
An Unusual Two-Domain Thyropin from Tick Saliva: NMR Solution Structure and Highly Selective Inhibition of Cysteine Cathepsins Modulated by Glycosaminoglycans

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. IrThy—A Salivary Protein Secreted by the I. ricinus Tick
2.2. IrThy Is a Two-Domain Thyropin: Sequence Analysis, Evolutionary Distribution, and Recombinant Production
2.3. The Unique, Narrow Inhibitory Specificity of IrThy
2.4. IrThy Is Prone to Proteolytic Degradation by Target Proteases at High Concentrations
2.5. Complex Glycosaminoglycan Modulation of IrThy Inhibitory Activity
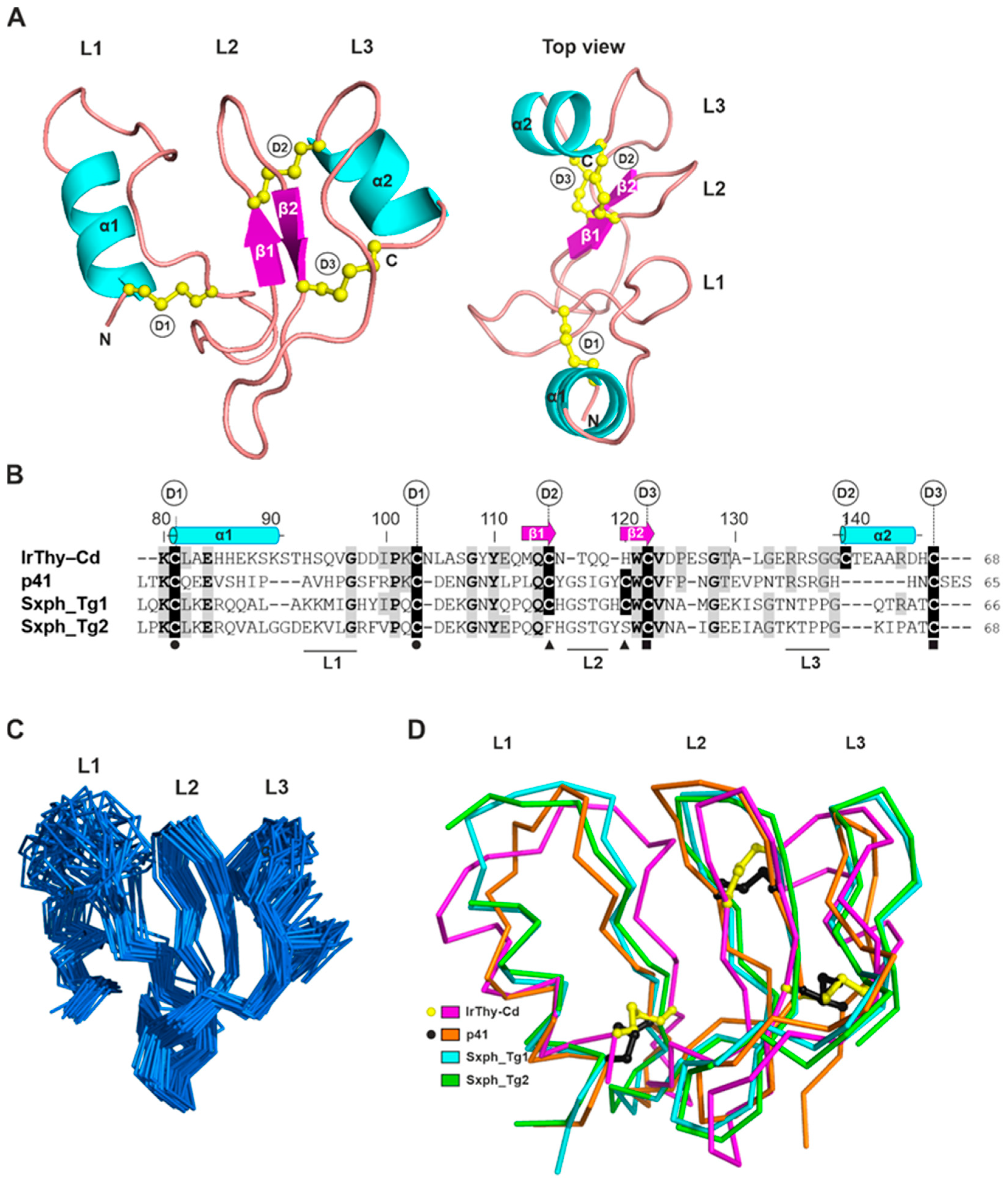
2.6. Spatial Structure of the IrThy C-Domain Determined by NMR Spectroscopy
3. Discussion
4. Materials and Methods
4.1. Materials
Ticks and Tick-Derived Materials
4.2. IrThy Cloning and Sequencing
4.3. Production of Recombinant IrThy and its Domains in Insect Cells
4.4. Production of Recombinant IrThy and its Domains in E. coli
4.5. Expression Analysis by Quantitative Real-Time PCR
4.6. Mass Spectrometry Proteomic Analysis
4.7. Phylogenetic Distribution Analysis
4.8. Protease Inhibition Assays
4.9. Proteolytic Degradation of IrThy
4.10. NMR Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Molina, F.; Bouanani, M.; Pau, B.; Granier, C. Characterization of the type-1 repeat from thyroglobulin, a cysteine-rich module found in proteins from different families. Eur. J. Biochem. 1996, 240, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Lenarcic, B.; Bevec, T. Thyropins—New structurally related proteinase inhibitors. Biol. Chem. 1998, 379, 105–111. [Google Scholar]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2017, 46, D624–D632. [Google Scholar] [CrossRef]
- Mihelic, M.; Turk, D. Two decades of thyroglobulin type-1 domain research. Biol. Chem. 2007, 388, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Mihelic, M.; Dobersek, A.; Guncar, G.; Turk, D. Inhibitory fragment from the p41 form of invariant chain can regulate activity of cysteine cathepsins in antigen presentation. J. Biol. Chem. 2008, 283, 14453–14460. [Google Scholar] [CrossRef]
- Guncar, G.; Pungercic, G.; Klemencic, I.; Turk, V.; Turk, D. Crystal structure of MHC class II-associated p41 Ii fragment bound to cathepsin L reveals the structural basis for differentiation between cathepsins L and S. EMBO J. 1999, 18, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Lenarcic, B.; Krishnan, G.; Borukhovich, R.; Ruck, B.; Turk, V.; Moczydlowski, E. Saxiphilin, a saxitoxin-binding protein with two thyroglobulin type 1 domains, is an inhibitor of papain-like cysteine proteinases. J. Biol. Chem. 2000, 275, 15572–15577. [Google Scholar] [CrossRef]
- Yen, T.J.; Lolicato, M.; Thomas-Tran, R.; Du Bois, J.; Minor, D.L. Structure of the saxiphilin: Saxitoxin (STX) complex reveals a convergent molecular recognition strategy for paralytic toxins. Sci. Adv. 2019, 5, eaax2650. [Google Scholar] [CrossRef]
- Lenarcic, B.; Turk, V. Thyroglobulin type-1 domains in equistatin inhibit both papain-like cysteine proteinases and cathepsin D. J. Biol. Chem. 1999, 274, 563–566. [Google Scholar] [CrossRef]
- Galesa, K.; Pain, R.; Jongsma, M.A.; Turk, V.; Lenarcic, B. Structural characterization of thyroglobulin type-1 domains of equistatin. FEBS Lett. 2003, 539, 120–124. [Google Scholar] [CrossRef]
- Bocock, J.P.; Edgell, C.J.; Marr, H.S.; Erickson, A.H. Human proteoglycan testican-1 inhibits the lysosomal cysteine protease cathepsin L. Eur. J. Biochem. 2003, 270, 4008–4015. [Google Scholar] [CrossRef] [PubMed]
- Meh, P.; Pavsic, M.; Turk, V.; Baici, A.; Lenarcic, B. Dual concentration-dependent activity of thyroglobulin type-1 domain of testican: Specific inhibitor and substrate of cathepsin L. Biol. Chem. 2005, 386, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Sankpal, N.V.; Brown, T.C.; Fleming, T.P.; Herndon, J.M.; Amaravati, A.A.; Loynd, A.N.; Gillanders, W.E. Cancer-associated mutations reveal a novel role for EpCAM as an inhibitor of cathepsin-L and tumor cell invasion. BMC Cancer 2021, 21, 541. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Konagaya, S. A novel cysteine protease inhibitor of the egg of chum salmon, containing a cysteine-rich thyroglobulin-like motif. J. Biol. Chem. 1996, 271, 1282–1284. [Google Scholar] [CrossRef] [PubMed]
- Fowlkes, J.L.; Thrailkill, K.M.; Serra, D.M.; Nagase, H. Insulin-like growth factor binding protein (IGFBP) substrate zymography. A new tool to identify and characterize IGFBP-degrading proteinases. Endocrine 1997, 7, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.J.; Ball, M.; Rukhlova, M.; Slinn, J.; L’Abbe, D.; Iqbal, U.; Monette, R.; Hagedorn, M.; O’Connor-McCourt, M.D.; Durocher, Y.; et al. IGFBP-4 anti-angiogenic and anti-tumorigenic effects are associated with anti-cathepsin B activity. Neoplasia 2013, 15, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Fiebiger, E.; Maehr, R.; Villadangos, J.; Weber, E.; Erickson, A.; Bikoff, E.; Ploegh, H.L.; Lennon-Duménil, A.M. Invariant chain controls the activity of extracellular cathepsin L. J. Exp. Med. 2002, 196, 1263–1269. [Google Scholar] [CrossRef]
- Porter, L.M.; Radulović, Ž.M.; Mulenga, A. A repertoire of protease inhibitor families in Amblyomma americanum and other tick species: Inter-species comparative analyses. Parasit. Vectors 2017, 10, 152. [Google Scholar] [CrossRef]
- Oliveira, C.J.; Anatriello, E.; de Miranda-Santos, I.K.; Francischetti, I.M.; Sá-Nunes, A.; Ferreira, B.R.; Ribeiro, J.M. Proteome of Rhipicephalus sanguineus tick saliva induced by the secretagogues pilocarpine and dopamine. Ticks Tick Borne Dis. 2013, 4, 469–477. [Google Scholar] [CrossRef]
- Bensaoud, C.; Tenzer, S.; Poplawski, A.; Medina, J.M.; Jmel, M.A.; Voet, H.; Mekki, I.; Aparicio-Puerta, E.; Cuveele, B.; Distler, U.; et al. Quantitative proteomics analysis reveals core and variable tick salivary proteins at the tick-vertebrate host interface. Mol. Ecol. 2022, 31, 4162–4175. [Google Scholar] [CrossRef]
- Kozelková, T.; Dyčka, F.; Lu, S.; Urbanová, V.; Frantová, H.; Sojka, D.; Šíma, R.; Horn, M.; Perner, J.; Kopáček, P. Insight Into the Dynamics of the Ixodes ricinus Nymphal Midgut Proteome. Mol. Cell Proteom. 2023, 22, 100663. [Google Scholar] [CrossRef]
- Kotál, J.; Buša, M.; Urbanová, V.; Řezáčová, P.; Chmelař, J.; Langhansová, H.; Sojka, D.; Mareš, M.; Kotsyfakis, M. Mialostatin, a Novel Midgut Cystatin from Ixodes ricinus Ticks: Crystal Structure and Regulation of Host Blood Digestion. Int. J. Mol. Sci. 2021, 22, 5371. [Google Scholar] [CrossRef]
- Kotál, J.; Stergiou, N.; Buša, M.; Chlastáková, A.; Beránková, Z.; Řezáčová, P.; Langhansová, H.; Schwarz, A.; Calvo, E.; Kopecký, J.; et al. The structure and function of Iristatin, a novel immunosuppressive tick salivary cystatin. Cell Mol. Life Sci. 2019, 76, 2003–2013. [Google Scholar] [CrossRef]
- Martins, L.A.; Buša, M.; Chlastáková, A.; Kotál, J.; Beránková, Z.; Stergiou, N.; Jmel, M.A.; Schmitt, E.; Chmelař, J.; Mareš, M.; et al. Protease-bound structure of Ricistatin provides insights into the mechanism of action of tick salivary cystatins in the vertebrate host. Cell Mol. Life Sci. 2023, 80, 339. [Google Scholar] [CrossRef]
- Kotsyfakis, M.; Sá-Nunes, A.; Francischetti, I.M.; Mather, T.N.; Andersen, J.F.; Ribeiro, J.M. Antiinflammatory and immunosuppressive activity of sialostatin L, a salivary cystatin from the tick Ixodes scapularis. J. Biol. Chem. 2006, 281, 26298–26307. [Google Scholar] [CrossRef]
- Schwarz, A.; von Reumont, B.M.; Erhart, J.; Chagas, A.C.; Ribeiro, J.M.; Kotsyfakis, M. De novo Ixodes ricinus salivary gland transcriptome analysis using two next-generation sequencing methodologies. FASEB J. 2013, 27, 4745–4756. [Google Scholar] [CrossRef]
- Perner, J.; Provazník, J.; Schrenková, J.; Urbanová, V.; Ribeiro, J.M.; Kopáček, P. RNA-seq analyses of the midgut from blood- and serum-fed Ixodes ricinus ticks. Sci. Rep. 2016, 6, 36695. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2022, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Pungercic, G.; Dolenc, I.; Dolinar, M.; Bevec, T.; Jenko, S.; Kolaric, S.; Turk, V. Individual recombinant thyroglobulin type-1 domains are substrates for lysosomal cysteine proteinases. Biol. Chem. 2002, 383, 1809–1812. [Google Scholar] [CrossRef] [PubMed]
- Denamur, S.; Chazeirat, T.; Maszota-Zieleniak, M.; Vivès, R.R.; Saidi, A.; Zhang, F.; Linhardt, R.J.; Labarthe, F.; Samsonov, S.A.; Lalmanach, G.; et al. Binding of heparan sulfate to human cystatin C modulates inhibition of cathepsin L: Putative consequences in mucopolysaccharidosis. Carbohydr. Polym. 2022, 293, 119734. [Google Scholar] [CrossRef]
- Rein, C.M.; Desai, U.R.; Church, F.C. Serpin-glycosaminoglycan interactions. Methods Enzymol. 2011, 501, 105–137. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kienetz, M.; Cherney, M.M.; James, M.N.G.; Brömme, D. The Crystal and Molecular Structures of a Cathepsin K: Chondroitin Sulfate Complex. J. Mol. Biol. 2008, 383, 78–91. [Google Scholar] [CrossRef] [PubMed]
- David, A.; Chazeirat, T.; Saidi, A.; Lalmanach, G.; Lecaille, F. The Interplay of Glycosaminoglycans and Cysteine Cathepsins in Mucopolysaccharidosis. Biomedicines 2023, 11, 810. [Google Scholar] [CrossRef] [PubMed]
- Chazeirat, T.; Denamur, S.; Bojarski, K.K.; Andrault, P.M.; Sizaret, D.; Zhang, F.; Saidi, A.; Tardieu, M.; Linhardt, R.J.; Labarthe, F.; et al. The abnormal accumulation of heparan sulfate in patients with mucopolysaccharidosis prevents the elastolytic activity of cathepsin V. Carbohydr. Polym. 2021, 253, 117261. [Google Scholar] [CrossRef] [PubMed]
- Aguda, A.H.; Panwar, P.; Du, X.; Nguyen, N.T.; Brayer, G.D.; Brömme, D. Structural basis of collagen fiber degradation by cathepsin K. Proc. Natl. Acad. Sci. USA 2014, 111, 17474–17479. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Bevec, T.; Stoka, V.; Pungercic, G.; Dolenc, I.; Turk, V. Major histocompatibility complex class II-associated p41 invariant chain fragment is a strong inhibitor of lysosomal cathepsin L. J. Exp. Med. 1996, 183, 1331–1338. [Google Scholar] [CrossRef]
- Lenarcic, B.; Ritonja, A.; Strukelj, B.; Turk, B.; Turk, V. Equistatin, a new inhibitor of cysteine proteinases from Actinia equina, is structurally related to thyroglobulin type-1 domain. J. Biol. Chem. 1997, 272, 13899–13903. [Google Scholar] [CrossRef]
- Musil, D.; Zucic, D.; Turk, D.; Engh, R.A.; Mayr, I.; Huber, R.; Popovic, T.; Turk, V.; Towatari, T.; Katunuma, N.; et al. The refined 2.15 A X-ray crystal structure of human liver cathepsin B: The structural basis for its specificity. EMBO J. 1991, 10, 2321–2330. [Google Scholar] [CrossRef]
- Guncar, G.; Podobnik, M.; Pungercar, J.; Strukelj, B.; Turk, V.; Turk, D. Crystal structure of porcine cathepsin H determined at 2.1 A resolution: Location of the mini-chain C-terminal carboxyl group defines cathepsin H aminopeptidase function. Structure 1998, 6, 51–61. [Google Scholar] [CrossRef]
- Horn, M.; Jílková, A.; Vondrášek, J.; Marešová, L.; Caffrey, C.R.; Mareš, M. Mapping the pro-peptide of the Schistosoma mansoni cathepsin B1 drug target: Modulation of inhibition by heparin and design of mimetic inhibitors. ACS Chem. Biol. 2011, 6, 609–617. [Google Scholar] [CrossRef]
- Jílková, A.; Horn, M.; Řezáčová, P.; Marešová, L.; Fajtová, P.; Brynda, J.; Vondrášek, J.; McKerrow, J.H.; Caffrey, C.R.; Mareš, M. Activation route of the Schistosoma mansoni cathepsin B1 drug target: Structural map with a glycosaminoglycan switch. Structure 2014, 22, 1786–1798. [Google Scholar] [CrossRef]
- Caglic, D.; Pungercar, J.R.; Pejler, G.; Turk, V.; Turk, B. Glycosaminoglycans facilitate procathepsin B activation through disruption of propeptide-mature enzyme interactions. J. Biol. Chem. 2007, 282, 33076–33085. [Google Scholar] [CrossRef]
- Almeida, P.C.; Nantes, I.L.; Chagas, J.R.; Rizzi, C.C.; Faljoni-Alario, A.; Carmona, E.; Juliano, L.; Nader, H.B.; Tersariol, I.L. Cathepsin B activity regulation. Heparin-like glycosaminogylcans protect human cathepsin B from alkaline pH-induced inactivation. J. Biol. Chem. 2001, 276, 944–951. [Google Scholar] [CrossRef]
- Novinec, M.; Lenarčič, B.; Turk, B. Cysteine cathepsin activity regulation by glycosaminoglycans. Biomed. Res. Int. 2014, 2014, 309718. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, Y.; Li, Z.; Greenbaum, D.; Bogyo, M.; Weber, E.; Brömme, D. Cathepsin V, a Novel and Potent Elastolytic Activity Expressed in Activated Macrophages. J. Biol. Chem. 2004, 279, 36761–36770. [Google Scholar] [CrossRef] [PubMed]
- Jmel, M.A.; Aounallah, H.; Bensaoud, C.; Mekki, I.; Chmelař, J.; Faria, F.; M’Ghirbi, Y.; Kotsyfakis, M. Insights into the Role of Tick Salivary Protease Inhibitors during Ectoparasite-Host Crosstalk. Int. J. Mol. Sci. 2021, 22, 892. [Google Scholar] [CrossRef] [PubMed]
- Panwar, P.; Hedtke, T.; Heinz, A.; Andrault, P.M.; Hoehenwarter, W.; Granville, D.J.; Schmelzer, C.E.H.; Brömme, D. Expression of elastolytic cathepsins in human skin and their involvement in age-dependent elastin degradation. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129544. [Google Scholar] [CrossRef] [PubMed]
- Dennemärker, J.; Lohmüller, T.; Mayerle, J.; Tacke, M.; Lerch, M.M.; Coussens, L.M.; Peters, C.; Reinheckel, T. Deficiency for the cysteine protease cathepsin L promotes tumor progression in mouse epidermis. Oncogene 2010, 29, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- Lecaille, F.; Chazeirat, T.; Saidi, A.; Lalmanach, G. Cathepsin V: Molecular characteristics and significance in health and disease. Mol. Aspects Med. 2022, 88, 101086. [Google Scholar] [CrossRef] [PubMed]
- Brömme, D.; Wilson, S. Role of Cysteine Cathepsins in Extracellular Proteolysis. In Extracellular Matrix Degradation; Parks, W.C., Mecham, R.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 23–51. [Google Scholar]
- Vizovišek, M.; Fonović, M.; Turk, B. Cysteine cathepsins in extracellular matrix remodeling: Extracellular matrix degradation and beyond. Matrix Biol. 2019, 75–76, 141–159. [Google Scholar] [CrossRef]
- Senjor, E.; Kos, J.; Nanut, M.P. Cysteine Cathepsins as Therapeutic Targets in Immune Regulation and Immune Disorders. Biomedicines 2023, 11, 476. [Google Scholar] [CrossRef] [PubMed]
- Khamtorn, P.; Peigneur, S.; Amorim, F.G.; Quinton, L.; Tytgat, J.; Daduang, S. De Novo Transcriptome Analysis of the Venom of Latrodectus geometricus with the Discovery of an Insect-Selective Na Channel Modulator. Molecules 2021, 27, 47. [Google Scholar] [CrossRef]
- Emerich, B.L.; Ferreira, R.C.M.; Cordeiro, M.N.; Borges, M.H.; Pimenta, A.M.C.; Figueiredo, S.G.; Duarte, I.D.G.; De Lima, M.E. δ-Ctenitoxin-Pn1a, a Peptide from Phoneutria nigriventer Spider Venom, Shows Antinociceptive Effect Involving Opioid and Cannabinoid Systems, in Rats. Toxins 2016, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Benýšek, J.; Buša, M.; Rubešová, P.; Fanfrlík, J.; Lepšík, M.; Brynda, J.; Matoušková, Z.; Bartz, U.; Horn, M.; Gütschow, M.; et al. Highly potent inhibitors of cathepsin K with a differently positioned cyanohydrazide warhead: Structural analysis of binding mode to mature and zymogen-like enzymes. J. Enzyme Inhib. Med. Chem. 2022, 37, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Brömme, D.; Li, Z.; Barnes, M.; Mehler, E. Human cathepsin V functional expression, tissue distribution, electrostatic surface potential, enzymatic characterization, and chromosomal localization. Biochemistry 1999, 38, 2377–2385. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Baudyš, M.; Voburka, Z.; Kluh, I.; Vondrášek, J.; Mareš, M. Free-thiol Cys331 exposed during activation process is critical for native tetramer structure of cathepsin C (dipeptidyl peptidase I). Protein Sci. 2002, 11, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Dolečková-Marešová, L.; Rulíšek, L.; Máša, M.; Vasiljeva, O.; Turk, B.; Gan-Erdene, T.; Baudys, M.; Mareš, M. Activation processing of cathepsin H impairs recognition by its propeptide. Biol. Chem. 2005, 386, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Máša, M.; Marešová, L.; Vondrášek, J.; Horn, M.; Ježek, J.; Mareš, M. Cathepsin D propeptide: Mechanism and regulation of its interaction with the catalytic core. Biochemistry 2006, 45, 15474–15482. [Google Scholar] [CrossRef]
- Perner, J.; Helm, D.; Haberkant, P.; Hatalová, T.; Kropáčková, S.; Ribeiro, J.M.; Kopáček, P. The Central Role of Salivary Metalloproteases in Host Acquired Resistance to Tick Feeding. Front. Cell Infect. Microbiol. 2020, 10, 563349. [Google Scholar] [CrossRef]
- Urbanová, V.; Šíma, R.; Šauman, I.; Hajdušek, O.; Kopáček, P. Thioester-containing proteins of the tick Ixodes ricinus: Gene expression, response to microbial challenge and their role in phagocytosis of the yeast Candida albicans. Dev. Comp. Immunol. 2015, 48, 55–64. [Google Scholar] [CrossRef]
- Perner, J.; Kropáčková, S.; Kopáček, P.; Ribeiro, J.M.C. Sialome diversity of ticks revealed by RNAseq of single tick salivary glands. PLoS Negl. Trop. Dis. 2018, 12, e0006410. [Google Scholar] [CrossRef]
- Jílková, A.; Řezáčová, P.; Lepšík, M.; Horn, M.; Váchová, J.; Fanfrlík, J.; Brynda, J.; McKerrow, J.H.; Caffrey, C.R.; Mareš, M. Structural basis for inhibition of cathepsin B drug target from the human blood fluke, Schistosoma mansoni. J. Biol. Chem. 2011, 286, 35770–35781. [Google Scholar] [CrossRef]
- Qi, D.; Scholthof, K.B. A one-step PCR-based method for rapid and efficient site-directed fragment deletion, insertion, and substitution mutagenesis. J. Virol. Methods 2008, 149, 85–90. [Google Scholar] [CrossRef]
- Sojka, D.; Franta, Z.; Frantová, H.; Bartošová, P.; Horn, M.; Váchová, J.; O’Donoghue, A.J.; Eroy-Reveles, A.A.; Craik, C.S.; Knudsen, G.M.; et al. Characterization of gut-associated cathepsin D hemoglobinase from tick Ixodes ricinus (IrCD1). J. Biol. Chem. 2012, 287, 21152–21163. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Fojtík, L.; Fiala, J.; Pompach, P.; Chmelík, J.; Matoušek, V.; Beier, P.; Kukačka, Z.; Novák, P. Fast Fluoroalkylation of Proteins Uncovers the Structure and Dynamics of Biological Macromolecules. J. Am. Chem. Soc. 2021, 143, 20670–20679. [Google Scholar] [CrossRef] [PubMed]
- Renshaw, P.S.; Lightbody, K.L.; Veverka, V.; Muskett, F.W.; Kelly, G.; Frenkiel, T.A.; Gordon, S.V.; Hewinson, R.G.; Burke, B.; Norman, J.; et al. Structure and function of the complex formed by the tuberculosis virulence factors CFP-10 and ESAT-6. EMBO J. 2005, 24, 2491–2498. [Google Scholar] [CrossRef] [PubMed]
- Veverka, V.; Lennie, G.; Crabbe, T.; Bird, I.; Taylor, R.J.; Carr, M.D. NMR assignment of the mTOR domain responsible for rapamycin binding. J. Biomol. NMR 2006, 36 (Suppl. 1), 3. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced software for biomolecular NMR spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, T.; Güntert, P.; Wüthrich, K. Protein NMR structure determination with automated NOE assignment using the new software CANDID and the torsion angle dynamics algorithm DYANA. J. Mol. Biol. 2002, 319, 209–227. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Delaglio, F.; Cornilescu, G.; Bax, A. TALOS+: A hybrid method for predicting protein backbone torsion angles from NMR chemical shifts. J. Biomol. NMR 2009, 44, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Janson, G.; Zhang, C.; Prado, M.G.; Paiardini, A. PyMod 2.0: Improvements in protein sequence-structure analysis and homology modeling within PyMOL. Bioinformatics 2017, 33, 444–446. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef]
- Meng, E.C.; Goddard, T.D.; Pettersen, E.F.; Couch, G.S.; Pearson, Z.J.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 2023, 32, e4792. [Google Scholar] [CrossRef]
- Kukačka, Z.; Rosůlek, M.; Jelínek, J.; Slavata, L.; Kavan, D.; Novák, P. LinX: A Software Tool for Uncommon Cross-Linking Chemistry. J. Proteome Res. 2021, 20, 2021–2027. [Google Scholar] [CrossRef]
- Götze, M.; Pettelkau, J.; Fritzsche, R.; Ihling, C.H.; Schäfer, M.; Sinz, A. Automated assignment of MS/MS cleavable cross-links in protein 3D-structure analysis. J. Am. Soc. Mass Spectrom. 2015, 26, 83–97. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Enzyme Specificity, Protease Class/Clan | Ki (nM) | ||
|---|---|---|---|---|
| IrThy | IrThy-Nd | IrThy-Cd | ||
| Host cathepsin proteases | ||||
| Cathepsin V | Endopeptidase, Cys/CA | 34.7 ± 2.9 | 27.2 ± 3.1 | 40.7 ± 3.8 |
| Cathepsin K | Endopeptidase, Cys/CA | 56.9 ± 4.3 | 43.7 ± 5.4 | 53.1 ± 7.7 |
| Cathepsin L | Endopeptidase, Cys/CA | 201.4 ± 15.8 | 178.6 ± 6.7 | 795.1 ± 19.8 |
| Cathepsin F | Endopeptidase, Cys/CA | n.i. | 207.2 ± 14.2 | n.i. |
| Cathepsin S | Endopeptidase, Cys/CA | n.i. | n.i. | n.i. |
| Cathepsin B | Endo- and carboxydipeptidase, Cys/CA | n.i. | n.i. | n.i. |
| Cathepsin X | Carboxypeptidase, Cys/CA | n.i. | n.i. | n.i. |
| Cathepsin C | Aminodipeptidase, Cys/CA | n.i. | n.i. | n.i. |
| Cathepsin H | Aminopeptidase, Cys/CA | n.i. | n.i. | n.i. |
| Model proteases | ||||
| Papain | Endopeptidase, Cys/CA | n.i. | 153.4 ± 11.1 | n.i. |
| Legumain | Endopeptidase, Cys/CD | n.i. | n.i. | n.i. |
| Cathepsin D | Endopeptidase, Asp/AA | n.i. | n.i. | n.i. |
| Chymotrypsin | Endopeptidase, Ser/PA | n.i. | n.i. | n.i. |
| Trypsin | Endopeptidase, Ser/PA | n.i. | n.i. | n.i. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matoušková, Z.; Orsághová, K.; Srb, P.; Pytelková, J.; Kukačka, Z.; Buša, M.; Hajdušek, O.; Šíma, R.; Fábry, M.; Novák, P.; et al. An Unusual Two-Domain Thyropin from Tick Saliva: NMR Solution Structure and Highly Selective Inhibition of Cysteine Cathepsins Modulated by Glycosaminoglycans. Int. J. Mol. Sci. 2024, 25, 2240. https://doi.org/10.3390/ijms25042240
Matoušková Z, Orsághová K, Srb P, Pytelková J, Kukačka Z, Buša M, Hajdušek O, Šíma R, Fábry M, Novák P, et al. An Unusual Two-Domain Thyropin from Tick Saliva: NMR Solution Structure and Highly Selective Inhibition of Cysteine Cathepsins Modulated by Glycosaminoglycans. International Journal of Molecular Sciences. 2024; 25(4):2240. https://doi.org/10.3390/ijms25042240
Chicago/Turabian StyleMatoušková, Zuzana, Katarína Orsághová, Pavel Srb, Jana Pytelková, Zdeněk Kukačka, Michal Buša, Ondřej Hajdušek, Radek Šíma, Milan Fábry, Petr Novák, and et al. 2024. "An Unusual Two-Domain Thyropin from Tick Saliva: NMR Solution Structure and Highly Selective Inhibition of Cysteine Cathepsins Modulated by Glycosaminoglycans" International Journal of Molecular Sciences 25, no. 4: 2240. https://doi.org/10.3390/ijms25042240