
Changing Temperature Conditions during Somatic Embryo Maturation Result in Pinus pinaster Plants with Altered Response to Heat Stress

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Results
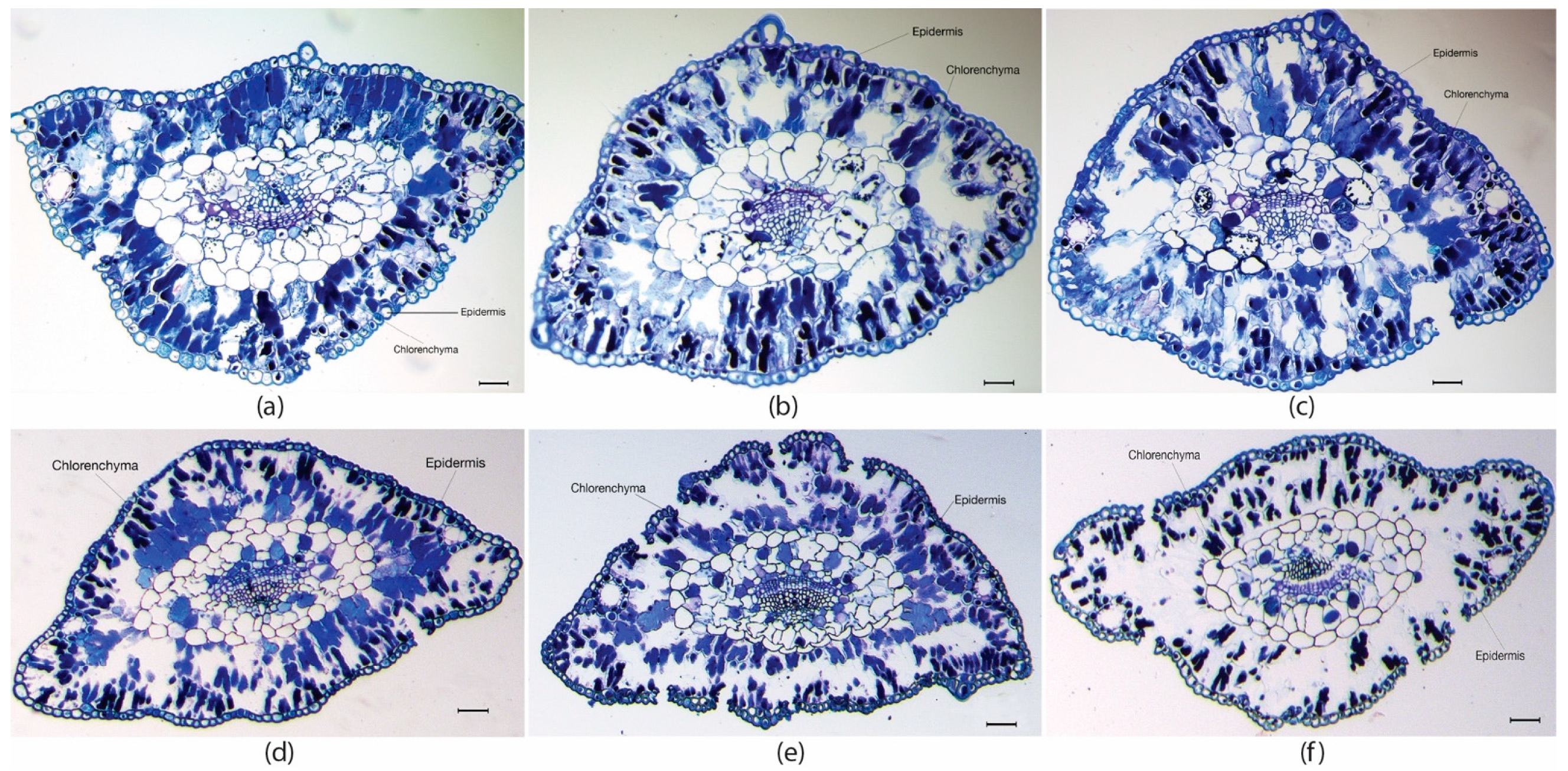
2.1. Plant Height and Histological Determinations in Maritime Pine Needles
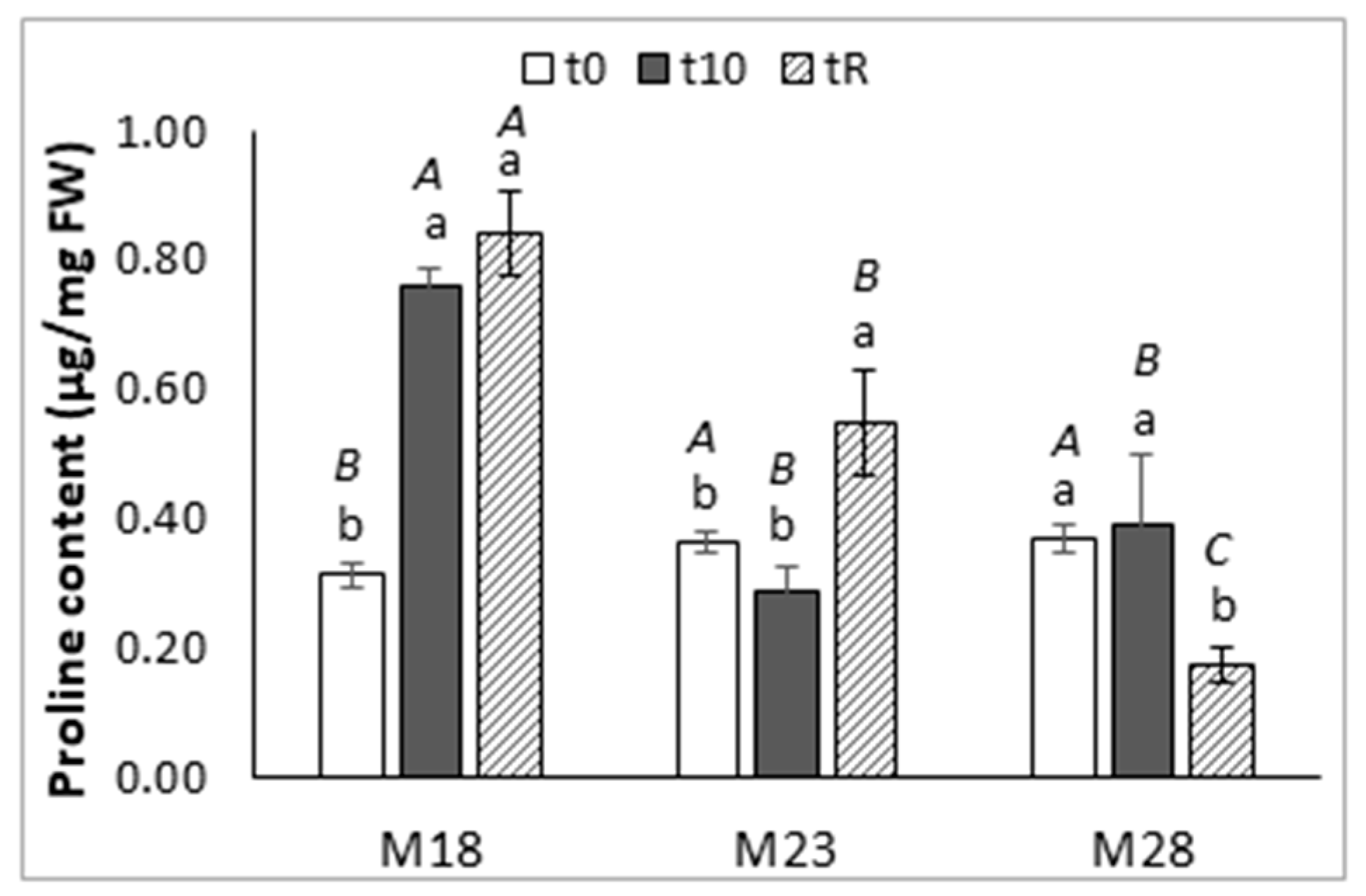
2.2. Water Relations and Osmotic Adjustment
2.3. Photosynthesiselated Parameters
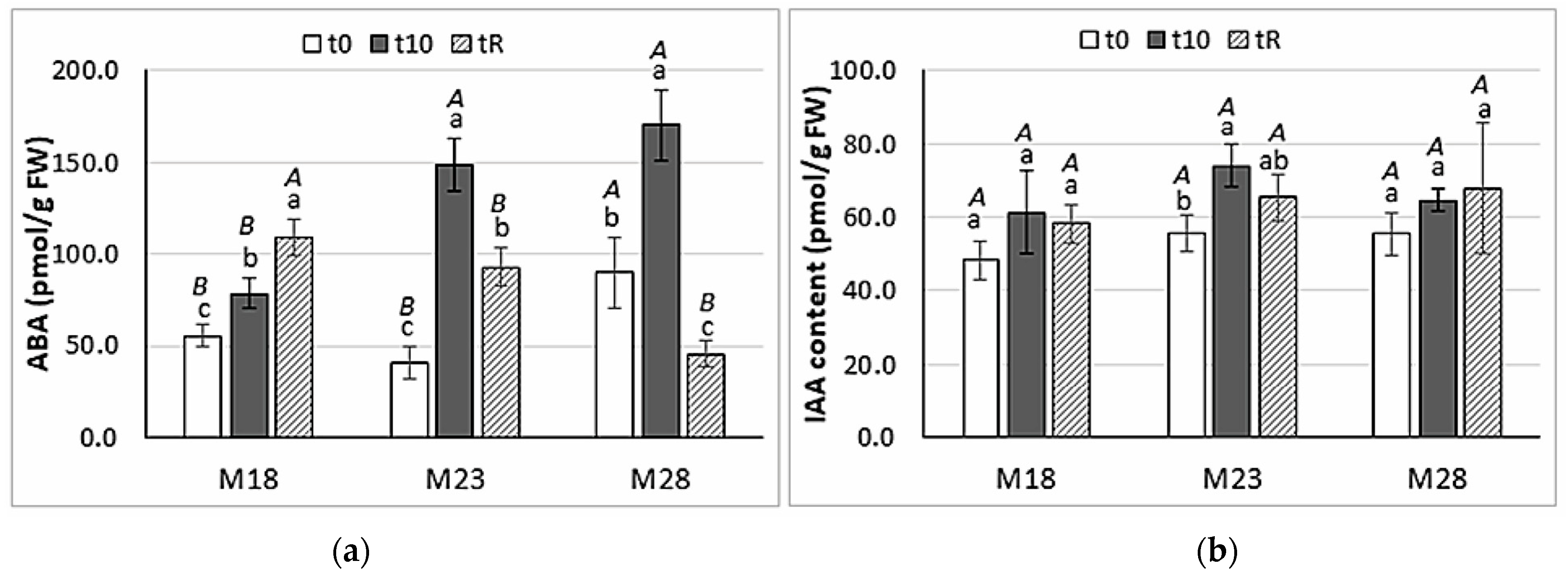
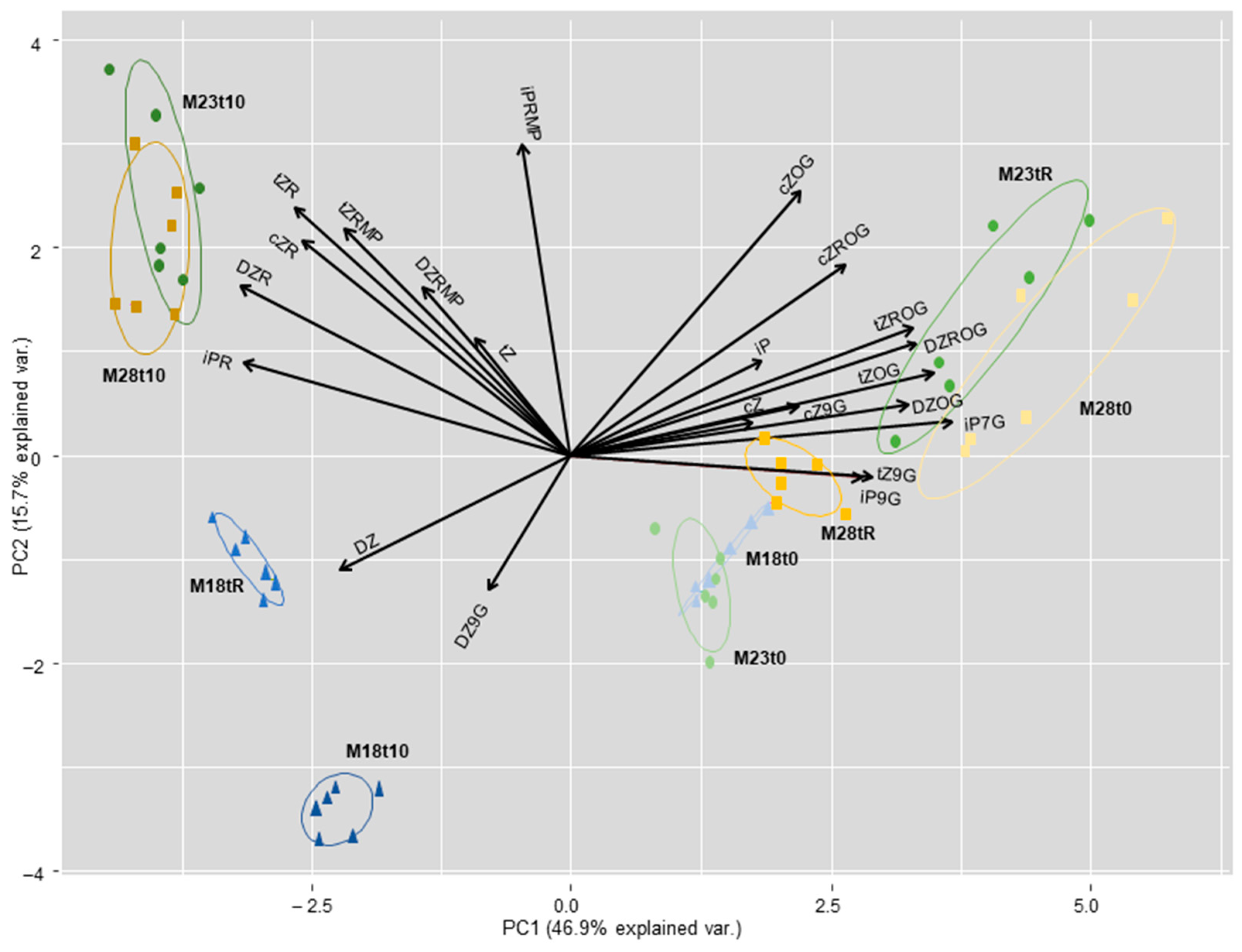
2.4. Hormone Content
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Osmotic Adjustment Determination
4.3. Photosynthesis-related Parameters
4.4. Hormone Content Determinations
4.5. Histological Analyses
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zas, R.; Sampedro, L.; Solla, A.; Vivas, M.; Lombardero, M.J.; Alía, R.; Rozas, V. Dendroecology in common gardens: Population differentiation and plasticity in resistance, recovery and resilience to extreme drought events in Pinus pinaster. Agric. For. Meteorol. 2020, 291, 108060. [Google Scholar] [CrossRef]
- Alía, R.; Martín, S. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Maritime Pine (Pinus pinaster); International Plant Genetic Resources Institute; Bioversity International: Rome, Italy, 2003; p. 6. [Google Scholar]
- Nguyen-Queyrens, A.; Bouchet-Lannat, F. Osmotic adjustment in three-year-old seedlings of five provenances of maritime pine (Pinus pinaster) in response to drought. Tree Physiol. 2003, 23, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menéndez-Gutiérrez, M.; Alonso, M.; Toval, G.; Díaz, R. Variation in pinewood nematode susceptibility among Pinus pinaster Ait. provenances from the Iberian Peninsula and France. Ann. For. Sci. 2017, 74, 76. [Google Scholar] [CrossRef] [Green Version]
- Arrillaga, I.; Guevara, M.A.; Muñoz-Bertomeu, J.; Lázaro-Gimeno, D.; Sáez-Laguna, E.; Díaz, L.M.; Torralba, L.; Mendoza-Poudereux, I.; Segura, J.; Cervera, M.T. Selection of haploid cell lines from megagametophyte cultures of maritime pine as a DNA source for massive sequencing of the species. Plant Cell Tiss. Org. Cult. 2014, 118, 147–155. [Google Scholar] [CrossRef]
- Arrillaga, I.; Morcillo, M.; Zanón, I.; Lario, F.; Segura, J.; Sales, E. New approaches to optimize somatic embryogenesis in maritime pine. Front. Plant Sci. 2019, 10, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andivia, E.; Ruiz-Benito, P.; Díaz-Martínez, P.; Carro-Martínez, N.; Zavala, M.A.; Madrigal-González, J. Inter-specific tolerance to recurrent droughts of pine species revealed in saplings rather than adult trees. For. Ecol. Manag. 2020, 459, 117858. [Google Scholar] [CrossRef]
- Neumann, M.; Mues, V.; Moreno, A.; Hasenauer, H.; Seidl, R. Climate variability drives recent tree mortality in Europe. Glob. Chang. Biol. 2017, 23, 4788–4797. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Lebedeva, T.N.; Chernodubov, A.I.; Shestibratov, K.A. Genomic selection for forest tree improvement: Methods, achievements and perspectives. Forests 2020, 11, 1190. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Klimaszewska, K.; Miguel, C.; Aronen, T.; Hargreaves, C.; Teyssier, C.; Trontin, J.F. Somatic embryogenesis for more effective breeding and deployment of improved varieties in Pinus spp.: Bottlenecks and recent advances. In Somatic Embryogenesis: Fundamental Aspects and Applications; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 319–365. [Google Scholar]
- Lelu-Walter, M.A.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic embryogenesis in forestry with a focus on Europe: State-of-the-art, benefits, challenges and future direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Turgut-Kara, N.; Arikan, B.; Celik, H. Epigenetic memory and priming in plants. Genetica 2020, 148, 47–54. [Google Scholar] [CrossRef]
- Galviz, Y.C.F.; Ribeiro, R.V.; Souza, G.M. Yes, plants do have memory. Theor. Exp. Plant Physiol. 2020, 32, 195–202. [Google Scholar] [CrossRef]
- Kvaalen, H.; Johnsen, Ø. Timing of bud set in Picea abies is regulated by a memory of temperature during zygotic and somatic embryogenesis. New Phytol. 2008, 177, 49–59. [Google Scholar] [CrossRef]
- Egertsdotter, U. Plant physiological and genetical aspects of the somatic embryogenesis process in conifers. Scand. J. For. Res. 2019, 34, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Cano, M.; Morcillo, A.; Humánez, A.; Mendoza-Poudereux, I.; Alborch, A.; Segura, J.; Arrillaga, I. Maritime pine (Pinus pinaster Aiton). In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants, 2nd ed.; Jain, S.M., Gupta, P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1, pp. 167–179. [Google Scholar]
- Humánez, A.; Blasco, M.; Brisa, C.; Segura, J.; Arrillaga, I. Somatic embryogenesis from different tissues of Spanish populations of maritime pine. Plant Cell Tissue Organ Cult. 2012, 111, 373–383. [Google Scholar] [CrossRef]
- Pérez-Oliver, M.A.; Haro, J.G.; Pavlović, I.; Novák, O.; Segura, J.; Sales, E.; Arrillaga, I. Priming maritime pine megagametophytes during somatic embryogenesis improved plant adaptation to heat stress. Plants 2021, 10, 446. [Google Scholar] [CrossRef]
- García-Mendiguren, O.; Montalbán, I.A.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Environmental conditions at the initial stages of Pinus radiata somatic embryogenesis affect the production of somatic embryos. Trees 2016, 30, 949–958. [Google Scholar] [CrossRef]
- Marques do Nascimento, A.M.; Alves-Barroso, P.; Ferreira do Nascimento, N.F.; Goicoa, T.; Ugarte, M.D.; Montalbán, I.A.; Moncaleán, P. Pinus spp. somatic embryo conversion under high temperature: Effect on the morphological and physiological characteristics of plantlets. Forests 2020, 11, 1181. [Google Scholar] [CrossRef]
- Chen, W.L.; Yang, W.J.; Lo, H.F.; Yeh, D.M. Physiology, anatomy, and cell membrane thermostability selection ofleafy radish (Raphanus sativus var. Oleiformis Pers.) with different tolerance under heat stress. Sci. Hortic. 2014, 179, 367–375. [Google Scholar] [CrossRef]
- Estravis-Barcala, M.; Mattera, M.G.; Soliani, C.; Bellora, N.; Opgenoorth, L.; Heer, K.; Arana, M.V. Molecular bases of responses to abiotic stress in trees. J. Exp. Bot. 2020, 71, 3765–3779. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a multifaceted signalling molecule in plant responses to abiotic stress: Understanding the physiological mechanisms. Plant Biol. 2021. [Google Scholar] [CrossRef]
- Yu, D.; Wildhagen, H.; Tylewicz, S.; Miskolczi, P.C.; Bhalerao, R.P.; Polle, A. Abscisic acid signalling mediates biomass trade-off and allocation in poplar. New Phytol. 2019, 223, 1192–1203. [Google Scholar] [CrossRef]
- Shen, H.F.; Zhao, B.; Xu, J.J.; Liang, W.; Huang, W.M.; Li, H.H. Effects of heat stress on changes in physiology and anatomy in two cultivars of Rhododendron. S. Afr. J. Bot. 2017, 112, 338–345. [Google Scholar] [CrossRef]
- Li, M.; Jannasch, A.H.; Jiang, Y. Growth and Hormone Alterations in Response to Heat Stress in Perennial Ryegrass Accessions Differing in Heat Tolerance. J. Plant Growth Regul. 2020, 39, 1022–1102. [Google Scholar] [CrossRef]
- Pereira, C.; Castander-Olarieta, A.; Sales, E.; Montalbán, I.A.; Canhoto, J.; Moncaleán, P. Heat stress in Pinus halepensis somatic embryogenesis induction: Effect in DNA methylation and differential expression of stress-related genes. Plants 2021, 10, 2333. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Pereira, C.; Sales, E.; Meijón, M.; Arrillaga, I.; Cañal, M.J.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P.; Montalbán, I.A. Induction of radiata pine somatic embryogenesis at high temperatures provokes a long-term decrease in DNA methylation/hydroxymethylation and differential expression of stress-related genes. Plants 2020, 9, 1762. [Google Scholar] [CrossRef]
- Pereira, C.; Castander-Olarieta, A.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlovic, I.; De Medeiros Oliveira, E.; Fraga, H.; Guerra, M.P.; Novak, O.; et al. Embryonal masses induced at high temperatures in Aleppo pine: Cytokinin profile and cytological characterization. Forests 2020, 11, 807. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Pereira, C.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlović, I.; Novák, O.; Strnad, M.; Moncaleán, P. Quantification of endogenous aromatic cytokinins in Pinus radiata embryonal masses after application of heat stress during initiation of somatic embryogenesis. Trees 2020, 35, 1075–1080. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Moncaleán, P.; Pereira, C.; Pěnčík, A.; Petřík, I.; Pavlovic, I.; Novák, O.; Strnad, M.; Goicoa, T.; Ugarte, M.D.; et al. Cytokinins are involved in drought tolerance of Pinus radiata plants originating from embryonal masses induced at high temperatures. Tree Physiol. 2021, 41, 912–926. [Google Scholar] [CrossRef]
- Moncaleán, P.; García-Mendiguren, O.; Novak, O.; Strnad, M.; Goicoa, T.; Ugarte, M.D.; Montalbán, I.A. Temperature and water availability during maturation affect the cytokinins and auxins profile of radiata pine somatic embryos. Front. Plant Sci. 2018, 9, 1898. [Google Scholar] [CrossRef] [Green Version]
- Escandón, M.; Cañal, M.; Pascual, J.; Pinto, G.; Correia, B.; Amaral, J.; Meijón, M. Integrated physiological and hormonal profile of heat-induced thermotolerance in Pinus radiata. Tree Physiol. 2016, 36, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Escandón, M.; Valledor, L.; Pascual, J.; Pinto, G.; Cañal, M.J.; Meijón, M. System-wide analysis of short-term response to high temperature in Pinus radiata. J. Exp. Bot. 2017, 68, 3629–3641. [Google Scholar] [CrossRef] [PubMed]
- Kolb, P.F.; Robberecht, R. High temperature and drought stress effects on survival of Pinus ponderosa seedlings. Tree Physiol. 1996, 16, 665–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turunen, M.; Latola, K. UV-B radiation and acclimation in timberline plants. Environ. Pollut. 2005, 137, 390–403. [Google Scholar] [CrossRef]
- Li, W.D.; Biswas, D.K.; Xu, H.; Xu, C.Q.; Wang, X.Z.; Liu, J.K.; Jiang, G.M. Photosynthetic responses to chromosome doubling in relation to leaf anatomy in Lonicera japonica subjected to water stress. Funct. Plant. Biol. 2009, 36, 783–792. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.; Steppe, K. Review: Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Yuan, L.; Tang, L.; Zhu, S.; Hou, J.; Chen, G.; Liu, F.; Liu, S.; Wang, C. Influence of heat stress on leaf morphology and nitrogen–carbohydrate metabolisms in two wucai (Brassica campestris L.) genotypes. Acta Soc. Bot. Pol. 2017, 86, 3554. [Google Scholar] [CrossRef]
- Natarajan, S.; Kuehny, J.S. Morphological, Physiological, and Anatomical Characteristics Associated with Heat Preconditioning and Heat Tolerance in Salvia splendens. J. Amer. Soc. Hort. Sci. 2008, 133, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, I.; Shalmani, A.; Ali, M.; Yang, Q.H.; Ahmad, H.; Li, F.B. Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Front. Plant Sci. 2021, 11, 2310. [Google Scholar] [CrossRef]
- Konôpková, A.; Kurjak, D.; Kmeť, J.; Klumpp, R.; Longauer, R.; Ditmarová, Ľ.; Gömöry, D. Differences in photochemistry and response to heat stress between silver fir (Abies alba Mill.) provenances. Trees 2018, 32, 73–86. [Google Scholar] [CrossRef]
- Kunert, N.; Hajek, P.; Hietz, P.; Morris, H.; Rosner, S.; Tholen, D. Summer temperatures reach the thermal tolerance threshold of photosynthetic decline in temperate conifers. Plant Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Devireddy, A.R.; Tschaplinski, T.J.; Tuskan, G.A.; Muchero, W.; Chen, J.G. Role of reactive oxygen species and hormones in plant responses to temperature changes. Int. J. Mol. Sci. 2021, 22, 8843. [Google Scholar] [CrossRef] [PubMed]
- Birami, B.; Gattmann, M.; Heyer, A.G.; Grote, R.; Arneth, A.; Ruehr, N.K. Heat waves alter carbon allocation and increase mortality of Aleppo pine under dry conditions. Front. For. Glob. Chang. 2018, 1, 8. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Per, T.S.; Khan, N.A. Salicylic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Signal. Behav. 2013, 8, e26374. [Google Scholar] [CrossRef] [Green Version]
- Rajametov, S.N.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Jeong, H.B.; Chae, W.B. Heat-tolerant hot pepper exhibits constant photosynthesis via increased transpiration rate, high proline content and fast recovery in heat stress condition. Sci. Rep. 2021, 11, 14328. [Google Scholar] [CrossRef]
- Correia, B.; Rodriguez, J.L.; Valledor, L.; Almeida, T.; Santos, C.; Cañal, M.J.; Pinto, G. Analysis of the expression of putative heat-stress related genes in relation to thermotolerance of cork oak. J. Plant Physiol. 2014, 171, 399–406. [Google Scholar] [CrossRef]
- Dias, M.C.; Santos, C.; Silva, S.; Pinto, D.C.G.A.; Silva, A.M.S. Physiological and Metabolite Reconfiguration of Olea europaea to cope and recover from a heat or high UV-B shock. J. Agric. Food Chem. 2020, 68, 11339–11349. [Google Scholar] [CrossRef]
- De Diego, N.; Saiz-Fernández, I.; Rodríguez, J.L.; Pérez-Alfocea, P.; Sampedro, M.C.; Barrio, R.J.; Lacuesta, M.; Moncaleán, P. Metabolites and hormones are involved in the intraspecific variability of drought hardening in radiata pine. J. Plant Physiol. 2015, 188, 64–71. [Google Scholar] [CrossRef]
- Hossain, M.A.; Li, Z.; Hoque, T.S.; Burritt, D.J.; Fujita, M.; Munné-Bosch, S. Heat or cold priming-induced cross-tolerance to abiotic stresses in plants: Key regulators and posible mechanisms. Protoplasma 2018, 255, 399–412. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Zhuang, L.; Gao, Y.; Huang, B. Abscisic acid mediation of drought priming-enhanced heat tolerance in tall fescue (Festuca arundinacea) and Arabidopsis. Physiol. Plant. 2019, 167, 488–501. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Kramna, B.; Gaudinova, A.; Knirsch, V.; Spichal, L.; Zatloukal, M.; Vankova, R. Heat Acclimation and Inhibition of Cytokinin Degradation Positively Affect Heat Stress Tolerance of Arabidopsis. Front. Plant Sci. 2020, 11, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ábrahám, E.; Hourton-cabassa, C.; Erdei, L.; Szabados, L. Methods for determination of proline in plants. Methods Mol. Biol. 2010, 639, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Svačinová, J.; Novák, O.; Plačková, L.; Lenobel, R.; Holík, J.; Strnad, M.; Doležal, K. A new approach for cytokinin isolation from Arabidopsis tissues using miniaturized purification: Pipette tip solid-phase extraction. Plant Methods 2012, 8, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.; Strnad, M.; Ljung, K.; Novák, O. Plant hormonomics: Multiple phyto-hormone profiling by targeted metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Needles Thickness x (µm) | Epidermis Thickness y (µm) | Chlorenchyma Thickness x (µm) | |||
|---|---|---|---|---|---|---|
| t0 | t10 | t0 | t10 | t0 | t10 | |
| M18 | 553 ± 30 b | 651 ± 32 a | 20.3 ± 0.9 a | 18.6 ± 2.3 ab | 143.0 ± 11.8 c | 184.8 ± 13.6 a |
| M23 | 542 ± 42 b | 657 ± 57 a | 18.8 ± 1.4 a | 18.9 ± 1.5 a | 139.8 ± 19.0 c | 159.0 ± 18.3 b |
| M28 | 635 ± 24 a | 557 ± 31 b | 19.1 ± 1.0 a | 16.6 ± 0.8 b | 166.8 ± 9.8 b | 136.4 ± 8.1 c |
| Parameter | Plants | t0 | t10 | tR |
|---|---|---|---|---|
| AN x | M18 | 7.4 ± 2.4 b | 7.0 ± 4.5 (−5.2%) a | 9.0 ± 2.7 (22.4%) a |
| M23 | 10.9 ± 3.8 a | 8.7 ± 2.9 (−20.5%) a | 10.6 ± 2.0 (−3.2%) a | |
| M28 | 9.9 ± 2.0 a | 9.1 ± 0.7 (−8.4%) a | 9.5 ± 1.0 (−4.0%) a | |
| gs y | M18 | 0.24 ± 0.06 a | 0.17 ± 0.06 (−29.2%) a | 0.17 ± 0.12 (−29.2%) a |
| M23 | 0.24 ± 0.11 a | 0.16 ± 0.07 (−33.3%) a | 0.17 ± 0.09 (−29.2%) a | |
| M28 | 0.25 ± 0.12 a | 0.19 ± 0.09 (−24.0%) ab | 0.13 ± 0.01 (−48.0%) b | |
| M18 | 313 ± 31 a | 294 ± 54 (−6.1%) ab | 264 ± 39 (−15.4%) b | |
| (Ci) z | M23 | 282 ± 12 a | 270 ± 36 (−4.3%) a | 255 ± 35 (−9.6%) a |
| M28 | 292 ± 21 a | 271 ± 52 (−7.2%) ab | 235 ± 39 (−19.4%) b |
| Plants | Time | Total iP-Types | Total tZ-Types | Total DZ-Types | Total cZ-Types |
|---|---|---|---|---|---|
| M18 | t0 | 0.34 ± 0.04 a | 1.45 ± 0.08 a | 2.30 ± 0.07 a | 2.65 ± 0.16 a |
| t10 | 0.22 ± 0.03 b | 0.34 ± 0.05 b | 0.86 ± 0.12 ab | 0.87 ± 0.08 b | |
| tR | 0.35 ± 0.04 a | 0.47 ± 0.05 ab | 0.62 ± 0.03 b | 1.87 ± 0.04 a | |
| M23 | t0 | 0.26 ± 0.04 b | 1.38 ± 0.05 ab | 1.56 ± 0.09 ab | 2.26 ± 0.06 b |
| t10 | 0.45 ± 0.06 a | 0.95 ± 0.11 b | 0.87 ± 0.15 b | 2.67 ± 0.22 ab | |
| tR | 0.45 ± 0.08 a | 2.72 ± 0.52 a | 4.12 ± 0.50 a | 4.22 ± 0.50 a | |
| M28 | t0 | 0.28 ± 0.06 b | 3.73 ± 0.62 a | 2.44 ± 0.34 a | 6.81 ± 1.14 a |
| t10 | 0.50 ± 0.12 a | 0.82 ± 0.05 b | 0.64 ± 0.09 b | 2.90 ± 0.28 b | |
| tR | 0.28 ± 0.03 b | 2.56 ± 0.14 a | 2.19 ± 0.16 a | 4.60 ± 0.32 ab |
| Plants | Time | Active Forms | Transport Forms | Precursors | Reversible Metabolites | Irreversible Metabolites | Total CK y |
|---|---|---|---|---|---|---|---|
| CKs Bases x | CK Ribosides y | CK Nucleotides y | CK O-glucosides y | CK N-glucosides y | |||
| M18 | t0 | 0.16 ± 0.03 (2.4%) a | 0.06 ± 0.01 (0.9%) b | 0.20 ± 0.02 (3.0%) a | 6.28 ± 0.17 (93.3%) a | 0.03 ± 0.00 (0.4%) a | 6.73 ± 0.18 a |
| t10 | 0.16 ± 0.03 (7.1%) a | 0.13 ± 0.02 (5.9%) ab | 0.09 ± 0.01 (3.0%) b | 1.89 ± 0.21 (82.6) b | 0.01 ± 0.00 (0.5%) ab | 2.28 ± 0.26 b | |
| tR | 0.18 ± 0.03 (5.3%) a | 0.24 ± 0.03 (7.1%) a | 0.16 ± 0.01 (4.8%) ab | 2.74 ± 0.06 (82.6%) ab | 0.01 ± 0.00 (0.2%) b | 3.31 ± 0.11 ab | |
| M23 | t0 | 0.12 ± 0.04 (2.2%) b | 0.05 ± 0.01 (0.9%) b | 0.20 ± 0.04 (3.6%) b | 5.07 ± 0.09 (92.8%) ab | 0.03 ± 0.02 (0.5%) ab | 5.46 ± 0.13 ab |
| t10 | 0.12 ± 0.01 (2.4%) b | 0.29 ± 0.03 (5.9%) a | 0.59 ± 0.09 (12.0%) a | 3.93 ± 0.37 (79.6%) b | 0.01 ± 0.00 (0.1%) b | 4.94 ± 0.47 b | |
| tR | 0.17 ± 0.02 (1.5%) a | 0.09 ± 0.00 (0.8%) ab | 0.29 ± 0.07 (2.5%) ab | 10.92 ± 1.48 (94.9%) a | 0.04 ± 0.01 (0.3%) a | 11.50± 1.55 a | |
| M28 | t0 | 0.21 ± 0.06 (1.6%) a | 0.04 ± 0.02 (0.3%) b | 0.14 ± 0.04 (1.1%) b | 12.82 ± 2.04 (96.7%) a | 0.04 ± 0.01 (0.3%) a | 13.25 ± 2.15 a |
| t10 | 0.17 ± 0.04 (3.5%) a | 0.33 ± 0.02 (6.9%) a | 0.41 ± 0.08 (8.4%) a | 3.94 ± 0.42 (81.1%) b | 0.01 ± 0.02 (0.1%) b | 4.86 ± 0.48 b | |
| tR | 0.09 ± 0.02 (1.0%) b | 0.07 ± 0.00 (0.7%) ab | 0.20 ± 0.03 (2.0%) ab | 9.24 ± 0.62 (96.0%) ab | 0.03 ± 0.00 (0.3%) ab | 9.63 ± 0.60 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sales, E.; Cañizares, E.; Pereira, C.; Pérez-Oliver, M.A.; Nebauer, S.G.; Pavlović, I.; Novák, O.; Segura, J.; Arrillaga, I. Changing Temperature Conditions during Somatic Embryo Maturation Result in Pinus pinaster Plants with Altered Response to Heat Stress. Int. J. Mol. Sci. 2022, 23, 1318. https://doi.org/10.3390/ijms23031318
Sales E, Cañizares E, Pereira C, Pérez-Oliver MA, Nebauer SG, Pavlović I, Novák O, Segura J, Arrillaga I. Changing Temperature Conditions during Somatic Embryo Maturation Result in Pinus pinaster Plants with Altered Response to Heat Stress. International Journal of Molecular Sciences. 2022; 23(3):1318. https://doi.org/10.3390/ijms23031318
Chicago/Turabian StyleSales, Ester, Eva Cañizares, Catia Pereira, María Amparo Pérez-Oliver, Sergio G. Nebauer, Iva Pavlović, Ondřej Novák, Juan Segura, and Isabel Arrillaga. 2022. "Changing Temperature Conditions during Somatic Embryo Maturation Result in Pinus pinaster Plants with Altered Response to Heat Stress" International Journal of Molecular Sciences 23, no. 3: 1318. https://doi.org/10.3390/ijms23031318