
Dynamics of Silurian Plants as Response to Climate Changes
by
 ,
,
Josef Pšenička
1,* ,
,
Jiří Bek
2,
Jiří Frýda
3,4,
Viktor Žárský
2,5,6,
Monika Uhlířová
1,7 and
Petr Štorch
2 1
Centre of Palaeobiodiversity, West Bohemian Museum in Pilsen, Kopeckého sady 2, 301 00 Plzeň, Czech Republic
2
Laboratory of Palaeobiology and Palaeoecology, Geological Institute of the Academy of Sciences of the Czech Republic, Rozvojová 269, 165 00 Prague 6, Czech Republic
3
Faculty of Environmental Sciences, Czech University of Life Sciences Prague, Kamýcká 129, 165 21 Praha 6, Czech Republic
4
Czech Geological Survey, Klárov 3/131, 118 21 Prague 1, Czech Republic
5
Department of Experimental Plant Biology, Faculty of Science, Charles University, Viničná 5, 128 43 Prague 2, Czech Republic
6
Institute of Experimental Botany of the Czech Academy of Sciences, v. v. i., Rozvojová 263, 165 00 Prague 6, Czech Republic
7
Institute of Geology and Palaeontology, Faculty of Science, Charles University, Albertov 6, 128 43 Prague 2, Czech Republic
*
Author to whom correspondence should be addressed.
Life 2021, 11(9), 906; https://doi.org/10.3390/life11090906
Submission received: 4 August 2021
/
Revised: 24 August 2021
/
Accepted: 25 August 2021
/
Published: 31 August 2021
(This article belongs to the Special Issue Bryophyte Responses to Climate Changes)
Abstract
:The most ancient macroscopic plants fossils are Early Silurian cooksonioid sporophytes from the volcanic islands of the peri-Gondwanan palaeoregion (the Barrandian area, Prague Basin, Czech Republic). However, available palynological, phylogenetic and geological evidence indicates that the history of plant terrestrialization is much longer and it is recently accepted that land floras, producing different types of spores, already were established in the Ordovician Period. Here we attempt to correlate Silurian floral development with environmental dynamics based on our data from the Prague Basin, but also to compile known data on a global scale. Spore-assemblage analysis clearly indicates a significant and almost exponential expansion of trilete-spore producing plants starting during the Wenlock Epoch, while cryptospore-producers, which dominated until the Telychian Age, were evolutionarily stagnate. Interestingly cryptospore vs. trilete-spore producers seem to react differentially to Silurian glaciations—trilete-spore producing plants react more sensitively to glacial cooling, showing a reduction in species numbers. Both our own and compiled data indicate highly terrestrialized, advanced Silurian land-plant assemblage/flora types with obviously great ability to resist different dry-land stress conditions. As previously suggested some authors, they seem to evolve on different palaeo continents into quite disjunct specific plant assemblages, certainly reflecting the different geological, geographical and climatic conditions to which they were subject.
1. Introduction
The final stages of the colonization of land by plants (Embryophyta) within the Ordovician-Silurian interval is one of the most important global events in the evolution of life on the Earth. Wellman and Strother [1] and Kenrick [2] hypothesized that life existed on the land since over 2.7 Gyr in the form of communities of bacteria (including cyanobacteria) and archaea [3,4], that colonized shallow places in the fresh-water environment.
Successful terrestrialization of plants was enabled by some crucial adaptations including tissues ensuring the distribution of water and nutrients for growth, body protection against desiccation and UV, anchoring in the substrate, and propagation by resistant spores [5,6].
Ordovician and Silurian plant communities already included possibly several groups of polysporangiate land plants (no monosporangiates are documented by the fossil record), including more ancient and enigmatic cryptosporophytes [7,8], accompanied by bacterial-cyanobacterial mats [9,10,11]. At the end of the Silurian and the beginning of the Devonian it is already possible to quite reliably identify the first lycophytes (Baragwanathia and Asteroxylon).
The first evidence indicating the existence of land plants are cryptospores of the Early Middle Ordovician age (c. 470MA) [12]. However, their producers are still unknown. Some very questionable polysporangiophytes have been described from the Ordovician [13,14] that coincide with the appearance of the first true trilete spores within the Late Ordovician [15,16]. These spores indicate an earlier origin of possibly several types of polysporangiophyte-grade plants, probably during Ordovician. Some authors hypothesized the colonization of land by embryophytes during the Cambrian (or even Proterozoic) and this recently has been strongly supported by phylostratigraphic data [3,4,17,18].
This paper provides an overview and a revision of Silurian plant assemblages (from Wenlock to Přídolí) of polysporangiophytes with dispersed spores and cryptospores. Special attention is paid to the dynamics of the Silurian plant evolution within the context of Silurian climate changes linked with perturbations of the global carbon cycle (Figure 1). The data on Llandoverian plants are not included in the latter analysis because (1) observations of well-preserved macroplants below the Wenlock are extremely rare and (2) the plants are represented only by sterile bifurcated axes [19].
1.1. Character of Early Land Plants and Their Classification
Gerrienne et al. [25] established a currently widely accepted approach for the classification of the earliest embryophytes (including monosporangiate hornworts and mosses not reported from fossil data), which, along with obscure embryophytes and probably bacterial-cyanobacterial mats, formed essential components of the entire terrestrial ecosystem, including the well-known genera Nematothallus, Pachytheca and Prototaxites. Nematothallus, which probably belonged to lichenized fungi [26]. Many different-sized tubular structures from Silurian-Devonian sediments (probably composed of hyphae) are assigned to Prototaxites. Pachytheca, represented by small spherical structures of still unknown affinity, stressed some similarities to Prototaxites [27]. Nematasketum, classified as an enigmatic non-embryophyte plant [28], was described based on vegetative parts and possesses an axial organization with peripheral tissues forming a ring. Nematasketum shows some similarities with Prototaxites [28].
The first credible fossil bryophytes, represented by Riccardiothallus devonicus [15,29], are of the Pragian (Early Devonian) age. Morris et al. [15] estimated the appearance of bryophytes during the Late Cambrian–Late Ordovician interval based on molecular clock data. The Ordovician occurrence of bryophytes would also be supported by fossil spore exine ultrastructural similar to some spores of recent liverworts [30]. However, there is a recent consensus of monophyletic origin of the bryophytes and its sister relationship with tracheophytes (i.e., tracheophytes did not evolve from bryophytes [18,31]). This clearly indicates that it might not make much sense to speak about monosporangiate mosses in the Ordovician or Silurian without fossil support for a polysporangiate common ancestor of both mosses and tracheophytes, which is currently probable (see our Discussion).
Polysporangiophytes [25,32,33] formed the only embryophytic plants documented by the fossil in the Silurian. The group includes the first true vascular plants (Tracheophyta). Although recent vascular plants may reach tens of meters in height, the earliest terrestrial forms were mostly diminutive [8,34,35,36,37], although some forms of polysporangiophytes could already have been over 10 cm tall (e.g., Tichavekia grandis) [38]. The main character of polysporangiophytes is, as the name suggests, the branched sporophyte with more than one sporangium [39]. Silurian plant communities most commonly include polysporangiophyte taxa, such as Cooksonia, Salopella, Aberlemnia, Hedeia, Tortillicaulis, Fusiformitheca, Steganotheca and Caia. Sterile fragments of plant axes are referred to the genus Hostinella. All those genera are usually placed among “rhyniophytoids“ [40].
Basal lycophytes are represented by zosterophylls [33]. Silurian zosterophylls include Zosterophyllum, Bathurstia, Distichophytum, Macivera, and Aberlemnia with bivalved sporangia and dehiscence along the distal sporangial margin, which has led to the placement of these plants [38,41] in the lycophyte stem group. The first unambiguous lycophyte is Baragwanathia, occurring in the late Silurian [42,43,44], which already had well-developed microphylls.
1.2. Silurian Plant Assemblages
Silurian plant assemblages and their geographical distribution were studied in several papers [45,46,47,48,49]. We must be aware that only a limited number of plants can be found at one locality and it is not exceptional that only one species of polysporangiophyte occurs at a locality. It appears that phytogeographical differentiation and disjunction (isolation) of land-plant types/groups was significant during the Late Silurian (Přídolí) and Early Devonian [47,50]. Raymond et al. [47] established 35 macrofloristic assemblages for Silurian to Early Devonian times based on correspondence and cluster analyses for different geographical areas and climatic zones. The latter analyses resulted in the establishment of four phytogeographic units: (1) North Laurussian unit (Bathurst Island) located near the palaeoequator; (2) South Laurussian–Northwest Gondwanan unit (UK; Podolia, Ukraine and Bolivia) located approximately 18° S palaeolatitude to 60–75° S palaeolatitude; (3) Kazakhstanian unit located north of the palaeoequator; and (4) Northeast Gondwanan (Australian) unit at 10° S palaeolatitude [47]. Later Wellman et al. [48] established 14 assemblages but they did not revise palaeophytogeographical areas proposed by Raymond et al. [47] that are used herein.
2. Materials and Methods
Specimens/samples were selected from the Collections of the Centre of Palaeobiodiversity of the West Bohemian Museum in Pilsen—WBM (Czech Republic), specimens F21761a, F21762; SEM-rack SILI-A1, Kosov Quarry; National Museum—NM (Prague, Czech Republic), specimens D-475 Kosov Quarry, D-552b Loděnice, Špičatý vrch-Barrandovy Jámy; Czech Geological Survey (Prague, Czech Republic)—CGS, specimen KR 1, Karlštejn locality; Faculty of Sciences, Charles University, Prague (Czech Republic)—FSCU, Kosov Quarry; Swedish Museum of Natural History (Stockholm, Sweden)—SMNH, specimen JE-Sch0260B, Kosov Quarry; private collections of Ondřej Zicha—PCC (Czech Republic), Špičatý vrch-Barrandovy Jámy and the collection of the University of Saskatchewan—US (Saskatoon, Canada), specimens US600-6791, US600-8144, US384-8137, US688-8152, US385-2398, Bathurst Island, Canada. Specimens from NM, WBM, CGS, FSCU, SMNH and PCC represented polysporangiophyte assemblages from the temperate zone of the lower Wenlock (Sheinwoodian) and Přídolí of the Barrandian area, the Prague Basin (Czech Republic). Mesofossils from the upper Wenlock (Homerian, parvus-nassa Biozone) of the Lištice locality are stored in WBM showing small plant fragments macerated from the rock (HCL 32% (5–10 days)-HF35% (5–8 days)-HNO3 65% (several minutes–10 days). Specimens of polysporangiophytes from the tropical zone of the Gorstian age, Ludlow (Canada) are stored in the US.
3. Results
3.1. Environment of Silurian Plants
Early land/polysporangiate plants diversified and colonized continents during Silurian-Devonian “terrestrial radiation” [51]. Recent analysis convincingly indicates a possibility that a great land plant radiation had already occurred in the Ordovician and argues that land plants were possibly not affected by the end-Ordovician extinction event [52]. Servais et al. [53] stated the importance of environmental factors, including atmospheric composition and the geochemical carbon cycle [54,55], as major influencers of the evolutionary dynamics of early plants. The Silurian Period has been considered as a stable interval characterized by gradual, long-term climatic change starting from the Middle to Late Ordovician cold climate and culminating in Silurian greenhouse conditions [56,57]. The Silurian, lasting from approximately 443.1 to 419.0 million years [22], represents a key interval in the biological evolution of marine ecosystems [58,59,60,61]. The transitional character of the Silurian is not illustrated only by the changes of marine communities and ecosystems, but also by the instability of the global carbon cycle including rapid changes in atmospheric pO2 and pCO2 [62,63,64]. Published data indicate that during the relatively short Silurian interval at least five globally observed carbon isotope excursions have been recognized (Figure 1). These anomalies include the δ13C excursions in the mid-Llandovery, early Wenlock, late Wenlock, late Ludlow, and across the Silurian-Devonian boundary [21,65]. Geochemical events were closely linked to major crises in marine ecosystems as well as to palaeoclimatic changes [24,56,66,67,68,69].
Global temperature, together with total precipitation, are key parameters influencing terrestrial community development. However, the reconstruction of global temperature change is complicated and has to be based on samples from different palaeoregions because ocean temperature varied with season, water depth, and palaeolatitude [23]. In addition, to calculate palaeotemperatures from marine calcite or apatite δ18O values, the oxygen isotope composition of the Silurian seawater has to be assumed. However, there is no consensus on whether the oxygen isotope composition of seawater evolved during the Phanerozoic [70,71] or was more or less constant [72]. Thus, without a better constrained Silurian seawater δ18O, the calculation of absolute temperatures remains speculative. However, relative temperature changes can be calculated independent of seawater δ18O (Figure 1).
Grossman and Joachimski [23] mentioned that the oxygen isotope record for Silurian brachiopods and conodonts from tropical and subtropical palaeolatitudes showed a Llandovery Warm Trend. On the other hand, Silurian brachiopod data revealed a Ludfordian Cool Event. New δ18O conodont data from Ludfordian temperate as well as tropical palaeolatitudes [24] confirms the marked cooling of sea surface temperatures at time a significant eustatic sea-level fall occurred on several palaeocontinents. These observations, interpreted as a major glaciation in polar and subpolar Gondwana, are referred to as the ‘Mid-Ludfordian Glaciation’ [24].
Taken together, recently available sedimentological (glacial diamictite deposition) and geochemical (δ18O) data suggest at least five glacial events within the Silurian Period, namely early Aeronian, late Aeronian, early Sheinwoodian, Homerian, and mid-Ludfordian glaciation [24,56,66,69,73] (Figure 1).
3.2. Diversity of Silurian Plants
Raymond et al. [47] proposed the following palaeophytogeographical units.
3.2.1. North Laurussian Unit
This unit, located near the palaeoequator, was of tropical character (Figure 2—La, Lb, Lc, L3, La) and includes north Laurussian basins. The oldest plant fossil record is known from the Ludlow strata (Figure 3) of Bathurst Island [45,74] and North Greenland [75]. The Bathurst Island plant assemblage provides several terrestrial species, e.g.,: Cf. Bathurstia sp., Zosterophyllum sp., Distichophytum sp., Aff. Zosterophyllum sp. A, Macivera gracilis [74] (Figure 4). The only plant from North Greenland is Salopella sp. [75]. Generally, this plant assemblage can be characterized as a zosterophyll-dominated flora with minor rhyniophytes.
3.2.2. South Laurussian–Northwest Gondwanan Unit
This unit includes territories from approximately 18° S palaeolatitude to 60–75° S palaeolatitude [75] and was probably a part of the temperate climatic zone (Figure 2). The fossil record of this palaeophytogeographical unit ranges from early Wenlock (Sheinwoodian) to Přídolí.
Northwest Gondwana
Northwest Gondwana (peri-Gondwana) provides the oldest true vascular land plants, Cooksonia barrandei and Cooksonia sp. (Figure 5), from the Czech Republic (Barrandian area, the Prague Basin) [77,78]. These globally important specimens come from the Monograptus belophorus Biozone of the Sheinwoodian Age. Sheinwoodian localities of the Barrandian area yielded several unpublished early land plants of different affinities (rhyniophytes, probably zosterophylls, cryptosporophytes) [77,78,79,80]. Current research suggests that cooksonioid dominated plant assemblages were accompanied by Ascomycota [81], Prototaxites sp., and enigmatic Pachytheca [82]. Libertín et al. [83] also recorded Cooksonia sp. from the same geographic area but from the stratigraphically younger Cyrtograptus rigidus Biozone (Figure 3—Wa). Generally, the latter assemblage (Figure 3—W1 plus Wa) is characterized by relatively rich vegetation cover of the temperate zone of the Southern hemisphere. The other probable plants, such as Prototaxites sp. (Figure 6e,f), Pachytheca sp. (Figure 6a,b), cf. Cosmochlaina (Figure 6d), and cf. Nematasketum (Figure 6c) belong to relatively abundant fossils in the Homerian interval.
The relatively rich plant assemblage from the Barrandian area (the Prague Basin) comes from Přídolí strata (Figure 2—P9, Pb). Mineral nutrient-rich volcanic islands formed from late Wenlock–early Ludlow Kosov and Svatý Jan volcanic centres could have provided suitable conditions for terrestrialization of early land plants [38,82,84,85]. Plants from the Skalograptus parultimus-Skalograptus ultimus Biozone interval (Figure 3—P9) include Cooksonia sp. (Figure 7f), Cooksonia cf. hemisphaerica (Figure 7a), Aberlemnia bohemica (Figure 7e), Fusiformitheca sp. (Figure 7d), and Tichavekia grandis (Figure 7c) [38,86,87]. Not only fertile but also abundant sterile axial parts of Cooksonia affinity are reported [82,88]. Occurrence of the basal lycophyte Baragwanathia brevifolia (Figure 7b) [43] from Skalograptus ultimus Biozone is unique, representing the single finding of a lycophyte with differentiated microphylls standing out among other Silurian plants of the Barrandian area, which, in contrast, exhibit ancient rhyniophytoid characters.
Figure 3.
Stratigraphical distribution of plants or plant assemblages from the Wenlock (W) through Ludlow (L) to Přídolí (P). Quantitative curves of cryptospores (see Figure 8) and isotopic temperature curves (see Figure 1) put together. The circle at the end of the connecting line between the assemblage/plant unit and stratigraphy levels indicates the accuracy of the stratigraphic determination-the more right this point is, the more accurate the determination. Assemblages: W1 = Barrandian area, the Prague Basin, Czechia [77,78,79,80]; W2 = Souther Bolivia [89]; W3 = Gasport Stone Quarry, Niagara County, New York State, USA [90]; L1 = Welsh Basin (Cwm Craig Ddu, Powys) [91]; L2 = Welsh Basin (Capel Horeb, Powys) [90]; L3 = Bathurst Island, Arctic Canada [74]; L4 = Victoria, Australia, Centennial Beds [42,92]; L5 = Victoria, Australia, Monograptus Beds [42,92]; L6 = Tarija, southern Bolivia [93]; L7 = Gotland, Sweden [94]; P1 = Anglo-Welsh Basin (Ludford Corner, Ludlow) [95]; P2 = Anglo-Welsh Basin (Perton Lane, Hereford) [96,97,98]; P3 = Anglo-Welsh Basin (Freshwater East, Pembrokeshire) [99]; P4 = Anglo-Welsh Basin (Capel Horeb, Powys) [91]; P5 = New York State, USA [90]; P6 = Ontario, Canada [90]; P7 = Holy Cross Mts., Poland [100]; P8 = Podolia, Ukraine [101]; P9 = Barrandian area, the Prague Basin, Czechia [38,43]; P10 = Kazakhstan [45]; P11 = Kazakhstan, Balkhash area [102,103]; P12 = Junggar Basin, Xinjiang, China [104]; P13 = Northern Vietnam [105]; Isolated finding of plants: Wa = Barrandian area, the Prague Basin, Czechia [83]; Wb = Tipperary, Ireland [106]; La = Northern Greenland [75]; Lb = Bathurst Island, Arctic Canada [74]; Lc = Bathurst Island, Arctic Canada [74]; Pa = Anglo-Welsh Basin (Little Wallop Hall, Shropshire) [107]; Pb = Barrandien area, the Prague Basin, Czechia [82]; Pc = Homra Basin, Libya [108]; Pd = Yunan, China [50]; Pe = Junggar Basin, Xinjiang, China [104]; Pf = Shropshire, UK [7].
Figure 3.
Stratigraphical distribution of plants or plant assemblages from the Wenlock (W) through Ludlow (L) to Přídolí (P). Quantitative curves of cryptospores (see Figure 8) and isotopic temperature curves (see Figure 1) put together. The circle at the end of the connecting line between the assemblage/plant unit and stratigraphy levels indicates the accuracy of the stratigraphic determination-the more right this point is, the more accurate the determination. Assemblages: W1 = Barrandian area, the Prague Basin, Czechia [77,78,79,80]; W2 = Souther Bolivia [89]; W3 = Gasport Stone Quarry, Niagara County, New York State, USA [90]; L1 = Welsh Basin (Cwm Craig Ddu, Powys) [91]; L2 = Welsh Basin (Capel Horeb, Powys) [90]; L3 = Bathurst Island, Arctic Canada [74]; L4 = Victoria, Australia, Centennial Beds [42,92]; L5 = Victoria, Australia, Monograptus Beds [42,92]; L6 = Tarija, southern Bolivia [93]; L7 = Gotland, Sweden [94]; P1 = Anglo-Welsh Basin (Ludford Corner, Ludlow) [95]; P2 = Anglo-Welsh Basin (Perton Lane, Hereford) [96,97,98]; P3 = Anglo-Welsh Basin (Freshwater East, Pembrokeshire) [99]; P4 = Anglo-Welsh Basin (Capel Horeb, Powys) [91]; P5 = New York State, USA [90]; P6 = Ontario, Canada [90]; P7 = Holy Cross Mts., Poland [100]; P8 = Podolia, Ukraine [101]; P9 = Barrandian area, the Prague Basin, Czechia [38,43]; P10 = Kazakhstan [45]; P11 = Kazakhstan, Balkhash area [102,103]; P12 = Junggar Basin, Xinjiang, China [104]; P13 = Northern Vietnam [105]; Isolated finding of plants: Wa = Barrandian area, the Prague Basin, Czechia [83]; Wb = Tipperary, Ireland [106]; La = Northern Greenland [75]; Lb = Bathurst Island, Arctic Canada [74]; Lc = Bathurst Island, Arctic Canada [74]; Pa = Anglo-Welsh Basin (Little Wallop Hall, Shropshire) [107]; Pb = Barrandien area, the Prague Basin, Czechia [82]; Pc = Homra Basin, Libya [108]; Pd = Yunan, China [50]; Pe = Junggar Basin, Xinjiang, China [104]; Pf = Shropshire, UK [7].

Sterile rhyniophytes were described from the Late Wenlock/Ludlow strata of southern Bolivia [89] (Figure 3—W2). Stratigraphically younger specimens from the Lipeón Formation at Tarija of Ludlow age yielded cf. Aberlemnia caledonica, sterile axes assigned to the genus Hostinella, and isolated sporangia determined based on their morphology as cf. Cooksonia hemisphaerica, cf. Tarrantia and isolated sporangia [93] (Figure 3—L6). One of the best-preserved specimens of cf. Aberlemnia caledonica was earlier published by Morel et al. [109].
Daber [108] described the uppermost Přídolí flora from Libya (Figure 2—Pc), including Cooksonia sp. This area was situated near the northern edge of the Gondwana mainland, far south of the Barrandian area (the Prague Basin) during Přídolí times.
Figure 4.
Tropical flora assemblage from Bathurst Island, Canada. (a) cf. Bathurstia, US—US600-6791, scale bar = 5 mm; (b) Distichophytum sp. US—US600-8144, scale bar = 10 mm; (c) aff. Zosterophyllum sp. A, US—US688-8152, scale bar = 5 mm; (d) Macivera gracilis, US—US385-2398, scale bar = 10 mm; (e) Zosterophyllum sp., US—US384-8137, scale bar = 5 mm. Used with the permission of the authors [74].
Figure 4.
Tropical flora assemblage from Bathurst Island, Canada. (a) cf. Bathurstia, US—US600-6791, scale bar = 5 mm; (b) Distichophytum sp. US—US600-8144, scale bar = 10 mm; (c) aff. Zosterophyllum sp. A, US—US688-8152, scale bar = 5 mm; (d) Macivera gracilis, US—US385-2398, scale bar = 10 mm; (e) Zosterophyllum sp., US—US384-8137, scale bar = 5 mm. Used with the permission of the authors [74].

South Laurussia
The South Laurussian unit includes basins located in southwest Laurussia, situated approximately 30° south latitude [76], and belonging to temperate zone (Figure 2). One of those basins, the classic Welsh Basin, UK, has a sedimentological record ranging from Cambrian to Early Devonian. Rhyniophytoids are the only plants reported from the Late Silurian and Early Devonian of this area. The oldest polysporangiophyte, Cooksonia sp. (Figure 3—Wb) from Homerian strata of the Tipperary County (Ireland) of Laurussia, was described by Edwards et al. [106]. This area was located probably c. 25° S palaeolatitude [76]. It is not possible to speculate about the general character of this vegetation because only one species has been recorded.
Gorstian assemblages from the Welsh-Basin probably belonging to the same phytogeographical unit are known from the Anglo-Welsh Basin, UK [91] (Figure 2—L1). They are typified by rhyniophytoid-dominated vegetation with Cooksonia sp., cf. Cooksonia cambrensis, Cooksonia pertoni and sterile axes of Hostinella (Figure 3—L1).
Freshwater East in Pembrokeshire is a famous locality in the Welsh Basin (Figure 3—P3) that yielded a rich plant assemblage of the Přídolí age. Cooksonia and sterile axes of Hostinella were described by Edwards [99], including a species, Cooksonia cambrensis, and the rhyniophytoid Tortilicaulis transwalliensis, resembling sporophytes of some recent liverworts.
A new rhyniophytoid-like species, Caia langii, was described from Perton Lane, UK [96] (Figure 3—P2), which is the original locality for Cooksonia pertoni [110]. Another plant from the same locality, Pertonella dativelethra, is characterized by distinctive sporangial projections [97]. The other plants from this site are Cooksonia cambrensis and Salopella sp. [98].
Ludford Corner, UK yielded exceptionally preserved plants (Figure 3—P1). Capel Horeb in Wales, UK (Figure 3—P4) is another locality of the Welsh Basin typified by coalified plant adpressions [91], such as Steganotheca and sterile axes of Hostinella. Rogerson et al. [107] described Cooksonia pertoni from Little Wallop Hall in Shropshire, UK (Figure 3—Pa). Similar flora is known from Ontario, Canada (Figure 3—P6) and New York, USA (Figure 3—P5). The polysporangiophytes from Canada and New York as coalified adpressions are preserved in fine-grained dolomite [90]. This area was located at the southeastern edge of the Laurussia palaeocontinent, approximately 40° south latitude [76] during Přídolí. The research was concentrated on six localities, of which Cooksonia sp., Cooksonia cf. hemisphaerica, and sterile axes assigned to Hostinella have been recorded.
Figure 5.
Plant fossils from the South Laurussian–Northwest Gondwanan unit (temperate zone), Wenlock, Scheinwoodian, Barrandian area (the Prague Basin), Czechia; (a) Cooksonia barrandei, specimen D-552b, NM, Špičatý vrch-Barrandovy Jámy; (b) Cooksonia sp. PCC (notice—will be a part of collection of NM), Špičatý vrch-Barrandovy Jámy; scale bar for both specimens = 5 mm.
Figure 5.
Plant fossils from the South Laurussian–Northwest Gondwanan unit (temperate zone), Wenlock, Scheinwoodian, Barrandian area (the Prague Basin), Czechia; (a) Cooksonia barrandei, specimen D-552b, NM, Špičatý vrch-Barrandovy Jámy; (b) Cooksonia sp. PCC (notice—will be a part of collection of NM), Špičatý vrch-Barrandovy Jámy; scale bar for both specimens = 5 mm.

Poland (eastern Laurussia) belonged to the South Laurussian–Northwest Gondwanan unit (Figure 3—P7) at approximately 20° south latitude [76]. Bodzioch et al. [100] published specimens from early Přídolí and assigned them to rhyniophytoids. Specimens come from the Holy Cross Mountains (Poland), including fertile Cooksonia sp. and sterile axes described as Hostinella.
Ishchenko [101] described an assemblage from the Přídolí dominated by rhyniophytoids including Cooksonia pertoni and C. hemisphaerica from Podolia (Ukraine). This area is typical for warm zones from the same latitude such as the Polish localities [100]. Unlike Poland Zosterophyllum sp., Lycopodolica, and Eorhynia (Salopella) are reported.
Wenlockian sterile branched axes were reported from the Gasport Stone Quarry, Niagara County, (New York, USA) [90] (Figure 2—W3). The section belongs to the graptolite zone of Medusaegraptus—a graptolite taxon that is also recognized in the upper Sheinwoodian to lower Homerian of the Barrandian region of the Prague Basin (Figure 3—W3).
3.2.3. Kazakhstanian Unit
This unit was located north of the palaeoequator (Figure 1) and its vegetation belongs to the subtropical zone. Two sites yielded plants, as seen on Figure 2 and Figure 3—P10, P11, and correspond with the stratigraphical interval of the Skalograptus boucek –Skalograptus transgrediens graptolite biozones. The Kazakhstanian unit was situated on the northern hemisphere in the subequatorial zone of the Palaeo-Tethys Ocean [76]. The assemblage includes the genera previously reported by Edwards and Wellman [45]. Other specimens described from the Balcan area are Cooksonella sphaerica, Baragwanathia sp., Taeniocrada sp., and Jugumella burubaensis [102,103].
Figure 6.
Plant assemblage from the South Laurussian–Northwest Gondwanan unit (temperate zone) known from the Homerian of the Barrandian area, the Prague Basin (parvus-nassa Biozone); (a) Pachytheca showing differentiated medulla (inner) and cortex (outer par), scale bar = 500 µm; (b) Cortical fragment of Pachytheca in detail composed of densely spaced radial tubes, scale bar = 20 µm; (c) Non-embryophyte plant cf. Nematasketum showing unevenly thickened tubes, SEM-rack SILI-A1, scale bar = 200 µm; (d) Cf. Cosmochlaina showing outer covering layer being partly perforated, scale bar = 200 µm; (e) Varying diameter of tubes of Prototaxites, scale bar = 100 µm; (f) Detail of tubes of Prototaxites, scale bar = 20 µm. All specimens were observed under SEM.
Figure 6.
Plant assemblage from the South Laurussian–Northwest Gondwanan unit (temperate zone) known from the Homerian of the Barrandian area, the Prague Basin (parvus-nassa Biozone); (a) Pachytheca showing differentiated medulla (inner) and cortex (outer par), scale bar = 500 µm; (b) Cortical fragment of Pachytheca in detail composed of densely spaced radial tubes, scale bar = 20 µm; (c) Non-embryophyte plant cf. Nematasketum showing unevenly thickened tubes, SEM-rack SILI-A1, scale bar = 200 µm; (d) Cf. Cosmochlaina showing outer covering layer being partly perforated, scale bar = 200 µm; (e) Varying diameter of tubes of Prototaxites, scale bar = 100 µm; (f) Detail of tubes of Prototaxites, scale bar = 20 µm. All specimens were observed under SEM.

Figure 7.
Plant assemblage from the South Laurussian–Northwest Gondwanan unit (temperate zone) from Přídolí (S. parultimus-S. ultimus Biozone) of the Barrandian area, the Prague Basin, Czechia; (a) Cooksonia cf. hemisphaerica, specimen of NM—D-475, Kosov Quarry; (b) Baragwanathia brevifolia, specimen of CGS—KR 1, Karlštejn locality; (c) Tichavekia grandis, specimen of WBM—F21761a, Kosov Quarry; (d) Cf. Fusiformitheca sp., specimen of WBM—F21762, Kosov Quarry; (e) Aberlemnia bohemica, specimen of SMNH—JE-Sch0260B, Dlouhá Hora locality; (f) Cooksonia sp., specimen of FSCU, Kosov Quarry. All scales 10 mm.
Figure 7.
Plant assemblage from the South Laurussian–Northwest Gondwanan unit (temperate zone) from Přídolí (S. parultimus-S. ultimus Biozone) of the Barrandian area, the Prague Basin, Czechia; (a) Cooksonia cf. hemisphaerica, specimen of NM—D-475, Kosov Quarry; (b) Baragwanathia brevifolia, specimen of CGS—KR 1, Karlštejn locality; (c) Tichavekia grandis, specimen of WBM—F21761a, Kosov Quarry; (d) Cf. Fusiformitheca sp., specimen of WBM—F21762, Kosov Quarry; (e) Aberlemnia bohemica, specimen of SMNH—JE-Sch0260B, Dlouhá Hora locality; (f) Cooksonia sp., specimen of FSCU, Kosov Quarry. All scales 10 mm.

3.2.4. Northeast Gondwanan Unit
This palaeophytogeographical unit includes Northeast Gondwana (Figure 2) and was probably situated as a transition between the tropical-subtropical zones, between 10°–15°south latitude [76]. Similar vegetation is reported from Yunnan (Figure 2—Pd), Xinjiang (Figure 2—P12, Pe) (both China) [50,104], and Vietnam (Figure 2—P13) [105].
Rich Ludfordian assemblage flora were described from Victoria, Australia [42,92], situated in the tropical zone of eastern Gondwana (approximately 10° S) [76]. Two stratigraphical levels with plants were recognised, (1) Stratigraphically older Monograptus Beds with seven plant taxa (Figure 3—L5), and (2) the Late Centennial Beds with four taxa (Figure 3—L4). Rhyniophytes dominated and zosterophylls were common. The assemblage includes the enigmatic lycopsid Baragwanathia longifolia and an undescribed lycophyte-like endemic plant [42,111].
The Yunnan and Xinjiang provinces of China yielded a zosterophyll-dominated flora of Přídolí age (Figure 3—Pd, P12, Pe) with e.g., Zosterophyllum qujingense [50]. Dou and Sun [112] and Cai et al. [104] described Zosterophyllum sp., Sciadophyton pristinum, Salopella xinjiangensis, Cooksonella (Junggaria) sphaerica, Hostinella, Psilophytites, Parka and lycophyte-like plants from Xinjinang, north China. Gonez et al. [105] published a plant assemblage from the Van Canh Formation, Vietnam (Figure 3—P13), which may belong to Přídolí, probably bouceki—transgrendies biozones and mentioned unbranched axes, cf. Sporogonites yunnanense, cf. Aberlemnia, cf. Filiformorama, and zosterophylls. The locality is close to other Chinese Silurian assemblages situated approximately 20° south latitude [76] on the South China Plate.
3.3. Palynological Aspects of Early Land Plants
The combination of the dispersed spore and plant megafossil records is necessary for research into the history of land plants, including the early ones [7,113,114]. Reconstructions of palaeoenvironmental conditions within the Silurian are based mainly on the marine fossil record. Dispersed spores, however, also occurring mainly in near-shore marine sediments, can also be a significant independent factor for successful reconstructions of contemporaneous palaeoenvironments on adjacent palaeocontinents. Indirect palynological evidence of the plant record is available most commonly in the form of spores dispersed in the rock record, and only sometimes in situ, i.e., spores isolated directly from the reproductive organs of plants. The megafossil record of Silurian plants is limited and incomplete compared to that of their spores. Spores have a much higher preservational potential than other plant tissues due to their size and the occurrence of sporopollenin in their exines that effectively protects them against degradation and, thus, leads to better fossilization. There is a very low preservational potential for plants without mineralized mechanical tissues [115,116]. Therefore, the first evidence for land plants is in the form of dispersed spores with a resistant sporopollenin wall [117,118]. Two other palaeobotanically very important aspects of spores are, first, their production in enormous numbers by homosporous plants, and second, their great dispersal potential over long distances by water, but mainly by wind. The optimum size for wind dispersal is 25 μm or less according to Mogensen [119], which corresponds to the diameter of the majority of isospores produced by early land plants (usually 20–40 μm) [120,121].
Cryptospores sensu Stemans [122] appeared earlier than true trilete spores [16]. Their producers/parent plants “cryptosporophytes” [7] are a heterogeneous group of different plant types. It is possible to recognize several types of cryptospores, including enveloped enclosed monads, dyads, and tetrads, naked monads, dyads, and tetrads (Table 1). A large number of dispersed trilete spores in the fossil record reflect the strategy of the earliest plants, which were isosporic, resulting in efficient spreading. Additionally, their abundance in the fossil record significantly exceeds the abundance of the parent plant macrofossils [48].
Based on the dispersed spore record it seems that relatively uniform vegetation was present for 30 million years from the Daipingian to the Llandovery [123].
Wellman et al. [123] summarized the records of 787 spore species of all palaeocontinents from Ordovician to Late Silurian. The graphical interpretation of global quantitative data from Katian to Přídolí (Figure 8) shows that biodiversity curves of cryptospores and trilete isospores are different. It also suggests that cryptosporophytes and trilete spore producers had different life strategies and dispersal potential. A prominent cryptospore peak occurs within Rhuddanian (17 genera with 33 species on average), and after a significant decline in the Sheinwoodian (only 9 genera with 9 species) the occurrence from the Homerian to Přídolí is relatively stable (14 genera with 25 species on average).
Figure 8.
Quantitative curves of cryptospores (dashed line) and trilete spores (full line) from Katian to Přídolí. Horizontal axis represents stratigraphical interval. Abreviations: K—Katian, H—Hirnantian, LR—Llandovery (Rhuddanian), LA—Llandovery (Aeronian), LT—Llandovery (Telychian), WS—lower Wenlock (Sheinwoodian), WH—upper Wenlock (Homerian), LG—lower Ludlow (Gorstian), LL—upper Ludlow (Ludfordian), P—Přídolí. Vertical axis represents average number of trilete spores. Based on data of Wellman et al. [123].
Figure 8.
Quantitative curves of cryptospores (dashed line) and trilete spores (full line) from Katian to Přídolí. Horizontal axis represents stratigraphical interval. Abreviations: K—Katian, H—Hirnantian, LR—Llandovery (Rhuddanian), LA—Llandovery (Aeronian), LT—Llandovery (Telychian), WS—lower Wenlock (Sheinwoodian), WH—upper Wenlock (Homerian), LG—lower Ludlow (Gorstian), LL—upper Ludlow (Ludfordian), P—Přídolí. Vertical axis represents average number of trilete spores. Based on data of Wellman et al. [123].

The diversity of trilete spores was constant from the Hirnantian to the Sheinwoodian (only 3 genera with 4 species on average); the first global proliferation event of trilete spores, i.e., presumably diversifying ancestors of bryopphytes and tracheophytes [79,123,124] is within Homerian (13 genera with 29 species). After a short period with a decreased number of trilete spore taxa in the Gorstian, a prominent increasing trend is recognized from the Ludfordian (20 genera with 48 species) to the second globally recorded event within the Přídolí (35 genera with 129 species).
The Homerian event is documented mainly palynologically [79,123] but the Přídolí event is recognized both palynologically and by megafossil plant records [38].
Hagström and Mehlqvist [94,114,125] mentioned embryophyte cryptospores of probable Ludfordian Age from approximately 15° south latitude [76] (Figure 2 and Figure 3—L7) from Sweden, i.e., a former eastern part of Laurisia.
All events in the history of Silurian land plants documented here palynologically correspond to changes in the temperature and atmospheric pO2 and pCO2, as we will discuss.
4. Discussion
4.1. Response of Silurian Plants to Environmental Perturbations
4.1.1. Wenlock
The Wenlockian flora is known [47] only from the cold temperate zones (Figure 2) of South Laurasia-Northwest Gondwana. It is characterized by a rhyniophytoid-dominated plant assemblage (e.g., Figure 5). Polysporangiophytes of the Wenlock Age are mostly found within the temperate climatic zone to subpolar areas, where significant seasonality can be expected. Generally, the Wenlock is characterized by colder climate culminating in the late Homerian glaciation (Figure 1 and Figure 3). A high diversity of plant types is indicated by the occurrence of dispersed spore assemblages from localities in the Barrandian area (the Prague Basin). For example, the analysis of a Homerian sample coming from a single bed revealed at least ten to twelve types of trilete spore producing plants and three to four types of cryptosporophytes as early as in the Middle Sheinwoodian [78,79]. This fact evoked an idea that relatively rich terrestrial plant assemblages evolved continuously through the Wenlock, through at least two graptolite biozones, in the volcanic islands of the Barrandian area (the Prague Basin), the Czech Republic. Generally, this assemblage (Figure 3—W1 plus Wa) characterized relatively rich vegetation that probably covered physically suitable areas of the Southern hemisphere. Some bryophyte-like microscopic tissue fragments are preserved abundantly within the Homerian (e.g., Figure 6).
The Homerian glaciation event probably created new large-scale habitats in the physical space that was created by the regression accompanying ice formation. These new habitats were certainly colonized by plants. Plants sensitively responded to seasonal temperature fluctuations and humidity as mentioned by Raymond et al. [47]. Nevertheless, the plants from the temperate zone probably did not have to face the frost during the winter, as suggested by Raymond et al. [47], despite the fact that the Wenlock was a cold period certainly limiting the possibility of plant growth and dispersal (Figure 1). This seems to be supported palynologically because Sheinwoodian cryptospore and trilete spore assemblages reach their minimum diversity here. The first global key proliferation event for trilete spore-producing land plants is documented palynologically after Homerian glaciation (Figure 3). It seems that the Wenlock can be palaeobotanically and palynologically divided into a less environmentally favorable Sheinwoodian interval and a more environmentally suitable late Homerian interval for early land plants.
Edwards and Richardson [46] proposed a ruderal species strategy for rhyniophytoid plants, including a reproductive cycle that was completed during the times of high humidity. Plants persisted as latent spores during periods of unfavorable conditions, although still in a constantly humid environment (mesophytic habitats) [46]. Unfortunately, we do not have evidence for tropical and subtropical plants from Wenlock time.
4.1.2. Ludlow
Since Early Ludlow, the global temperature was increasing until the Middle Ludfordian when this trend was interrupted by a rapid cooling during the Mid-Ludfordian glaciation (Figure 3). After the glaciation, the global temperature reached the same level as before the glaciation.
Unique Ludlow data come from the equatorial zone where well-preserved plant assemblages from Arctic Canada (Figure 4) and Northern Greenland are reported. This vegetation, characterized by dominance of zosterophylls, has been referred to the North Laurussian palaeophytogeographic unit [47]. Similar plant assemblages of Ludlow age are known from the Northeast Gondwanan (Australia) unit (sensu [47]) where the climate has a transitional character between the tropical and subtropical zones. This area was located approximately 10 ° south latitude during Late Silurian times [76]. Raymond et al. [47] stated that flora from Australia cannot be compared with zosterophyll-dominated vegetation from Bathurst Island of Arctic Canada because different taxa occurred in the two areas, indicating an evolutionary disjunction.
North Laurussian and northeast Gondwanan units are characterized by the predominance of zosterophylls due to the warm climate (sensu [126]). Zosterophyll-dominated assemblages from northeast Gondwana also yielded a number of rhyniophytes. It is likely that these areas represent the cradle of zosterophyll origin based on their absence in the lower Silurian and on their sudden abundance during Late Silurian in areas where they became the dominant element [50]. So far, we can only speculate why in this warm zone plants appear that have sporangia in the terminal part of their shoots. Certainly, a significant factor was the relatively constant and high temperature and humidity throughout the year, when the growing season was not interrupted by any significant temperature or water stresses. As a basal group of lycophytes, they could have had very similar ecological requirements as the later lycophytes-from the Carboniferous Age. These younger plants preferred a very humid climate and were a significant part of most peat forming assemblages. Based on the absence of zosterophylls in the Early Silurian and the sudden increase in abundance during the Late Silurian in areas where they became the dominant element of the flora, it is likely that these areas also represent the cradle of their origin [50]. In any case it could be assumed that both rhyniophytes and zosterophyllophytes in this period were already equipped with a primitive type of stomata [127].
Raymond et al. [47] interpreted assemblages from Australia and Bathurst Island as products of warm zones and mentioned the same ecological conditions accompanying the origin of zosterophyll-like plants. However, it can be concluded that the overall warm climate with episodes of the expansion to higher latitudes (after glacials) of the humid tropical environment during the Silurian undoubtedly led to the evolution of this lycophyte type of plant. Generally, based on palynological results, Gorstian spore and cryptospore assemblages are less numerous and less diverse than previous Homerian associations, which was probably influenced by the preceding Late Homerian glaciation.
4.1.3. The Importance of Mid-Ludfordian Glaciation
The Mid-Ludfordian has been known for one of the largest perturbations in the Phanerozoic carbon cycle, the Mid-Ludfordian Carbon Isotope Excursion, linked with an extinction event recorded in global marine ecosystem. Significant global cooling was recently documented from temperate areas of the Prague Basin and Carnic Alps (peri-Gondwana), as well as from the tropical areas of Baltica (Laurussia) and Australia (Gondwana) based on δ18O data [24]. The marked cooling of sea-surface temperatures, coupled with a significant eustatic sea-level fall recorded on all corresponding palaeocontinents, evidences a major glaciation (“Mid-Ludfordian Glaciation”) in polar and subpolar western Gondwana.
Glaciation passed during Late Ludlow (Ludfordian) accompanied by a significant reduction of average temperature (Figure 1). Large-scale dry land habitat space was created by the Mid-Ludfordian glaciation event and associated sea-level drop, certainly accompanied by plant colonization of these new areas following oceanic regression. We suppose that the vacant ecological windows were occupied by trilete spore producing embryophytes together with bacterial-cyanobacterial mats and still some cryptosporophytes, which colonized coastal areas. Plants established complex and specialized ecosystems on the new land habitats that served as a nutrient source for the rapid global expansion of polysporangiophyte plants, supported also by distinct climatic changes. The expansion is especially apparent for plants preferring warmer and more humid conditions. Raymond et al. [47] supposed that the lycophyte genus Baragwanathia is typical for warm and humid climatic zones. However, Baragwanathia brevifolia and another basal lycophyte, Aberlemnia bohemica, are described also from a higher latitude of the peri-Gondwana Realm (namely Barrandian area, the Prague Basin, Czech Republic) and a higher stratigraphical level (Přídolí) [38,43]. It seems that the expansion of Baragwanathia-like plants to higher latitudes is associated with increased humidity during glaciation and following global warming after the Mid-Ludfordian glacial.
A similar shift to higher latitudes of lycophytes and zosterophylls can be traced in other localities that belong to the temperate zone of the South Laurussian-Northwest Gondwanan unit [47]. For example, a Podolian assemblage (Figure 2—P8) from eastern Laurussia shows dominance of rhyniophytoids, including Cooksonia pertoni, C. hemisphaerica, and mainly Zosterophyllum sp. and Lycopodolica, rather typical for the warmer equatorial Silurian zones. The Ludfortian is palynologically characterized by the increasing variability of spore and cryptospore assemblages with a high number of new taxa.
4.1.4. Přídolí
The prominent rise in temperature during the Přídolí [128] was permissive of the evolution of new forms of plants. Transgression contributed to the frequent transport of land plants into the sea and created Silurian taphonomic windows.
The number of Silurian localities with rhyniophytoid-dominated vegetation (including the typical occurrence of Cooksonia) is higher during the Přídolí than during the Ludlow. This homogenization was based by Raymond et al. [47] on floras of similar composition in both areas where the rhyniophytoid-dominated flora is present and includes some common taxa, e.g., Cooksonia hemisphaerica. In this context however, the occurrence of C. hemisphaerica s.s. in Northwest Gondwana is still questionable and needs re-evaluation. A similar situation is also found with respect to a comparison of the character of assemblages from Great Britain and Bolivia [47]. Raymond et al. [47] explained this similarity by the seasonality that equally affected plants from SE Laurussia in the subtropical zone and Northwest Gondwana or peri-Gondwana in the temperate zone.
The general plant character from China and Vietnam shows some similarities [105,126] to Ludlowian vegetation from tropical Bathurst Island or maybe subtropical Victoria, Australia, in having Zosterophyllum-like plants as dominant. Plants of Přídolí age are known from the Kazakhstanian unit situated in the subtropical zone of the North hemisphere. Northern latitude floras differ from those of environmentally similar southern latitudes in the common occurrence of Zosterophyllum-like plants in the former. Edwards and Wellman [45] pointed out this phenomenon and placed the Kazakhstanian flora on the South China Block (Xinjiang flora). Relatively rich localities already appear in several areas during the Přídolí and this trend continues across the Silurian/Devonian boundary into the Early Devonian. The number and variations of spore and cryptospore assemblages rapidly increased during the Přídolí Series, reflecting the second globally important key event for land plants.
5. Conclusions
Atmospheric oxygen steadily rose from the Middle Ordovician and through almost the entire Silurian. By the beginning of the Wenlock, oxygen levels surpassed the current level for the first time in the Earth’s history, and at the end of Silurian attained 35% [64,129]. This trend supports previous conclusions [49,64] and our own herein (based on our data and compiled fossil data as discussed) that, despite climate fluctuations, terrestrial flora expanded during the Silurian. We hope we have relaxed at least a small bit of constraints on Silurian phytogeography and support the conclusions of Edwards [49]. Current phylo-genomic/-stratigraphy dating of the embryophyte split into tracheophytes and bryophytes ranges from approximately 440 up to 800 Myr for tracheophytes [15,18]. This on one hand indicates uncertainty related to the timing of this major evolutionary cross-roads in land plant evolution. On the other hand, it also indicates that the tracheophyte lineage was certainly well established in the Silurian. Our analysis of spore diversity evolution clearly indicates a very distinct exponential evolutionary acceleration of trilete spore producers starting in the lower Wenlock (and continuing in fact until today—Figure 8). This expansion of trilete spore producers is also reflected in an apparent geographical spread of documented macrofossils between the Wenlock and Ludlow/Přídolí (Figure 2). Cryptospore-producers, which dominated Silurian flora until the upper Llandovery stagnated evolutionarily until their Devonian extinction. In a more detailed attempt to correlate trilete spore diversity and global temperature changes (Figure 3), we conclude that after three distinct Middle to Late Silurian glaciations we can clearly see an increase of diversity correlated with the rise of the global temperature after the glaciations ended. Interestingly, while Homerian glaciation did not affect the diversity of cryptospores, it resulted in the distinct decline in the diversity of trilete spores. This indicates that there was a different sensitivity of cryptosporophytes vs. trilete spores-producers to the global cold temperature. In respect to fragmentary Silurian phytogeography data summarized vividly by Edwards [49] (see above), our data essentially support her conclusions that the four distinct plant assemblages correlate very well with the four palaeocontinents.
It certainly is very difficult even to speculate about the physiological character of Silurian plants. However, both detailed, enhanced anatomical knowledge of the immediately following Early Devonian rhyniophyte flora, and also advancements in phylogenomic and phylostratigraphic analyses provide important facts allowing an informed reconstruction of at least some aspects of the biology of Silurian trilete spore-producing plants (we do not speculate here about the biology of cryptosporophytes.) Based on all the current available fossil data, the Silurian flora is composed of different types of polysporangiate plants (including cryptophytes)—any reliable evidence for the monosporangiates is absent. There are no identified gametophytes of any Silurian plant. Thus, the first reliable life cycles/alteration of gametophyte and sporophyte generations is not known before that of several representatives of Devonian Rhynie chert flora [130]. In relation to plant physiology, stomata are one of the crucial ecophysiological features related to both temperature, water availability and atmospheric composition. Based on both phylogenomics-indicating reduction or loss of stomatal functioning in the common ancestor of extant embryophytes in bryophytes [127], as well as the palaeobotanical demonstration of distinct stomata described both on sporophytes and gametophytes of Rhynie chert plants [131], we should accept that the common ancestor of both mosses and tracheophytes had stomata—most probably already dynamically regulated (opening-closing) by the abscisic acid signalling pathway [127,132]. This pathway is crucial for the optimization of land-plant water economy and survival of water stresses both during day-night cycles, but importantly also during seasons allowing local adaptations dependent on latitude. As is the case for abscisic acid signalling, the evolution of lignified secondary-thickened conductive tissues is well documented in Rhynie chert plants. Lignification most probably started much earlier and precursors of tracheophyte xylem and phloem were with high probability also present in the common Bryophyta-Tracheophyta ancestor. As in the case of stomata [127], it is possible that secondary simplification accompanied the evolution of other traits in the Bryophyta. Major transcriptional regulators of xylem development are also encoded in bryophyte genomes and VND-NST-SOMBRERO (VNS) genes—vessel-element differentiation master regulators—from Sphagnum moss (which does not have hydroids and instead uses hyaline cells with thickened, helical-patterned cell walls and pores to store water in the leaves) and induces secondary cell wall thickening when ectopically expressed in the angiosperm Nicotiana benthamiana [133]. The anatomy of Silurian (and Devonian) plant fossils/assemblages, along with recent phylogenomic analyses, clearly indicate highly terrestrialized advanced plant types with likely a great ability to resist different dry land stress conditions and to evolve on different palaeocontinets into quite disjunctive specific floras/plant assemblages certainly also in response to different geological, geographical and climatic conditions.
Author Contributions
Conceptualization, J.P., V.Ž. and J.B.; methodology, J.P., J.B. and J.F.; formal analysis, J.P., J.B. and M.U.; investigation, J.P., J.B., J.F., M.U., V.Ž. and P.Š.; resources, J.P.; data curation, J.P., J.B., J.F., M.U. and P.Š.; writing—original draft preparation, J.P.; writing—review and editing, J.P., J.B., J.F. and V.Ž.; visualization, J.P.; supervision, V.Ž.; project administration, J.P.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was financial supported by the Czech Science Foundation—GAČR 21-10799S. Part of this contribution was also supported by the Research Program of the Institute of Geology AS CR, v.v.i. (RVO67985831) and West Bohemian Museum in Pilsen DKRVOZCM2020-25/93P.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank to the authors of Kotyk et al. [74] who provided us with photos of the specimen from Bathurst Island through Patricia Gensel. We also thank the National Museum (Prague), West Bohemian Museum in Pilsen, and private collector Ondřej Zicha, who provided us with photographs of specimens from the Silurian of the Barrandien. The SEM observations were made in the Institute of Geology and Palaeontology, Faculty of Science, Charles University in Prague from examination under JEOL 6380LV. We would like to thank Bill DiMichele for comments and corrections in the use of the English language. Last but not least, the first author of this contribution thanks to Francis Beer Café in Pilsen for the amazing atmosphere that helped originate of this manuscript.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Wellman, C.H.; Strother, P.K. The terrestrial biota prior to the origin of land plants (embryophytes): A review of the evidence. Palaeontology 2015, 58, 601–627. [Google Scholar] [CrossRef]
- Kenrick, P. Changing expressions: A hypothesis for the origin of the vascular plant life cycle. Philos. Trans. R. Soc. B 2017, 373, 20170149. [Google Scholar] [CrossRef]
- Cooper, E.D.; Henwood, M.J.; Brown, E.A. Are the liverworts really that old? Cretaceous origins and Cenozoic diversifications in Lepidoziaceae reflect a recurrent theme in liverwort evolution. Biol. J. Linn. Soc. 2012, 107, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Zhong, B.; Fong, R.; Collins, L.J.; McLenachan, P.A.; Penny, D. Two new fern chloroplasts and decelerated evolution linked to the long generation time in tree ferns. Genome Biol. Evol. 2014, 6, 1166–1173. [Google Scholar] [CrossRef] [Green Version]
- Willis, K.J.; McElwain, J.C. The Evolution of Plants; Oxford University Press: Oxford, UK, 2002; p. 378. [Google Scholar]
- Taylor, W.A.; Wellman, C.H. Ultrastructure of enigmatic phytoclasts (banded tubes) from the Silurian-lower Devonian: Evidence for affinities and role in early terrestrial ecosystems. Palaios 2009, 24, 167–180. [Google Scholar] [CrossRef]
- Edwards, D.; Morris, J.L.; Richardson, J.B.; Kenrick, P. Cryptospores and cryptophytes reveal hidden diversity in early land floras. New Phytol. 2014, 202, 50–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, D.; Kenrick, P. The early evolution of land plants, from fossils to genomics: A commentary on Lang (1937) ‘On the plant-remains from the Downtonian of England and Wales’. Philos. Trans. R. Soc. B 2015, 370, 20140343. [Google Scholar] [CrossRef] [Green Version]
- Tomescu, A.M.F.; Rothwell, G.W. Wetlands before tracheophytes: Thalloid terrestrial communities of the Early Silurian Passage Creek biota (Virginia). Geol. Soc. Am. Spec. Pap. 2006, 399, 41–56. [Google Scholar]
- Tomescu, A.M.F.; Honegger, R.; Rothwell, G.W. Earliest fossil record of bacterial–cyanobacterial mat consortia: The early Silurian Passage Creek biota (440 Ma, VA, USA). Geobiology 2008, 6, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Tomescu, A.M.F.; Tate, R.W.; Mack, N.G.; Calder, V.J. Simulating fossilization to resolve the taxonomic affinities of thalloid fossils in Early Silurian (ca. 425 Ma) terrestrial assemblages. Bibl. Lichenol. 2010, 105, 183–189. [Google Scholar]
- Rubinstein, C.V.; Gerrienne, P.; de la Puente, G.S.; Astini, R.A.; Steemans, P. Early middle Ordovician evidence for land plants in Argentina (eastern Gondwana). New Phytol. 2010, 188, 365–369. [Google Scholar] [CrossRef]
- Salamon, S.A.; Gerrienne, P.; Steemans, P.; Gorzelak, P.; Filipiak, P.; Le Hérissé, A.; Paris, F.; Cascales-Minana, B.; Brachaniec, T.; Misz-Kennan, M.; et al. Putative late Ordovician land plants. New Phytol. 2018, 218, 1305–1309. [Google Scholar] [CrossRef] [Green Version]
- Naugolnykh, S.V. Plants of the First Terrestrial Ecosystems. Her. Russ. Acad. Sci. 2019, 89, 502–511. [Google Scholar] [CrossRef]
- Morris, J.L.; Puttick, M.N.; Clark, J.W.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Yang, Z.; Schneider, H.; Donoghue, P.C.J. The timescale of early land plant evolution. Proc. Natl. Acad. Sci. USA 2018, 115, E2274–E2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steemans, P.; Le Hérissé, A.; Melvin, J.; Miller, M.A.; Paris, F.; Verniers, J.; Wellman, C.H. Origin and Radiation of the Earliest Vascular Land Plants. Science 2009, 324, 353. [Google Scholar] [CrossRef] [Green Version]
- Strother, P.K.; Beck, J. Spore–like microfossils from Middle Cambrian strata: Expanding the meaning of the term cryptospore, 413–424. In Pollen and Spores: Morphology and Biology; Harley, M.M., Morton, C.M., Blackmore, S., Eds.; Royal Botanic Gardens, Kew: Richmond, UK, 2000. [Google Scholar]
- Su, D.; Yang, L.; Shi, X.; Ma, X.; Zhou, X.; Hedges, S.B.; Zhong, B. Large-Scale Phylogenomic Analyses Reveal the Monophyly of Bryophytes and Neoproterozoic Origin of Land Plants. Mol. Biol. Evol. 2021, 1–13. [Google Scholar] [CrossRef]
- Schopf, J.M.; Mencher, E.; Boucot, A.J.; Andrews, H.N. Erect plants in the early Silurian of Maine. US Geol. Sur. Rest. VA Prof. Pap. 1966, 550-D, D69–D75. [Google Scholar]
- McAdams, N.E.B.; Cramer, B.D.; Bancroft, A.M.; Melchin, M.J.; Devera, J.A.; Day, J.E. Integrated δ13Ccarb, conodont, and graptolite biochemostratigraphy of the Silurian from the Illinois Basin and stratigraphic revision of the Bainbridge Group. GSA Bull. 2018, 131, 335–352. [Google Scholar] [CrossRef]
- Cramer, B.D.; Brett, C.E.; Melchin, M.J.; Männik, P.; Kleffner, M.A.; McLaughlin, P.I.; Lloydell, D.K.; Munnecke, A.; Jeppsson, L.; Corradini, C.; et al. Revised correlation of Silurian provincial series of North America with global and regional chronostratigraphic and δ13Ccarb chemostratigraphy. Lethaia 2011, 44, 185–202. [Google Scholar] [CrossRef]
- Melchin, M.J.; Sadler, P.M.; Cramer, B.D. Chapter 21—The Silurian period. In The Geological Time Scale; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2020; pp. 695–732. [Google Scholar]
- Grossman, E.L.; Joachimski, M.M. Oxygen isotope stratigraphy. In The Geological Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2020; pp. 279–307. [Google Scholar]
- Frýda, J.; Lehnert, O.; Joachimski, M.; Mannik, P.; Kubajko, M.; Mergl, M.; Farkaš, J.; Frýdová, B. The Mid-Ludfordian (late Silurian) Glaciation: A link with global changes in ocean chemistry and ecosystem overturns. Earth Sci. Rev. 2021, 220, 103652. [Google Scholar] [CrossRef]
- Gerrienne, P.; Servais, T.; Vecoli, M. Plant evolution and terrestrialization during Palaeozoic times—The phylogenetic context. Rev. Palaeobot. Palynol. 2016, 227, 4–18. [Google Scholar] [CrossRef]
- Edwards, D.; Axe, L.; Honeggert, R. Contributions to the diversity in cryptogamic covers in the mid-Palaeozoic: Nematothallus revisited. Bot. J. Linn. Soc. 2013, 173, 505–534. [Google Scholar] [CrossRef]
- Abbott, G.D.; Ewbank, G.; Edwards, D.; Wang, G.Y. Molecular characterization of some enigmatic Lower Devonian fossils. Geochim. Cosmochim. Acta 1998, 62, 1407–1418. [Google Scholar] [CrossRef]
- Edwards, D.; Axe, L. Evidence for a fungal affinity for Nematasketum, a closeally of Prototaxites. Bot. J. Linn. Soc. 2012, 168, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.Q.; Edwards, D.; Wu, P.C.; Duckett, J.G.; Hueber, F.M.; Li, C.S. Riccardiothallus devonicus gen. et sp. nov., the earliest simple thalloid liverwort from the Lower Devonian of Yunnan, China. Rev. Palaeobot. Palynol. 2012, 176–177, 35–40. [Google Scholar] [CrossRef]
- Strother, P.K.; Al-Hajri, S.; Traverse, A. New evidence for land plants from the lower Middle Ordovician of Saudi Arabia. Geology 1996, 24, 55–59. [Google Scholar] [CrossRef]
- Cox, C.J.; Li, B.; Foster, P.G.; Embley, T.M.; Civáň, P. Conflicting Phylogenies for Early Land Plants are Caused by Composition Biases among Synonymous Substitutions. Syst. Biol. 2014, 63, 272–279. [Google Scholar] [CrossRef]
- Kenrick, P.; Crane, P.R. Water-Conducting Cells in Early Fossil Land Plants: Implications for the Early Evolution of Tracheophytes. Bot. Gaz. 1991, 152, 335–356. [Google Scholar] [CrossRef]
- Cascales-Miñana, B.; Steemans, P.; Servais, T.; Lepot, K.; Gerrienne, P. An alternative model for the earliest evolution of vascular plants. Lethaia 2019, 52, 445–453. [Google Scholar] [CrossRef]
- Edwards, D. Embryophytic sporophytes in the Rhynie and Windyfield cherts. Trans. R. Soc. Edinb. Earth Sci. 2004, 94, 397–410. [Google Scholar] [CrossRef]
- Wellman, C.H. The invasion of the land by plants: When and where? New Phytol. 2010, 188, 306–309. [Google Scholar] [CrossRef]
- Graham, L.; Lewis, L.A.; Taylor, W.; Wellman, C.H.; Cook, M. Early terrestrialization: Transition from algal to bryophyte grade. In Photosynthesis in Bryophytes and Early Land Plants; Hanson, D.T., Rice, S.K., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 9–28. [Google Scholar]
- Harrison, C.J. Development and genetics in the evolution of land plant body plans. Philos. Trans. R. Soc. B 2017, 372, 20150490. [Google Scholar] [CrossRef] [Green Version]
- Kraft, P.; Pšenička, J.; Sakala, J.; Frýda, J. Initial plant diversification and dispersal event in upper Silurian of the Prague Basin. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 514, 144–155. [Google Scholar] [CrossRef]
- Kenrick, P.; Crane, P.R. The Origin and Early Diversification of Land Plants. Nature 1997, 389, 33–39. [Google Scholar] [CrossRef]
- Edwards, D. New insights into early land ecosystems: A glimpse of a Lilliputian world. Rev. Palaeobot. Palynol. 1996, 90, 159–174. [Google Scholar] [CrossRef]
- Gonez, P.; Gerrienne, P. Aberlemnia caledonica gen. et comb. nov., a new name for Cooksonia caledonica Edwards 1970. Rev. Palaeobot. Palynol. 2010, 163, 64–72. [Google Scholar] [CrossRef]
- Tims, J.D.J.; Chambers, T.C. Rhyniophytina and Trimerophytina from the early land flora of Victoria, Australia. Palaeontology 1984, 27, 265–279. [Google Scholar]
- Kraft, P.; Kvaček, Z. Where the lycophytes come from?—A piece of the story from the Silurian of peri-Gondwana. Gondwana Res. 2017, 45, 180–190. [Google Scholar] [CrossRef]
- Rickards, R.B. The age of the earliest club mosses: The Silurian Baragwanathia flora in Victoria, Australia. Geol. Mag. 2000, 137, 207–209. [Google Scholar] [CrossRef]
- Edwards, D.; Wellman, C.H. Embryophytes on Land: The Ordovician to Lochkovian (Lower Devonian) Record. In Plants Invade the land, Evolutionary and Environmental Perspectives; Gensel, P.G., Edwards, D., Eds.; Columbia University Press: New York, NY, USA, 2001; pp. 3–28. [Google Scholar]
- Edwards, D.; Richardson, J.B. Silurian and Lower Devonian plant assemblages from the Anglo-Welsh Basin: A palaeobotanical and palynological synthesis. Geol. J. 2004, 39, 375–402. [Google Scholar] [CrossRef]
- Raymond, A.; Gensel, P.; Stein, W.E. Phytogeography of Late Silurian macrofloras. Rev. Palaeobot. Palynol. 2006, 142, 165–192. [Google Scholar] [CrossRef]
- Wellman, C.H.; Steemans, P.; Vecoli, M. Paleophytogeography of Ordovician–Silurian land plants. In Early Palaeozoic Biogeography and Palaeogeography; Harper, D.T., Servais, T., Eds.; Geological Society, London, Memoirs: London, UK, 2013; Volume 38, pp. 461–476. [Google Scholar]
- Edwards, D. Constraints on Silurian and Early Devonian phytogeographic analysis based on megafossils. In Palaeozoic Palaeogeography and Biogeography; McKerrow, W.S., Scotese, C.R., Eds.; Geological Society, London, Memoirs: London, UK, 1990; pp. 233–242. [Google Scholar]
- Hao, S.; Xue, J.; Liu, Z.; Wang, D. Zosterophyllum penhallow around the Silurian-Devonian Boundary of Northeastern Yunnan, China. Int. J. Plant Sci. 2007, 168, 477–489. [Google Scholar] [CrossRef]
- Bateman, R.M.; Crane, P.R.; DiMichele, W.A.; Kenrick, P.R.; Rowe, N.P.; Speck, T.; Stein, W.E. Early evolution of land plants: Phylogeny, physiology, and ecology of the primary terrestrial radiation. Annul. Rev. Ecol. Syst. 1998, 29, 263–292. [Google Scholar] [CrossRef] [Green Version]
- Servais, T.; Cascales-Miñana, B.; Cleal, C.J.; Gerrienne, P.; Harper, D.A.T.; Neumann, M. Revisiting the Great Ordovician Diversification of land plants: Recent data and perspectives. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 534, 109280. [Google Scholar] [CrossRef]
- Servais, T.; Martin, R.E.; Nützel, A. The impact of the ‘terrestrialisation process’ in the late Palaeozoic: pCO2, pO2, and the ‘phytoplankton blackout’. Rev. Palaeobot. Palynol. 2016, 224, 26–37. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Boyce, C.K. Carbon sources for the Palaeozoic giant fungus Prototaxites inferred from modern analogues. Proc. R. Soc. B 2010, 277, 2149–2156. [Google Scholar] [CrossRef] [Green Version]
- Kenrick, P.; Wellman, C.H.; Schneider, H.; Edgecombe, G.D. A timeline for terrestrialization: Consequences for the carbon cycle in the Palaeozoic. Philos. Trans. R. Soc. London Ser. B 2012, 367, 519–536. [Google Scholar] [CrossRef] [PubMed]
- Trotter, J.A.; Williams, I.S.; Barnes, C.R.; Männik, P.; Simpson, A. New conodont δ18O records of Silurian climate change: Implications for environmental and biological events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 443, 34–48. [Google Scholar] [CrossRef]
- Gambacorta, G.; Menichetti, E.; Trincianti, E.; Torricelli, S. The Silurian climatic transition recorded in the epicontinental Baltica Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 517, 16–29. [Google Scholar] [CrossRef]
- Signor, P.W.; Brett, C.E. The mid−Paleozoic precursor to the Mesozoic marine revolution. Paleobiology 1984, 10, 229–245. [Google Scholar] [CrossRef]
- Brett, C.E.; Walker, S.E. Predators and predation in Paleozoic marine environments. Paleontol. Soc. Pap. 2002, 8, 93–118. [Google Scholar] [CrossRef]
- Nützel, A.; Frýda, J. Palaeozoic plankton revolution: Evidence from early gastropod ontogeny. Geology 2003, 31, 829–831. [Google Scholar] [CrossRef]
- Klug, C.; Kroger, B.; Kiessling, W.; Mullins, G.L.; Servais, T.; Frýda, J.; Korn, D.; Turner, S. The Devonian nekton revolution. Lethaia 2010, 43, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Berner, R.A. Phanerozoic atmospheric oxygen: New results using the GEOCARBSULF model. Am. J. Sci. 2009, 309, 603–606. [Google Scholar] [CrossRef]
- Berner, R.A.; Beerling, D.J.; Dudley, R.; Robinson, J.M.; Wildman, R.A. Phanerozoic atmospheric oxygen. Annu. Rev. Earth Planet. Sci. 2003, 31, 105–134. [Google Scholar] [CrossRef] [Green Version]
- Schachat, S.R.; Labandeira, C.C.; Saltzman, M.R.; Cramer, B.D.; Payne, J.L.; Boyce, C.K. Phanerozoic pO2 and the early evolution of terrestrial animals. Proc. R. Soc. B 2018, 285, 20172631. [Google Scholar] [CrossRef] [Green Version]
- Saltzman, M.R.; Thomas, E. Carbon isotope stratigraphy. In The Geologic Time Scale 2012; Gradstein, F., Ogg, J., Schmitz, M.D., Ogg, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 207–232. [Google Scholar] [CrossRef]
- Munnecke, A.; Samtleben, C.; Bickert, T. The Ireviken event in the Lower Silurian of Gotland, Sweden—Relation to similar Palaeozoic and Proterozoic events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 195, 99–124. [Google Scholar] [CrossRef]
- Loydell, D.K. Reply to ‘Early Silurian positive δ13C excursions and their relationship to glaciations, sea-level changes and extinction events: Discussion’ by Bradley, D. Cramer and Axel Munnecke. Geol. J. 2008, 43, 511–515. [Google Scholar] [CrossRef]
- Calner, M. Silurian global events—At the tipping point of climate change. In Mass Extinctions; Ashraf, M.T., Ed.; Springer–Verlag: Berlin/Heidelberg, Germany, 2008; pp. 21–58. [Google Scholar]
- Lehnert, O.; Mannik, P.; Joachimski, M.M.; Calner, M.; Frýda, J. Palaeoclimate perturbations before the Sheinwoodian glaciation: A trigger for extinctions during the ‘Ireviken Event’. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 296, 320–331. [Google Scholar] [CrossRef]
- Veizer, J.; Ala, D.; Azmy, K.; Bruckschen, P.; Buhl, D.; Bruhn, F.; Carden, G.A.F.; Diener, A.; Ebneth, S.; Godderis, Y.; et al. 87Sr/86Sr, δ13C and δ18O evolution of Phanerozoic seawater. Chem. Geol. 1999, 161, 59–88. [Google Scholar] [CrossRef] [Green Version]
- Veizer, J.; Prokoph, A. Temperatures and oxygen isotopic composition of Phanerozoic oceans. Earth Sci. Rev. 2015, 146, 92–104. [Google Scholar] [CrossRef]
- Lécuyer, C.; Allemand, P. Modelling of the oxygen isotope evolution of seawater: Implication for the climate interpretation of the δ18O of marine sediments. Geochim. Cosmochim. Acta 1999, 63, 351–361. [Google Scholar] [CrossRef]
- Azmy, K.; Veizer, J.; Bassett, M.G.; Copper, P. Oxygen and carbon isotopic composition of Silurian brachiopods: Implications for coeval seawater and glaciations. Geol. Soc. Am. Bull. 1998, 100, 1499–1512. [Google Scholar] [CrossRef]
- Kotyk, M.E.; Basinger, J.F.; Gensel, P.G.; de Freitas, T.A. Morphologically complex plant macrofossils from the late Silurian of Arctic Canada. Am. J. Bot. 2002, 89, 1004–1013. [Google Scholar] [CrossRef]
- Larsen, P.-H.; Edwards, D.; Escher, J.C. Late Silurian plant megafossils from the Peary Land Group, North Greenland. Rapp. Gronlands Geol. Undersøgelse 1987, 133, 17–112. [Google Scholar]
- Scotese, C.R. Atlas of Silurian and Middle-Late Ordovician Paleogeographic Maps (Mollweide Projection), Maps 73–80, Volumes 5, The Early Paleozoic, PALEOMAP Atlas for ArcGIS; PALEOMAP Project: Evanston, IL, USA, 2014. [Google Scholar]
- Libertín, M.; Kvaček, J.; Bek, J.; Žárský, V.; Štorch, P. Sporophytes of polysporangiate land plants from the early Silurian period may have been photosynthetically autonomous. Nat. Plants 2018, 4, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Libertín, M.; Kvaček, J.; Bek, J. Plant diversity of the mid Silurian (lower Wenlock, Sheinwoodian) terrestrial vegetation preserved in marine sediments from the Barrandian area, the Czech Republic. Foss. Impr. 2018, 74, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Bek, J.; Štorch, P.; Tonarová, P.; Libertín, M. Early Silurian (mid-Sheinwoodian) palynomorphs from the Prague Basin, Czech Republic. Bull. Geosci. 2021. under review. [Google Scholar]
- Zicha, O. (Jugoslávských Partyzánů, Prague, Czech Republic). Personal communication, 2021. [Google Scholar]
- Honegger, R.; Edwards, D.; Axe, L.; Strullu-Derriens, C. Fertile Prototaxites taiti: A basal ascomycete with inoperculate, polysporous asci lacking croziers. Phil. Trans. R. Soc. B 2017, 373, 20170146. [Google Scholar] [CrossRef] [Green Version]
- Kříž, J. Silurian Field Excursions. Prague Basin (Barrandian), Bohemia; Geological Series of National Museum of Wales; National Museum of Wales: Cardiff, UK, 1992; Volume 13, pp. 1–111. [Google Scholar]
- Libertín, M.; Labuťa, R.; Dašková, J. The oldest vascular plants from the Bohemian Massif. Zprávy Geol. Výzkumech Roce 2002, 2002, 127. (In Czech) [Google Scholar]
- Horný, R. Studie o vrstvách budňanských v západní části barrandienského siluru. Sborník Ústředního Ust. Geol. 1955, 21, 315–448. [Google Scholar]
- Horný, R. Stratigraphy and tectonics of the western closures of the Silurian-Devonian synclinorium in the Barrandian area. Sborník Ústředního Ust. Geol. 1960, 26, 495–524. [Google Scholar]
- Obrhel, J. Die Flora der Pridoli-Schichten (Budnany-Stufe) des mittelböhmishcen Silurs. Geologie 1962, 11, 83–97. [Google Scholar]
- Schweitzer, H.J. Die Gattungen Renalia Gensel und Psilophyton Dawson im Unterdevon des Rheinlandes. Bonn. Palaeobot. Mitt. 1980, 6, 1–34. [Google Scholar]
- Kříž, J.; Jaeger, H.; Paris, F.; Schönlaub, H.P. Přídolí—The Fourth Subdivison of the Silurian. Jahrb. Geol. Bundesanst. 1986, 129, 291–360. [Google Scholar]
- Toro, M.; Chamon, O.; Salguero, R.; Vargas, C. Las plantas de la Formacion Kirusillas (Silurico) en la region de La Angostura Departamento de Cochabamba. In Memorias del XII Congreso Geológico de Bolivia–Tarija, Bolivia; Sociedad Geologica: Cochabamba, Boliviana, 1996; pp. 523–529. [Google Scholar]
- Edwards, D.; Banks, H.P.; Ciurca, S.J., Jr.; Laub, R.S. New Silurian cooksonias from dolostones of north-eastern North America. Bot. J. Linn. Soc. 2004, 146, 399–413. [Google Scholar] [CrossRef]
- Edwards, D.; Rogerson, E.C.W. New records of fertile Rhyniophytina from the late Silurian of Wales. Geol. Mag. 1979, 116, 93–98. [Google Scholar] [CrossRef]
- Douglas, J.; Holmes, F. The Baragwanathia story: An update. Foss. Collect. 2006, 77, 9–26. [Google Scholar]
- Edwards, D.; Morel, E.M.; Paredes, F.; Ganuza, D.G.; Zúniga, A. Plant assemblages from the Silurian of southern Bolivia and their palaeogeographic significance. Bot. J. Linn. Soc. 2001, 135, 229–250. [Google Scholar] [CrossRef]
- Hagström, J.; Mehlqvist, K. The dawn of terrestrial ecosystems on Baltica: First report on land plant remains and arthropod coprolites from the Upper Silurian of Gotland, Sweden. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 317–318, 162–170. [Google Scholar] [CrossRef]
- Rogerson, C.; Edwards, D.; Axe, L.; Davies, K.L. A new Embryophyte from the Upper Silurian of Shropshire, England. Spec. Pap. Palaeontol. 2002, 67, 233–249. [Google Scholar]
- Fanning, U.; Edwards, D.; Richardson, J.B. Further evidence for diversity in late Silurian land vegetation. J. Geol. Soc. Lond. 1990, 147, 725–728. [Google Scholar] [CrossRef]
- Fanning, U.; Edwards, D.; Richardson, J.B. A new rhyniophytoid from the late Silurian of the Welsh Borderland. Neues Jahrb. Geol. Paläontol. Abh. 1991, 183, 37–47. [Google Scholar]
- Fanning, U.; Richardson, J.B.; Edwards, D. A review of in situ spores in Silurian land plants. In Systematics Association, Pollen and Spores, Special Volume; Blackmore, S., Barnes, S.H., Eds.; Clarendon Press: Oxford, UK, 1991; Volume 44, pp. 25–47. [Google Scholar]
- Edwards, D. A late Silurian flora from the Lower Old Red Sandstone of South-West Dyfed. Palaeontology 1979, 22, 23–52. [Google Scholar]
- Bodzioch, A.; Kozlowski, W.; Poplawska, A. A Cooksonia−type flora from the Upper Silurian of the Holy Cross Mountains, Poland. Acta Palaeontol. Pol. 2003, 48, 653–656. [Google Scholar]
- Ishchenko, T.A. The Late Silurian Flora of Podolia; Institute of Geological Science, Academy of Science of the Ukrainian SSR: Kiev, Ukrainian, 1975; pp. 1–80. [Google Scholar]
- Senkevich, M.A. Fossil plants in the Tokrau horizon of the Upper Silurian. In The Tokrau Horizon of the Upper Silurian Series: Balkhash Segment; Nikitin, I.F., Bandaletoc, S.M., Eds.; Nauka, Alma-Ata: Almaty, Kazakhstan, 1986. (In Russian) [Google Scholar]
- Senkevich, M.A. New Devonian psilophytes from Kazakhstan. Esheg Vses Paleontol Obs. 1975, 21, 288–298. (In Russian) [Google Scholar]
- Cai, C.Y.; Dou, Y.W.; Edwards, D. New observations on a Přídolí plant assemblage from north Xinjing, northwest China, with comments on its evolutionary and palaeogeographical significance. Geol. Mag. 1993, 130, 155–170. [Google Scholar]
- Gonez, P.; Huu, H.N.; Hoa, P.T.; Clément, G.; Janvire, P. The oldest flora of the South China Block, and the stratigraphic bearings of the plant remains from the Ngoc Vung Series, northern Vietnam. J. Asian Earth Sci. 2012, 43, 51–63. [Google Scholar] [CrossRef]
- Edwards, D.; Feehan, J.; Smith, D.G. A late Wenlock flora from Co. Tipperary, Ireland. Bot. J. Linn. Soc. 1983, 86, 19–36. [Google Scholar] [CrossRef]
- Rogerson, E.C.W.; Edwards, D.; Davies, K.L.; Richardson, J.B. Identification of in situ spores in a Silurian Cooksonia from the Welsh Borderland. In Studies in Palaeobotany and Palynology in Honour of Professor W. G. Chaloner, F.R.S.; Collinson, M.E., Scott, A.C., Eds.; Special Papers in Palaeontology; Palaeontological Association: Durham, UK, 1993; Volume 49, pp. 17–30. [Google Scholar]
- Daber, R. Cooksonia—One of the most ancient psilophytes—Widely distributed, but rare. Botanique 1971, 2, 35–40. [Google Scholar]
- Morel, E.; Edwards, D.; Inigez Rodriguez, M. The first record of Cooksonia from South America in Silurian rocks of Bolivia. Geol. Mag. 1995, 132, 449–452. [Google Scholar] [CrossRef]
- Lang, W.H. On plant-remains from the Downtonian of England and Wales. Philos. Trans. R. Soc. Lond. 1937, B227, 245–291. [Google Scholar]
- Tims, J.D.J. The Early Land Flora of Victoria. Ph.D. Thesis, University of Melbourne, Melbourne, Australia, 1980. [Google Scholar]
- Dou, Y.W.; Sun, Z.H. Devonian Plants. In Palaeontological Atlas of Xinjiang, Vol II, Late Palaeozoic Section; Geological Publishing House: Beijing, China, 1983. (In Chinese) [Google Scholar]
- Steemans, P.; Petus, E.; Breuer, P.; Mauller-Mendlowicz, P.; Gerrienne, P. Palaeozoic innovations in the micro and megafossil plant record from the earliest plant spores to the earliest seeds. In Earth and life: Global Biodiversity, Extinction Intervals and Biogeographic Perturbations through Time; Talent, J.A., Ed.; International Year of Planet Eart; Springer Science: Dordrecht, The Netherlands, 2012; pp. 437–477. [Google Scholar]
- Mehlqvist, K.; Vajda, V.; Steemans, P. Early land plant spore assemblages from the Late Silurian of Skåne, Sweden. GFF 2012, 134, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Gray, J. The microfossil record of early land plants: Advances in understanding early terrestrialization, 1870–1984. Philos. Trans. Royal. Soc. Lond. Ser. B 1985, 309, 167–195. [Google Scholar]
- Morris, J.L.; Wright, V.P.; Edwards, D. Siluro-Devonian landscapes of southern Britain: The stability and nature of early vascular plant habitats. J. Geol. Soc. 2012, 169, 173–190. [Google Scholar] [CrossRef]
- Gray, J. Major Paleozoic land plant evolutionary bio-events. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1993, 104, 153–169. [Google Scholar] [CrossRef]
- Wellman, C.H. Origin, function and development of the spore wall in early land plants. In The Evolution of Plant Physiology; Hemsleyx, A.R., Poole, I., Eds.; Academic Press: Cambridge, MA, USA, 2004; pp. 43–63. [Google Scholar]
- Mogensen, G.S. The biological significance of morphological characters in bryophytes: The spore. Bryologist 1981, 84, 187–207. [Google Scholar] [CrossRef]
- Allen, K.C. A review of in situ late Silurian and Devonian spores. Rev. Palaeobot. Palynol. 1980, 29, 253–270. [Google Scholar] [CrossRef]
- Balme, B.A. Fossil in situ spores and pollen grains: An annotated catalogue. Rev. Palaeobot. Palynol. 1995, 87, 81–323. [Google Scholar] [CrossRef]
- Stemans, P. Miospore evolution from the Ordovician to the Silurian. Rev. Palaeobot. Palynol. 2000, 113, 189–196. [Google Scholar] [CrossRef]
- Wellman, C.H.; Steemans, P.; Vecoli, M. Palaeophytogeography of Ordovician-Silurian Land Plants. Geol. Soc. Lond. Mem. 2014, 38, 461–476. [Google Scholar] [CrossRef]
- Brown, R.C.; Lemmon, B.E.; Shimamura, M.; Villarreal, J.C.; Renzaglia, K.S. Spores of relictual bryophytes: Diverse adaptations to life on land. Rev. Palaeobot. Palynol. 2015, 216, 1–17. [Google Scholar] [CrossRef]
- Mehlqvist, K.; Steemans, P.; Vajda, V. First evidence of Devonian strata in Sweden—A palynological investigation of Övedskloster drillcores 1 and 2, Skåne, Sweden. Rev. Palaeobot. Palynol. 2015, 221, 144–159. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Xue, J. The early Devonian Posongchong flora of Yunnan: A Contribution to an Understanding of the Evolution and Early Diversification of Vascular Plants; Science Press: Beijing, China, 2013; pp. 1–366. [Google Scholar]
- Harris, B.J.; Harrison, C.J.; Hetherington, A.M.; Williams, T.A. Phylogenomic Evidence for the Monophyly of Bryophytes and the Reductive Evolution of Stomata. Curr. Biol. 2020, 30, P2001-20012E2. [Google Scholar] [CrossRef]
- Lehnert, O.; Frýda, J.; Joachimski, M.; Meinhold, G.; Čáp, P. A latest Silurian Supergreenhouse: The trigger for the Pridoli Transgrediens Extinction Event. In Proceedings of the 34th International Geological Congress, Brisbane, Australia, 5–10 August 2012; p. 3255. [Google Scholar]
- Qu, Q.M.; Zhu, M.; Zhao, W.J. Silurian atmospheric O2 changes and the early radiation of gnathostomes. Palaeoworld 2010, 19, 146–159. [Google Scholar] [CrossRef]
- Kerp, H.; Hass, H. De Onder-Devonische Rhynie Chert—Het oudste en meest compleet bewaard gebleven terrestriche. Grondboor Hamer 2004, 58, 33–50. [Google Scholar]
- Taylor, T.N.; Kerp, H.; Hass, H. Life history biology of early land plants: Deciphering the gametophyte phase. Proc. Natl. Acad. Sci. USA 2005, 102, 5892–5897. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Pri-Tal, O.; Michaeli, D.; Mosquna, A. Evolution of Abscisic Acid Signaling Module and Its Perception. Front. Plant Sci. 2020, 11, 934. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Kubo, M.; Akiyoshi, N.; Sano, R.; Nomura, T.; Sawa, S.; Ohtani, M.; Demura, T. Expression of peat moss VASCULAR RELATED NAC-DOMAIN homologs in Nicotiana benthamiana leaf cells induces ectopic secondary wall formation. Plant Mol. Biol. 2021, 106, 309–317. [Google Scholar] [CrossRef]
Figure 1.
Silurian time scale showing conodont and graptolite biozones, stage slices (s.s.), and generalized δ13Ccarb curve, modified from [20,21,22], combined with isotopic temperature curves (blue based on brachiopod, and red on conodont δ18O data) from [23]. The conodont isotopic temperature curves for Ludfordian were modified based on [24]. The isotopic temperatures assume “ice-free” conditions (δ18Oseawater = −1.1‰ vs. VSMOW). Abreviations: Shein.—Sheinwoodian; Hom.—Homerian; Gor.—Gorstian; Lud.—Ludfordian; Loch.—Lochkovian. Abreviations used in graptolite column, bottom to top: Cy.—Cyrtograptus; M. riccarton.—Monograptus riccartonensis; Pr. par./G. nas.—Pristiograptus parvus/Gothograptus nassa; Col.—Colonograptus; Col. prae./deub.—Col. praedeubeli/Col. deubeli; N./ Lo.—Neodiversograptus/ Lobograptus; Lo.—Lobograptus; Sa. leintward./Sa. Linearis—Saetograptus linearis; Ne./Po.—Neocucullograptus/Polonograptus; Neo.—Skalograptus; Neo. parult.—Neocolonograptus parultimus; M. loc./M. bra.—Skalograptus lochkovensis/M. lochkovensis branikensis; M.—Monograptus. Conodont column, bottom to top: procerus—Pterospathodus pennatus procerus; K.—Kockelella; K. ranuli.—K. ranuliformis; O. s. rhenana—Ozarkodina sagitta rhenana; O. s. sagitta—Ozarkodina sagitta sagitta; O. boh. Longa—Ozarkodina bohemica longa; K. o. absid.—Kockelella ortus absidata; A.—Ancoradella; Po.—Polygnathoides; Ped.—Pedavis; O.—Ozarkodina; ‘O.’ eostein.—“Ozarkodina” eosteinhornensis sensu lato; Ou.—Oulodus. S.Z.—Superbiozone; Z.G.—Zonal Group; I.Z.—Interval Biozone.
Figure 1.
Silurian time scale showing conodont and graptolite biozones, stage slices (s.s.), and generalized δ13Ccarb curve, modified from [20,21,22], combined with isotopic temperature curves (blue based on brachiopod, and red on conodont δ18O data) from [23]. The conodont isotopic temperature curves for Ludfordian were modified based on [24]. The isotopic temperatures assume “ice-free” conditions (δ18Oseawater = −1.1‰ vs. VSMOW). Abreviations: Shein.—Sheinwoodian; Hom.—Homerian; Gor.—Gorstian; Lud.—Ludfordian; Loch.—Lochkovian. Abreviations used in graptolite column, bottom to top: Cy.—Cyrtograptus; M. riccarton.—Monograptus riccartonensis; Pr. par./G. nas.—Pristiograptus parvus/Gothograptus nassa; Col.—Colonograptus; Col. prae./deub.—Col. praedeubeli/Col. deubeli; N./ Lo.—Neodiversograptus/ Lobograptus; Lo.—Lobograptus; Sa. leintward./Sa. Linearis—Saetograptus linearis; Ne./Po.—Neocucullograptus/Polonograptus; Neo.—Skalograptus; Neo. parult.—Neocolonograptus parultimus; M. loc./M. bra.—Skalograptus lochkovensis/M. lochkovensis branikensis; M.—Monograptus. Conodont column, bottom to top: procerus—Pterospathodus pennatus procerus; K.—Kockelella; K. ranuli.—K. ranuliformis; O. s. rhenana—Ozarkodina sagitta rhenana; O. s. sagitta—Ozarkodina sagitta sagitta; O. boh. Longa—Ozarkodina bohemica longa; K. o. absid.—Kockelella ortus absidata; A.—Ancoradella; Po.—Polygnathoides; Ped.—Pedavis; O.—Ozarkodina; ‘O.’ eostein.—“Ozarkodina” eosteinhornensis sensu lato; Ou.—Oulodus. S.Z.—Superbiozone; Z.G.—Zonal Group; I.Z.—Interval Biozone.

Figure 2.
Silurian palaeocontinental reconstructions according to Scotese [76] plotting palaeogeographical distribution of plants (rectangle) or plant assemblages (circle). Colored ellipses represent individual phytogeographic units according to Raymond et al. [48]; W = Wenlock, L = Ludlow, P = Přídolí.
Figure 2.
Silurian palaeocontinental reconstructions according to Scotese [76] plotting palaeogeographical distribution of plants (rectangle) or plant assemblages (circle). Colored ellipses represent individual phytogeographic units according to Raymond et al. [48]; W = Wenlock, L = Ludlow, P = Přídolí.


{kind=link}
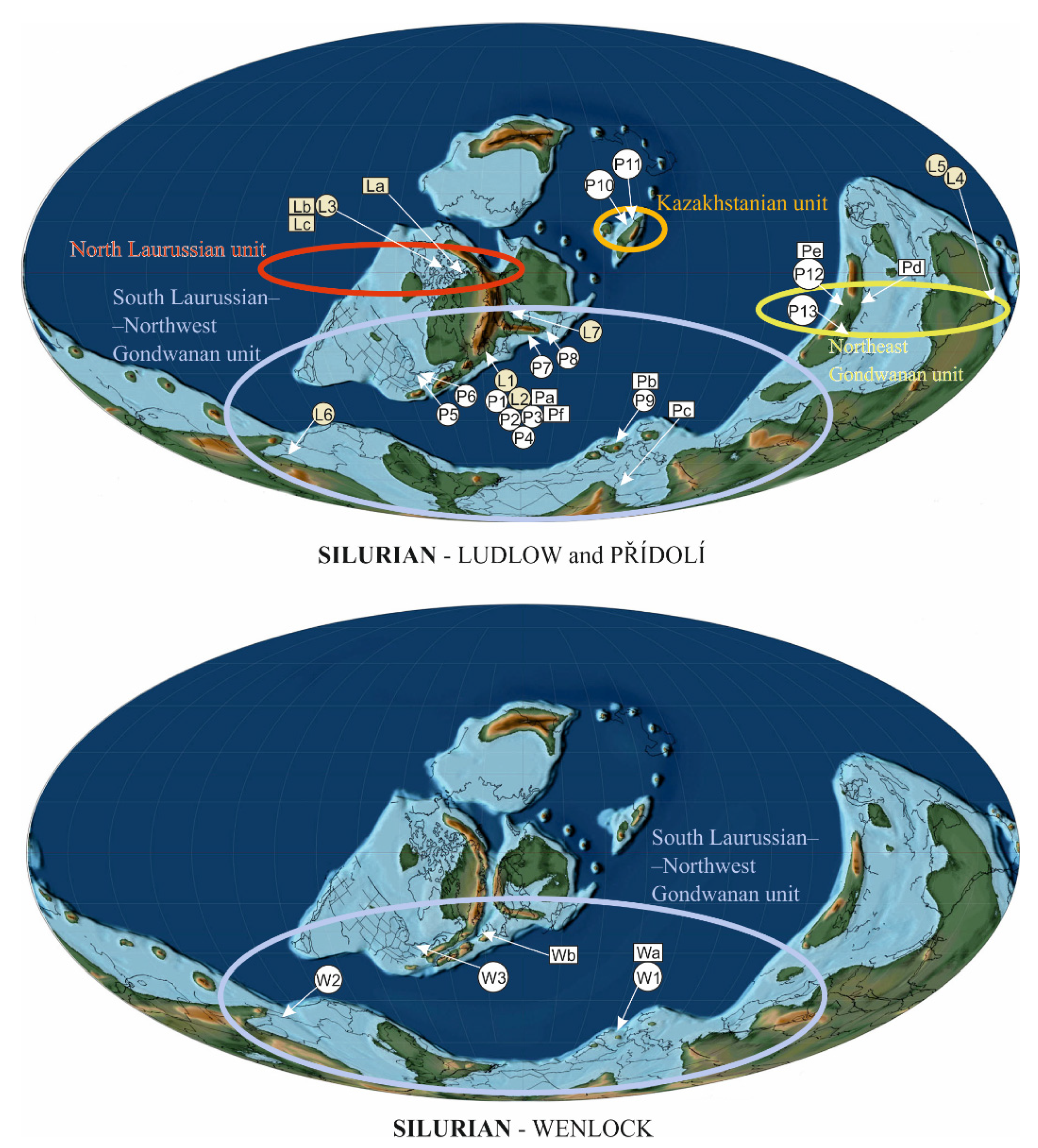{kind=link}
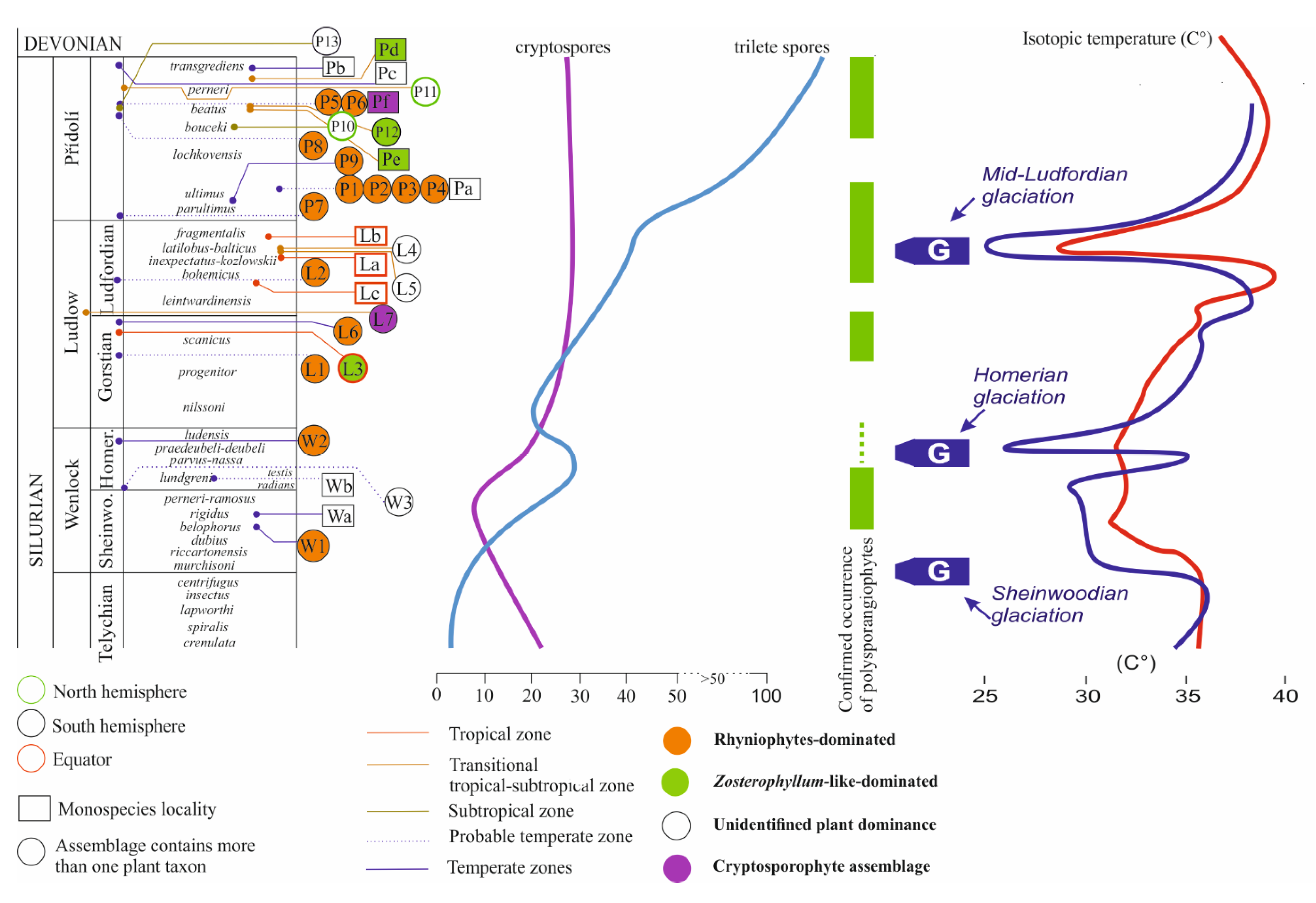{kind=link}
{kind=link}
{kind=link}
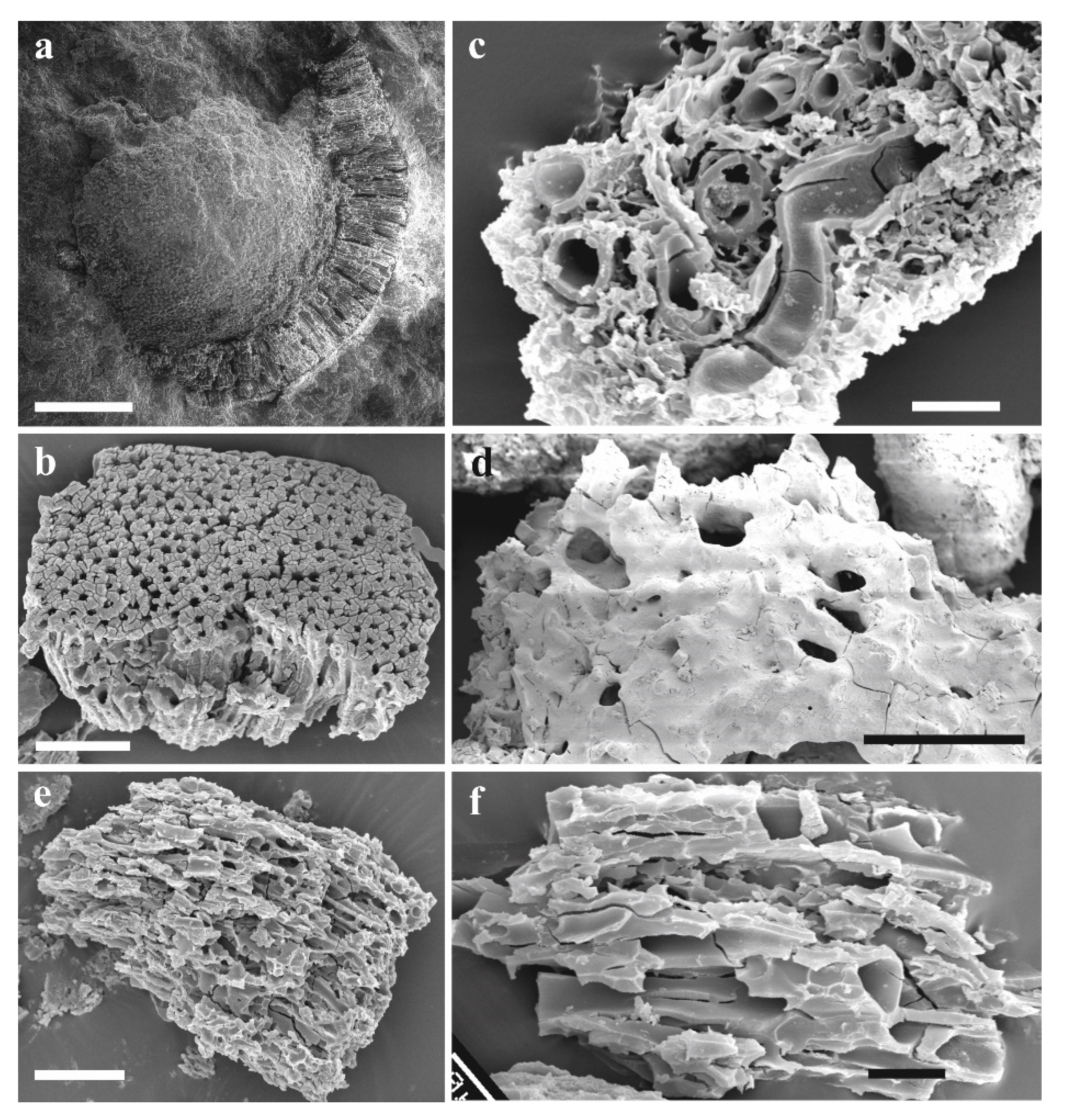{kind=link}
{kind=link}
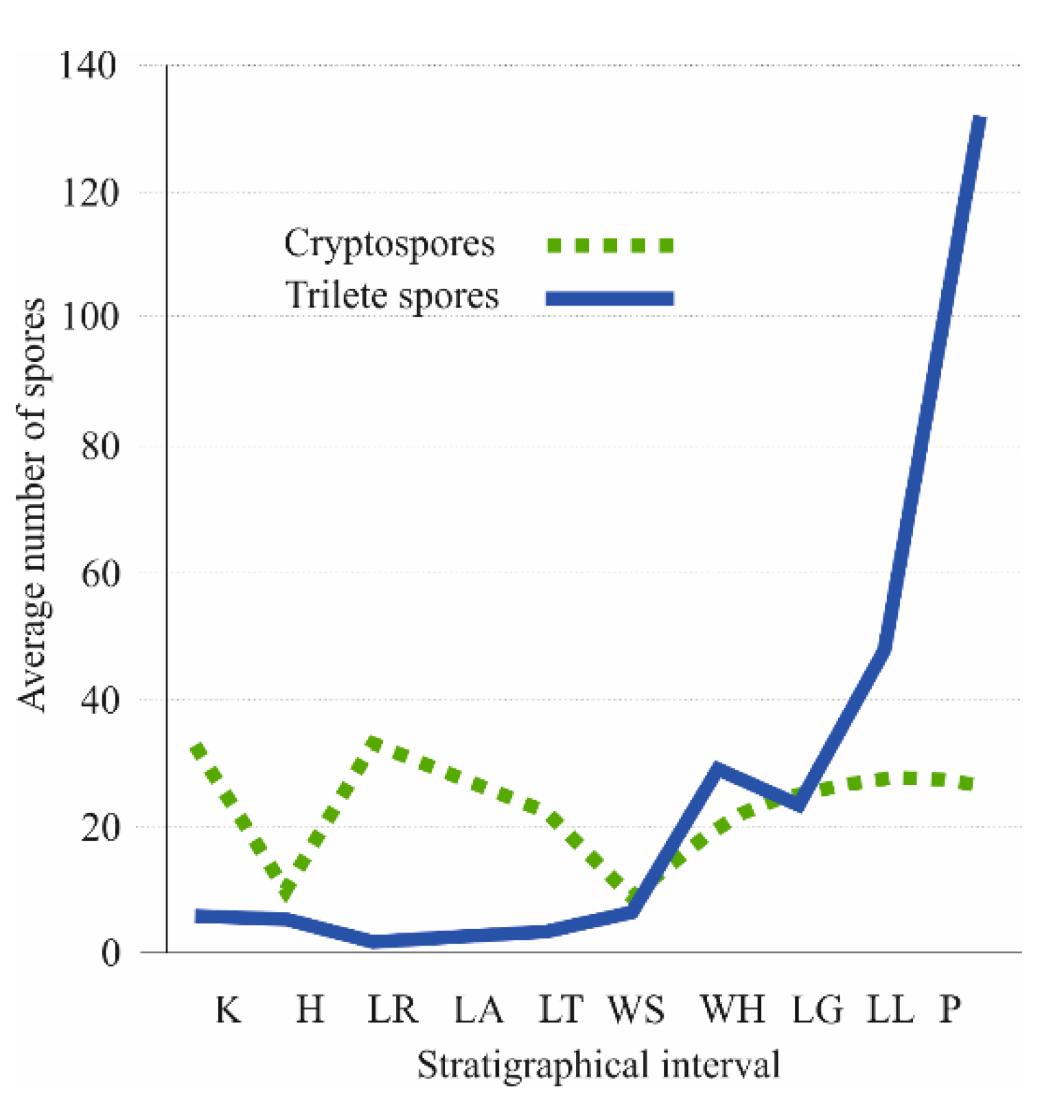{kind=link}
Table 1.
Quantitative occurrence of main types of cryptospores, hilate spores and true trilete spores from Llandovery to Přídolí.
Table 1.
Quantitative occurrence of main types of cryptospores, hilate spores and true trilete spores from Llandovery to Přídolí.
| Age | Envelope Enclosed Monads | Envelope Enclosed Dyads | Envelope Enclosed Tetrad | Naked Monads | Naked Dyads | Naked Tetrads | Hilate Spores | Trilete Spores |
|---|---|---|---|---|---|---|---|---|
| Přídolí | █ █ | █ █ | █ █ | ██ ██ | █ █ | █ █ | ███ ███ | ███ ███ |
| Ludlow | █ █ | █ █ | █ █ | ██ ██ | █ █ | █ █ | ██ ██ | ██ ██ |
| Wenlock | █ █ | █ █ | █ █ | █ █ | ██ ██ | ██ ██ | ██ ██ | ██ ██ |
| Llandovery | █ █ | █ █ | █ █ | █ █ | ██ ██ | ██ ██ | █ █ | █ █ |
| ██ ██ | ███ ███ | ███ ███ | █ █ | ███ ███ | ███ ███ | █ █ | █ █ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pšenička, J.; Bek, J.; Frýda, J.; Žárský, V.; Uhlířová, M.; Štorch, P. Dynamics of Silurian Plants as Response to Climate Changes. Life 2021, 11, 906. https://doi.org/10.3390/life11090906
AMA Style
Pšenička J, Bek J, Frýda J, Žárský V, Uhlířová M, Štorch P. Dynamics of Silurian Plants as Response to Climate Changes. Life. 2021; 11(9):906. https://doi.org/10.3390/life11090906
Chicago/Turabian StylePšenička, Josef, Jiří Bek, Jiří Frýda, Viktor Žárský, Monika Uhlířová, and Petr Štorch. 2021. "Dynamics of Silurian Plants as Response to Climate Changes" Life 11, no. 9: 906. https://doi.org/10.3390/life11090906
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.