, 169–176. [Google Scholar] [CrossRef]
- Wahdan, S.F.M.; Buscot, F.; Purahong, W. Future Climate Alters Pathogens-Microbiome Co-Occurrence Networks in Wheat Straw Residues during Decomposition. Proceedings 2021, 66, 22. [Google Scholar] [CrossRef]
- McGorum, B.C.; Chen, Z.; Glendinning, L.; Gweon, H.S.; Hunt, L.; Ivens, A.; Keen, J.A.; Pirie, R.S.; Taylor, J.; Wilkinson, T.; et al. Equine Grass Sickness (a Multiple Systems Neuropathy) Is Associated with Alterations in the Gastrointestinal Mycobiome. Anim. Microbiome 2021, 3, 70. [Google Scholar] [CrossRef]
- Pérez, L.I.; Gundel, P.E.; Zabalgogeazcoa, I.; Omacini, M. An Ecological Framework for Understanding the Roles of Epichloë Endophytes on Plant Defenses against Fungal Diseases. Fungal Biol. Rev. 2020, 34, 115–125. [Google Scholar] [CrossRef]
- Card, S.D.; Bastías, D.A.; Caradus, J.R. Antagonism to Plant Pathogens by Epichloë Fungal Endophytes—A Review. Plants 2021, 10, 1997. [Google Scholar] [CrossRef]
- Xia, C.; Li, N.; Zhang, Y.; Li, C.; Zhang, X.; Nan, Z. Role of Epichloë Endophytes in Defense Responses of Cool-Season Grasses to Pathogens: A Review. Plant Dis. 2018, 102, 2061–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vujanovic, V. Tremellomycetes Yeasts in Kernel Ecological Niche: Early Indicators of Enhanced Competitiveness of Endophytic and Mycoparasitic Symbionts against Wheat Pathobiota. Plants 2021, 10, 905. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.S.; White, J.F.; Zhang, X.; Hinton, D.M.; Bacon, C.W. Endophyte-Mediated Adjustments in Host Morphology and Physiology and Effects on Host Fitness Traits in Grasses. Fungal Ecol. 2012, 5, 322–330. [Google Scholar] [CrossRef]
- Kou, M.-Z.; Bastías, D.A.; Christensen, M.J.; Zhong, R.; Nan, Z.-B.; Zhang, X.-X. The Plant Salicylic Acid Signalling Pathway Regulates the Infection of a Biotrophic Pathogen in Grasses Associated with an Epichloë Endophyte. J. Fungi 2021, 7, 633. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, G.; Pandharikar, G.; Frendo, P. Salicylic Acid in Plant Symbioses: Beyond Plant Pathogen Interactions. Biology 2022, 11, 861. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | SS | MS | Pseudo-F | P(perm) | Unique Perms |
|---|---|---|---|---|---|---|
| Tissue | 1 | 33964 | 433964 | 30.86 | 0.001 | 998 |
| Endophyte | 3 | 9306.3 | 3102.1 | 22.82 | 0.001 | 997 |
| Tissue × Endophyte | 3 | 6968.3 | 2322.8 | 2.11 | 0.001 | 997 |
| PERMANOVA analysis on total fungal communities for leaves | ||||||
| Endophyte | 3 | 14915 | 4971.8 | 66.1988 | 0.001 | 998 |
| PERMANOVA analysis on total fungal communities for roots | ||||||
| Endophyte | 3 | 4130 | 1376.7 | 0.840 | 0.79 | 997 |
| Source | df | SS | MS | Pseudo-F | P(perm) | Unique Perms |
|---|---|---|---|---|---|---|
| Tissue | 1 | 34398 | 34398 | 27.47 | 0.001 | 998 |
| Endophyte | 3 | 7297.7 | 2432.6 | 1.9426 | 0.001 | 995 |
| Tissue × Endophyte | 3 | 5175.1 | 1725 | 1.3776 | 0.028 | 998 |
| Total fungal communities | |||
| VE- | VE+ | ||
| Av.Abund | Av.Abund | Taxa | |
| 0.02 | 59.43 | Epichloë | |
| 28.53 | 8.75 | Mycosphaerella_tassiana | |
| KE- | KE+ | ||
| Av.Abund | Av.Abund | ||
| 0.03 | 36.83 | Epichloë | |
| 30.87 | 7.06 | Mycosphaerella_tassiana | |
| 0.03 | 18.91 | Cadophora | |
| 0.01 | 7.82 | Heliotales_unidentified | |
| Epichloë-depleted communities | |||
| VE- | VE+ | ||
| Av.Abund | Av.Abund | Taxa | |
| 28.60 | 19.39 | Mycosphaerella_tassiana | |
| 12.11 | 7.81 | Cadophora | |
| 5.27 | 3.08 | Heliotales_sps. | |
| 5.93 | 7.75 | Pleosporales_sps. | |
| 4.25 | 0.72 | Phaeosphaeria triglochinicola | |
| 4.70 | 2.01 | Capnodiales sps. | |
| KE- | KE+ | ||
| Av.Abund | Av.Abund | ||
| 0.03 | 29.32 | Cadophora | |
| 31.6 | 11.49 | Mycosphaerella_tassiana | |
| 0.01 | 12.1 | Heliotales_sps. | |
| 6.48 | 4.06 | Pleosporales_sps. | |
| 7.36 | 1.83 | Vishniacozyma victoriae | |
| ‘Kasper’ | ‘Valtteri’ | |||||
|---|---|---|---|---|---|---|
| t | F | p-Value | t | F | p-Value | |
| Plant height | −3.172 | 9.939 | 0.01 | −2.937 | 7.793 | 0.02 |
| Longest leaf | −4.23 | 8.993 | 0.68 | −0.148 | 7.203 | 0.89 |
| Circumference | 2.474 | 9.019 | 0.04 | 1.961 | 5.119 | 0.11 |
| Tiller no. | −0.308 | 9.981 | 0.76 | −0.468 | 7.387 | 0.653 |
| Flower head no. | −2.895 | 6.592 | 0.02 | −2.539 | 7.859 | 0.04 |
| SPAD value | −1.982 | 9.951 | 0.08 | −2.144 | 6.163 | 0.07 |
| Phytohormone | ‘Kasper’ | ‘Valtteri’ | ||
|---|---|---|---|---|
| F | p-Value | F | p-Value | |
| ABA | 3.32 | 0.04 | 0.21 | 0.84 |
| PA | 0.07 | 0.95 | 1.06 | 0.32 |
| IAA | 4.2 | <0.01 | 3.68 | <0.01 |
| PAA | 2.52 | 0.03 | 3.92 | <0.01 |
| SA | 2.43 | 0.05 | 3.12 | 0.01 |
| BzA | 1.18 | 0.27 | 1.37 | 0.21 |
| JA | 0.78 | 0.45 | 0.35 | 0.74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathew, S.A.; Helander, M.; Saikkonen, K.; Vankova, R.; Dobrev, P.I.; Dirihan, S.; Fuchs, B. Epichloë Endophytes Shape the Foliar Endophytic Fungal Microbiome and Alter the Auxin and Salicylic Acid Phytohormone Levels in Two Meadow Fescue Cultivars. J. Fungi 2023, 9, 90. https://doi.org/10.3390/jof9010090
Mathew SA, Helander M, Saikkonen K, Vankova R, Dobrev PI, Dirihan S, Fuchs B. Epichloë Endophytes Shape the Foliar Endophytic Fungal Microbiome and Alter the Auxin and Salicylic Acid Phytohormone Levels in Two Meadow Fescue Cultivars. Journal of Fungi. 2023; 9(1):90. https://doi.org/10.3390/jof9010090
Chicago/Turabian StyleMathew, Suni Anie, Marjo Helander, Kari Saikkonen, Radomira Vankova, Petre I. Dobrev, Serdar Dirihan, and Benjamin Fuchs. 2023. "Epichloë Endophytes Shape the Foliar Endophytic Fungal Microbiome and Alter the Auxin and Salicylic Acid Phytohormone Levels in Two Meadow Fescue Cultivars" Journal of Fungi 9, no. 1: 90. https://doi.org/10.3390/jof9010090