
Incidence of Phage Capsid Organization on the Resistance to High Energy Proton Beams

 , , ,
, , ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteriophages
2.2. Phage Propagation
2.3. Phage Collection
2.4. Proton Irradiations
2.5. Dose Absorbed (Da) and Dose Rate
2.6. Phage Titration (TU/mL)
2.7. Radiation Damage (RD)
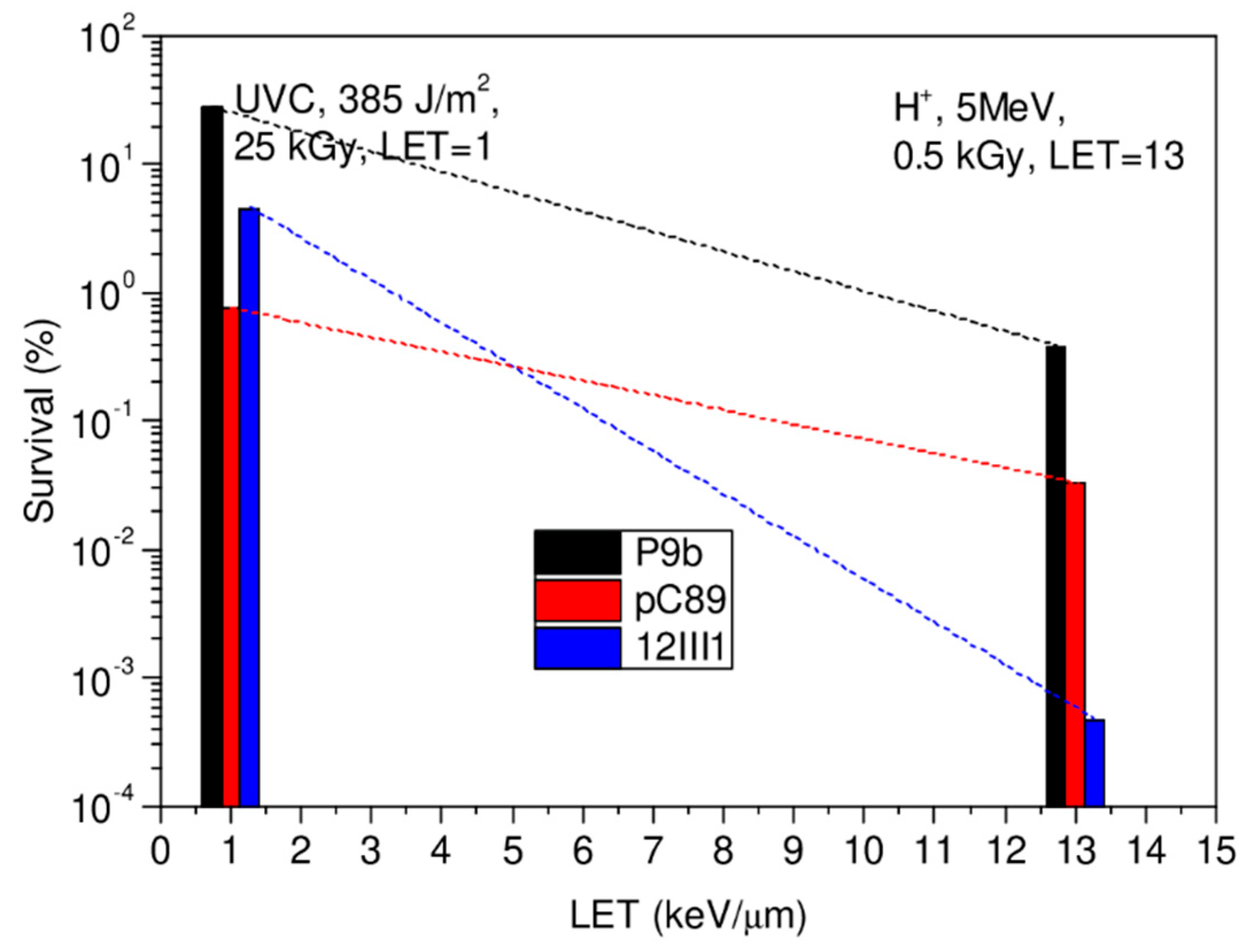
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Roychoudhury, S.; Das, A.; Sengupta, P.; Dutta, S.; Roychoudhury, S.; Choudhury, A.P.; Ahmed, A.B.F.; Bhattacharjee, S.; Slama, P. Viral Pandemics of the Last Four Decades: Pathophysiology, Health Impacts and Perspectives. Int. J. Environ. Res. Public Health 2020, 17, 9411. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Dennehy, J.J. Evolutionary ecology of virus emergence. Ann. N. Y. Acad. Sci. 2017, 1389, 124–146. [Google Scholar] [CrossRef]
- Snedden, C.E.; Makanani, S.K.; Schwartz, S.T.; Gamble, A.; Blakey, R.V.; Borremans, B.; Helman, S.K.; Espericueta, L.; Valencia, A.; Endo, A.; et al. SARS-CoV-2: Cross-scale Insights from Ecology and Evolution. Trends Microbiol. 2021, 29, 593–605. [Google Scholar] [CrossRef]
- Excler, J.-L.; Saville, M.; Berkley, S.; Kim, J.H. Vaccine development for emerging infectious diseases. Nat. Med. 2021, 27, 591–600. [Google Scholar] [CrossRef]
- Perlmutter, J.D.; Hagan, M.F. Mechanisms of virus assembly. Annu. Rev. Phys. Chem. 2015, 66, 217–239. [Google Scholar] [CrossRef] [Green Version]
- Prasad, B.V.V.; Schmid, M.F. Principles of virus structural organization. In Viral Molecular Machines; Rossmann, M.G., Rao, V.B., Eds.; Springer: Boston, MA, USA, 2012; pp. 17–47. [Google Scholar]
- Petrenko, V.A. Landscape Phage: Evolution from Phage Display to Nanobiotechnology. Viruses 2018, 10, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.P.; Petrenko, V.A. Phage display. Chem. Rev. 1997, 97, 391–410. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Liu, I.J.; Lu, R.M.; Wu, H.C. Advancement and applications of peptide phage display technology in biomedical science. J. Biomed. Sci. 2016, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, J.W.; Yang, L.; De Plano, L.M.; Stackhouse, M.A.; Petrenko, V.A. Evolution of a landscape phage library in a mouse xenograft model of human breast cancer. Viruses 2019, 11, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Liu, P.; Petrenko, V.A.; Liu, A.H. A Label-Free Electrochemical Impedance Cytosensor Based on Specific Peptide-Fused Phage Selected from Landscape Phage Library. Sci. Rep. 2016, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Wang, D.; Yan, L.; Petrenko, V.A.; Liu, A. Specific phages-based electrochemical impedimetric immunosensors for label-free and ultrasensitive detection of dual prostate-specific antigens. Sens. Actuators B Chem. 2019, 297, 126727. [Google Scholar] [CrossRef]
- Rizzo, M.G.; Carnazza, S.; De Plano, L.M.; Franco, D.; Nicolò, M.S.; Zammuto, V.; Petralia, S.; Calabrese, G.; Gugliandolo, C.; Conoci, S.; et al. Rapid detection of bacterial pathogens in blood through engineered phages-beads and integrated Real-Time PCR into MicroChip. Sens. Actuators B Chem. 2021, 329, 129227. [Google Scholar] [CrossRef]
- Yoo, S.Y.; Merzlyak, A.; Lee, S.W. Synthetic phage for tissue regeneration. Mediat. Inflamm. 2014, 2014, 192790. [Google Scholar] [CrossRef] [Green Version]
- Bazan, J.; Całkosiński, I.; Gamian, A. Phage display-a powerful technique for immunotherapy: 1. Introduction and potential of therapeutic applications. Hum. Vaccines Immunother. 2012, 8, 1817–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrenko, V.A.; Gillespie, J.W.; Xu, H.; O’Dell, T.; De Plano, L.M. Combinatorial avidity selection of mosaic landscape phages targeted at breast cancer cells-an alternative mechanism of directed molecular evolution. Viruses 2019, 11, 785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, E.C.; Jakobson, C.M.; Favor, A.H.; Lobba, M.J.; Álvarez-Benedicto, E.; Francis, M.B.; Tullman-Ercek, D. Quantitative characterization of all single amino acid variants of a viral capsid-based drug delivery vehicle. Nat. Commun. 2018, 9, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.; Shi, J.; Liu, A. Novel biotemplated MnO2 1D nanozyme with controllable peroxidase-like activity and unique catalytic mechanism and its application for glucose sensing. Sens. Actuators B Chem. 2017, 252, 919–926. [Google Scholar] [CrossRef]
- De Plano, L.M.; Scibilia, S.; Rizzo, M.G.; Crea, S.; Franco, D.; Mezzasalma, A.M.; Guglielmino, S.P.P. One-step production of phage–silicon nanoparticles by PLAL as fluorescent nanoprobes for cell identification. Appl. Phys. A 2018, 124, 222. [Google Scholar] [CrossRef]
- Torrisi, L.; Guglielmino, S.; Silipigni, L.; de Plano, L.M.; Kovacik, L.; Lavrentiev, V.; Torrisi, A.; Fazio, M.; Fazio, B.; di Marco, G. Study of gold nanoparticle transport by M13 phages towards disease tissues as targeting procedure for radiotherapy applications. Gold Bull. 2019, 52, 135–144. [Google Scholar] [CrossRef]
- Henry, K.A.; Arbabi-Ghahroudi, M.; Scott, J.K. Beyond phage display: Non-traditional applications of the filamentous bacteriophage as a vaccine carrier, therapeutic biologic, and bioconjugation scaffold. Front. Microbiol. 2015, 6, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateu, M.G. Virus engineering: Functionalization and stabilization. Protein Eng. Des. Sel. 2011, 24, 53–63. [Google Scholar] [CrossRef]
- Howorka, S. Molecular assembly in natural and engineered systems. Prog. Mol. Biol. Transl. Sci. 2011, 103, 414. [Google Scholar]
- Berchtikou, A.; Sokullu, E.; Nahar, S.; Tijssen, P.; Gauthier, M.A.; Ozaki, T. Comparative study on the inactivation of MS2 and M13 bacteriophages using energetic femtosecond lasers. J. Biophotonics 2020, 13, e202000109. [Google Scholar] [CrossRef]
- De Plano, L.M.; Franco, D.; Rizzo, M.G.; Zammuto, V.; Gugliandolo, C.; Silipigni, L.; Torrisi, L.; Guglielmino, S.P.P. Role of Phage Capsid in the Resistance to UV-C Radiations. Int. J. Mol. Sci. 2021, 22, 3408. [Google Scholar] [CrossRef] [PubMed]
- Morag, O.; Sgourakis, N.G.; Baker, D.; Goldbourt, A. The NMR-Rosetta capsid model of M13 bacteriophage reveals a quadrupled hydrophobic packing epitope. Proc. Natl. Acad. Sci. USA 2015, 112, 971–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumova, K.; Cosa, G. Overview of reactive oxygen species. In Singlet Oxygen: Applications in Biosciences and Nanosciences; The Royal Society of Chemistry: London, UK, 2016; Chapter 1; Volume 1, pp. 1–21. [Google Scholar]
- Lomax, M.E.; Folkes, L.K.; O’Neill, P. Biological consequences of radiation-induced DNA damage: Relevance to radiotherapy. Clin. Oncol. 2013, 25, 578–585. [Google Scholar] [CrossRef] [Green Version]
- Zammuto, V.; Fuchs, F.M.; Fiebrandt, M.; Stapelmann, K.; Ulrich, N.J.; Maugeri, T.L.; Pukall, R.; Gugliandolo, C.; Moeller, R. Comparing spore resistance of Bacillus strains isolated from hydrothermal vents and spacecraft assembly facilities to environmental stressors and decontamination treatments. Astrobiology 2018, 18, 1425–1434. [Google Scholar] [CrossRef]
- Felici, F.; Castagnoli, L.; Musacchio, A.; Jappelli, R.; Cesareni, G. Selection of antibody ligands from a large library of oligopeptides express on a multivalente exposition vector. J. Mol. Biol. 2019, 222, 301–310. [Google Scholar] [CrossRef]
- Luzzago, A.; Felici, F. Construction of disulfide-constrained random peptide libraries displayed on phage coat protein VIII. Methods Mol. Biol. 1998, 87, 155–164. [Google Scholar]
- Carnazza, S.; Foti, C.; Gioffrè, G.; Felici, F.; Guglielmino, S.P.P. Specific and selective probes for Pseudomonas aeruginosa from phage-displayed random peptide libraries. Biosens. Bioelectron. 2008, 23, 1137–1144. [Google Scholar] [CrossRef]
- De Plano, L.M.; Carnazza, S.; Franco, D.; Rizzo, M.G.; Conoci, S.; Petralia, S.; Nicoletti, A.; Zappia, M.; Campolo, M.; Esposito, E.; et al. Innovative IgG biomarkers based on phage display microbial amyloid mimotope for state and stage diagnosis in Alzheimer’s disease. ACS Chem. Neurosci. 2020, 11, 1013–1026. [Google Scholar] [CrossRef]
- Kay, B.; Winter, J.; McCafferty, J. Phage Display of Poplides and Proteins: A Laboratory Manual; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Torrisi, L.; Havranek, V.; Cutroneo, M.; Torrisi, A. Gafchromic HD-V2 investigations using MeV ion beams in vacuum. Radiat. Eff. Defects Solids 2019, 174, 1063–1075. [Google Scholar] [CrossRef]
- Nuclear Physics Institute CAS, Rez, Czech Republic. 2021. Available online: http://www.ujf.cas.cz/en/research-development/large-research-infrastructures-and-centres/canam/about-the-project/ (accessed on 15 November 2021).
- Cutroneo, M.; Havranek, V.; Torrisi, A.; Mackova, A.; Malinsky, P.; Slepicka, P.; Sofer, Z.; Torrisi, L. Polydimethylsiloxane–graphene oxide composite improving performance by ion beam irradiation. Surf. Interface Anal. 2020, 52, 1156–1162. [Google Scholar] [CrossRef]
- Torrisi, L.; Silipigni, L.; Manno, D.; Serra, A.; Nassisi, V.; Cutroneo, M.; Torrisi, A. Investigations on graphene oxide for ion beam dosimetry applications. Vacuum 2020, 178, 109451. [Google Scholar] [CrossRef]
- Ziegler, J.F.; Ziegler, M.D.; Biersack, J.P. SRIM—The stopping and range of ions in matter. Nucl. Instrum. Methods B 2010, 268, 1818–1823. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.L.; Stanley, J.A.; Robinson, M.C.; Sher, J.W.; Li, Z.; Chan, Y.A.; Omdahl, A.R.; Wattiez, R.; Godzik, A.; Matallana-Surget, S. Protein structure, amino acid composition and sequence determine proteome vulnerability to oxidation-induced damage. EMBO J. 2020, 39, e104523. [Google Scholar] [CrossRef] [PubMed]
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of ionizing radiation on biological molecules-mechanisms of damage and emerging methods of detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef]
- Stopar, D.; Spruijt, R.B.; Wolfs, C.J.; Hemminga, M.A. Protein-lipid interactions of bacteriophage M13 major coat protein. Biochim. Biophys. Acta 2003, 1611, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Vos, W.L.; Schor, M.; Nazarov, P.V.; Koehorst, R.B.M.; Spruijt, R.B.; Hemminga, M.A. Structure of membrane-embedded M13 major coat protein is insensitive to hydrophobic stress. Biophys. J. 2007, 93, 3541–3547. [Google Scholar] [CrossRef] [Green Version]
- Hemminga, M.A.; Vos, W.L.; Nazarov, P.V.; Koehorst, R.B.; Wolfs, C.J.; Spruijt, R.B.; Stopar, D. Viruses: Incredible nanomachines. New advances with filamentous phages. Eur. Biophys. J. 2010, 39, 541–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, R.; Pribil, W.; Appelt, S.; Gehringer, P.; Eschweiler, H.; Leth, H.; Cabaj, A.; Haider, T. Inactivation of bacteriophages in water by means of non-ionizing (UV-253.7 nm) and ionizing (gamma) radiation: A comparative approach. Water Res. 2001, 35, 3109–3116. [Google Scholar] [CrossRef]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]






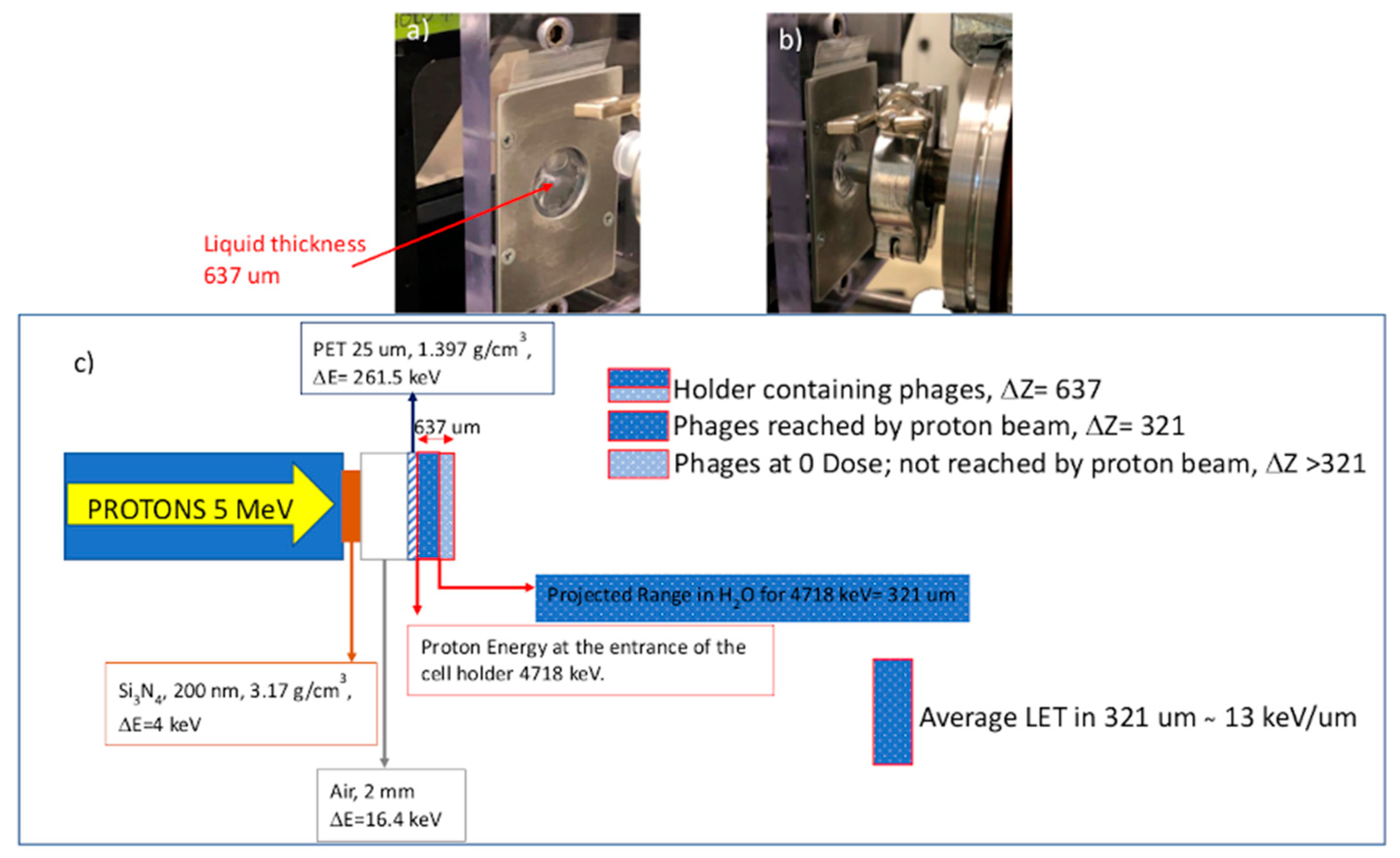{kind=link}
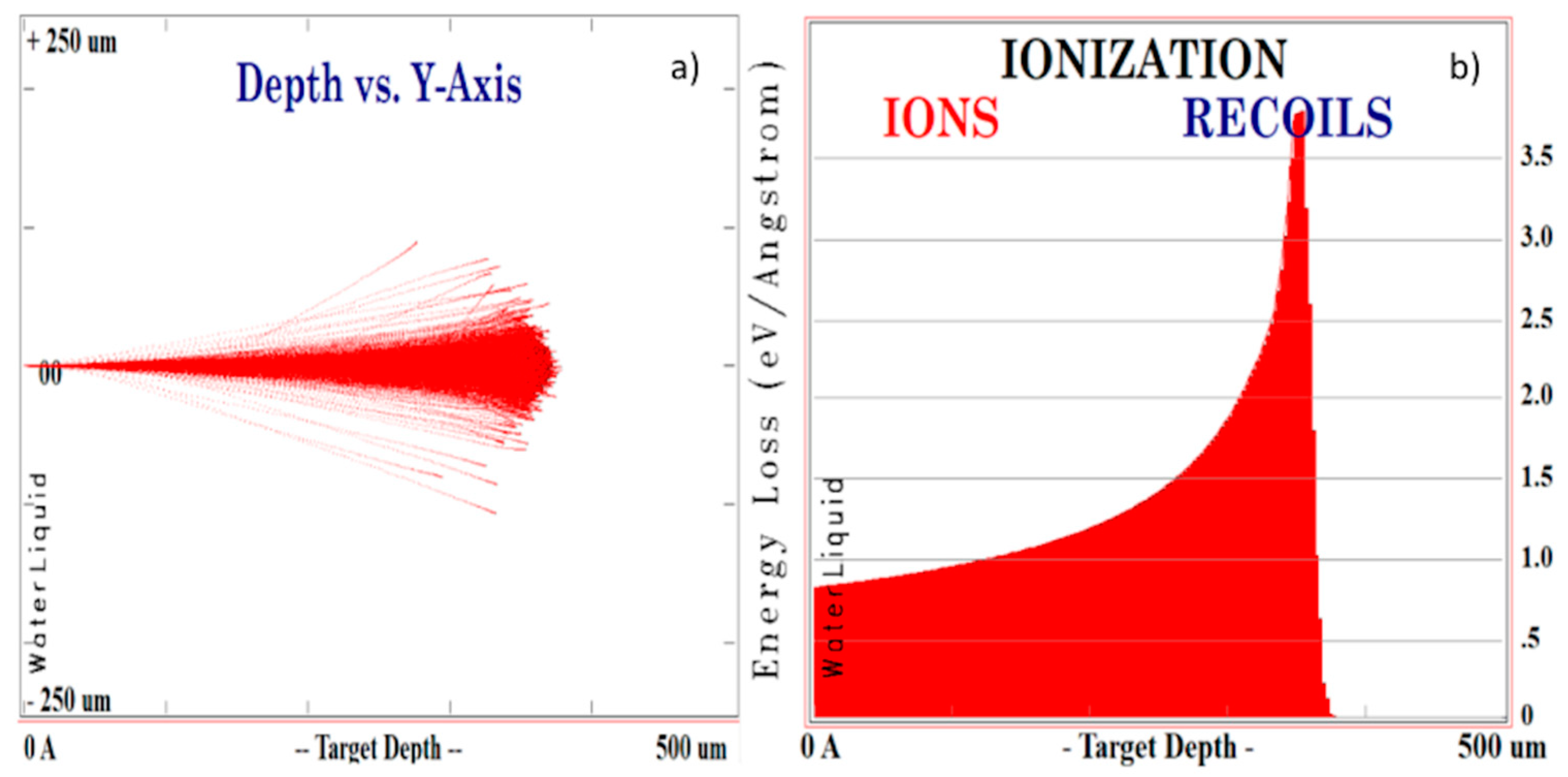{kind=link}
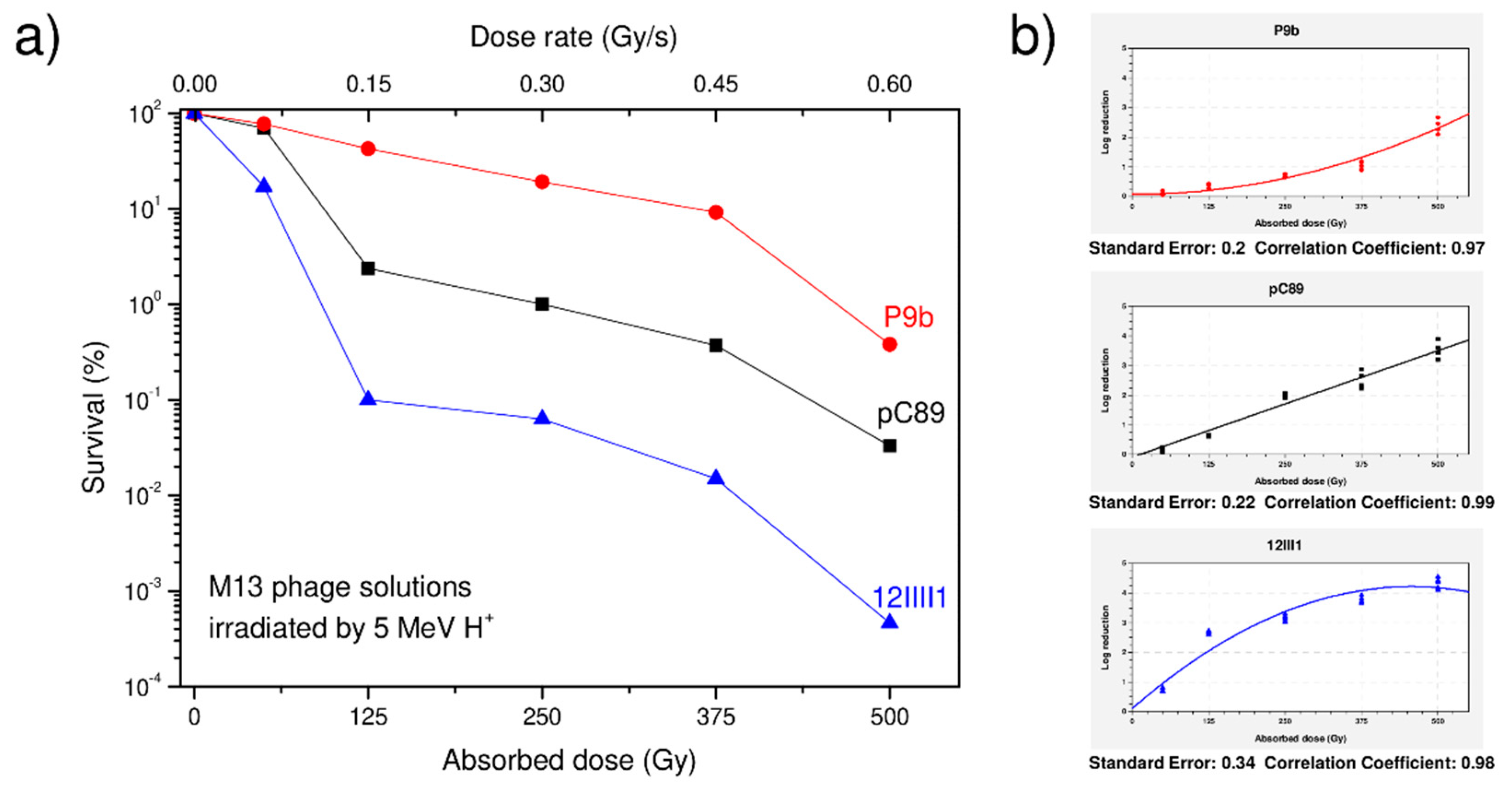{kind=link}
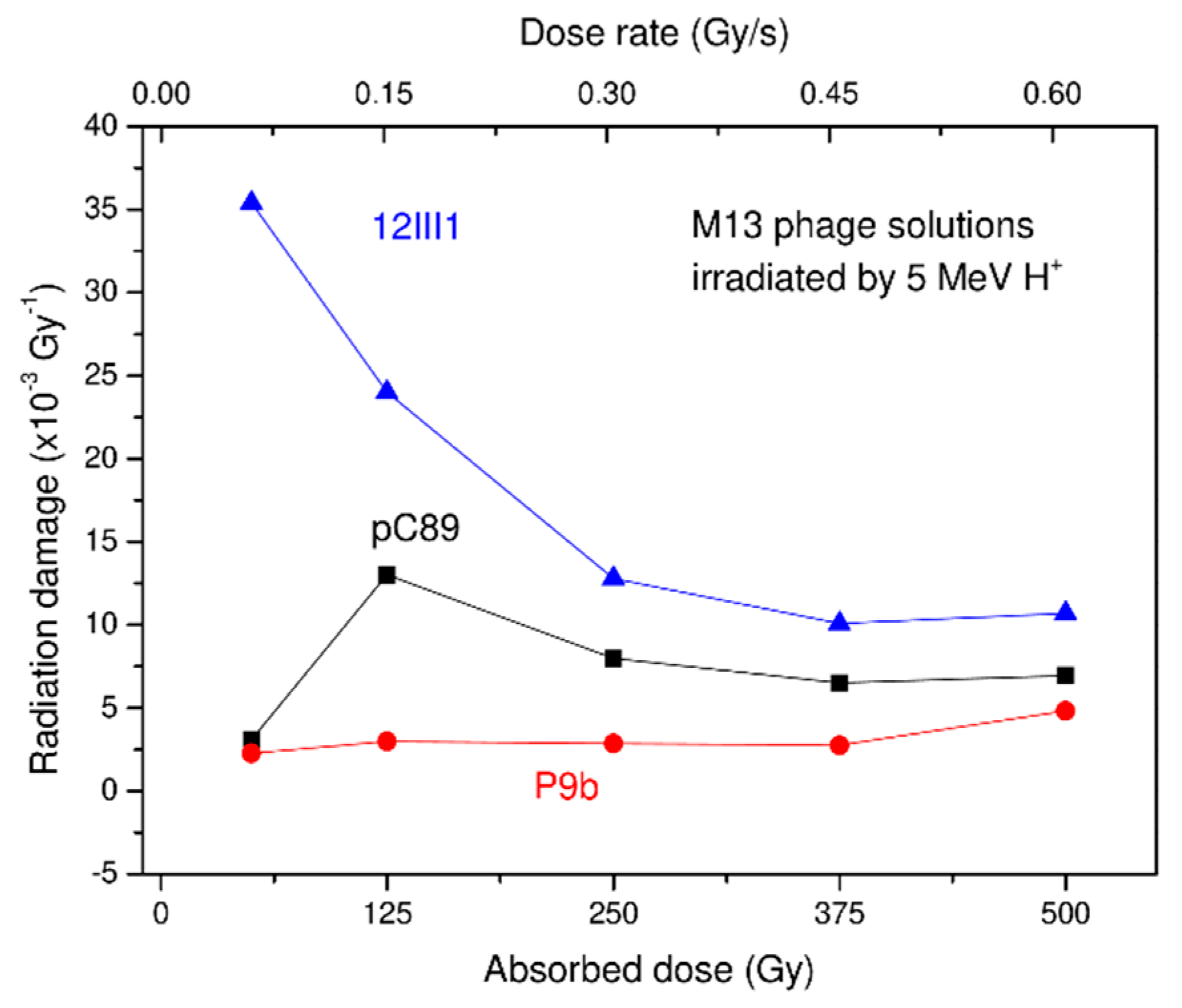{kind=link}
{kind=link}
| Dose (Gy) | ||||
| 50 | 125 | 250 | 375 | 500 |
| Dose Rate (Gy/s) | ||||
| 0.0615 | 0.155 | 0.309 | 0.463 | 0.615 |
| Phage Clone | Exposed | Dose (Gy) | ||||
|---|---|---|---|---|---|---|
| 50 Gy | 125 Gy | 250 Gy | 375 Gy | 500 Gy | ||
| pC89 | (3.0 ± 0.14) × 1010 (100%) | (2.1 ± 0.3) × 1010 (70%) | (7.1 ± 0.27) × 108 (2.37%) | (3.0 ± 0.41) × 108 (1%) | (1.1 ± 0.8) × 108 (0.37%) | (1.0 ± 0.5) × 107 (0.033%) |
| P9b (QRKLAAKLT) | (2.6 ± 0.21) × 1010 (100%) | (2.0 ± 0.25)×1010 (77%) | (1.1 ± 0.17) × 1010 (42.3%) | (5.0 ± 0.34) × 109 (19.2%) | (2.4 ± 0.45) × 109 (9.2%) | (1.0 ± 0.49) × 108 (0.38%) |
| 12III1 (RWPPHFEWHFDD) | (3.0 ± 0.38) × 1010 (100%) | (5.1 ± 0.23) × 108 (17%) | (3.0 ± 0.39) × 107 (0.1%) | (1.9 ± 0.5) × 107 (0.063%) | (4.7 ± 0.48) × 106 (0.0157%) | (1.4 ± 0.51) × 104 (0.00047 × 10%) |
| Phage Clone | 50 Gy | 125 Gy | 250 Gy | 375 Gy | 500 Gy |
|---|---|---|---|---|---|
| pC89 | 3.1 | 13 | 8.0 | 6.5 | 6.95 |
| P9b (QRKLAAKLT) | 2.28 | 2.99 | 2.86 | 2.75 | 4.83 |
| 12III1 (RWPPHFEWHFDD) | 35.4 | 24.0 | 12.8 | 10.1 | 10.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Plano, L.M.; Silipigni, L.; Torrisi, L.; Torrisi, A.; Cutroneo, M.; Havranek, V.; Mackova, A.; Zammuto, V.; Gugliandolo, C.; Rizzo, M.G.; et al. Incidence of Phage Capsid Organization on the Resistance to High Energy Proton Beams. Appl. Sci. 2022, 12, 988. https://doi.org/10.3390/app12030988
De Plano LM, Silipigni L, Torrisi L, Torrisi A, Cutroneo M, Havranek V, Mackova A, Zammuto V, Gugliandolo C, Rizzo MG, et al. Incidence of Phage Capsid Organization on the Resistance to High Energy Proton Beams. Applied Sciences. 2022; 12(3):988. https://doi.org/10.3390/app12030988
Chicago/Turabian StyleDe Plano, Laura Maria, Letteria Silipigni, Lorenzo Torrisi, Alfio Torrisi, Mariapompea Cutroneo, Vladimir Havranek, Anna Mackova, Vincenzo Zammuto, Concetta Gugliandolo, Maria Giovanna Rizzo, and et al. 2022. "Incidence of Phage Capsid Organization on the Resistance to High Energy Proton Beams" Applied Sciences 12, no. 3: 988. https://doi.org/10.3390/app12030988