
Abstract
The Hawk–Dove model has been used to explain how aggression evolves in animal species. However, testing this model with experimental data has proven challenging because the two model parameters, V and C, are difficult to measure. We propose a novel consumer-resource model that overcomes these difficulties, and we explore the dynamical behavior of the model. Furthermore, by studying a series of consumer-resource models with interactions based on the Hawk–Dove game, we make new predictions for how the level of aggression may change with the richness of the environment, animal mortality, and the amount of time spent fighting.



Similar content being viewed by others
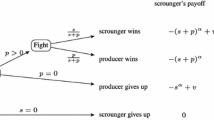

Availability of Data and Materials
Not applicable.
Notes
We assume that N is large.
In both cases, we also calculate the proportion of Hawks in the population as the population size approaches its limit.
We numerically solved model (7) for 20,000 time steps. Then, using the final abundances as initial conditions, we numerically solved the model for an additional 20,000 time steps. Using this solution, we integrated the average proportion of Hawks over this time.
This prediction assumes that the value of the resource V is less than the cost of fighting C. Otherwise, the Hawk–Dove game predicts all individuals will be aggressive.
A similar model was studied in Křivan et al. [17]. In Sect. 4 of their paper, they interpreted the Hawk–Dove model as a model of contest competition. Resources were nesting sites, and the number of nesting sites was fixed. They showed that Hawks would outcompete Doves, thus occupying all of the available nesting sites. This underscores a major difference between how aggression might evolve under scramble competition or contest competition.
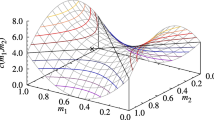
Bistability was also observed in Auger et al. [3] and can be related to similar dynamics in models of intraguild predation [cf., 5, 23, 13, 16] if we conceive of Hawks as intraguild predators and Doves as intraguild “prey.” This is because both Hawks and Doves compete for the common resource, but Hawks also obtain energy from Doves through pair-wise interactions when they capture the resource that a Dove is handling. In both situations, Hawks reduce the Doves’ net growth rate, albeit in our case Hawks do not kill Doves but they collect the Doves’ resource. Holt et al. [11] showed that intraguild predators and intraguild prey can coexist only if the resource environmental carrying capacity is intermediate and the intraguild predators are inferior to the intraguild prey at exploiting a common resource. Otherwise, the intraguild prey is outcompeted. We observe a similar pattern in our model too. First, Hawks are weaker competitors because of the additional cost C. Second, Fig. 1b shows that when m is large enough (i.e., \(m>C/8\)), Doves are outcompeted from the system at high environmental resource carrying capacity. At lower mortality rates we observe bistability at high values of K where Doves are either outcompeted, or they coexist with Hawks at the equilibrium proportion \(p_2\).
Exploitation and interference competition are also known, respectively, as scramble and contest competition.
References
Amarasekare P (2002) Interference competition and species coexistence. Proc Biol Sci 269(1509):2541–2550
Armstrong RA, McGehee R (1980) Competitive exclusion. Am Nat 115:151–170
Auger P, de la Parra RB, Morand S, Sánchez E (2002) A predator–prey model with predators using hawk and dove tactics. Math Biosci 177 &178:185–200
Bailey CJ, Andersson LC, Arbeider M, Bradford K, Moore JW (2019) Salmon egg subsidies and interference competition among stream fishes. Environ Biol Fishes 102(6):915–926
Brown DH, Ferris H, Fu S, Planta R (2004) Modeling direct positive feedback between predators and prey. Theor Popul Biol 65:143–152
Chesson P (2000) Mechanisms of maintenance of species diversity. Annu Rev Ecol Syst 31:343–366
Collie J, Granela O, Brown EB, Keene AC (2020) Aggression is induced by resource limitation in the monarch caterpillar. iScience 23(12):101791
Dubois F, Giraldeau L-A (2005) Fighting for resources: the economics of defense and appropriation. Ecology 86:3–11
Gause GF (1934) The struggle for existence. Williams and Wilkins, Baltimore
Hofbauer J, Sigmund K (1998) Evolutionary games and population dynamics. Cambridge University Press, Cambridge
Holt RD, Polis GA (1997) A theoretical framework for intraguild predation. Am Nat 149:745–764
Huxley J (1966) A discussion on ritualization of behaviour in animals and man. Philos Trans R Soc Lond Ser B Biol Sci 251:249–271
Křivan V (2000) Optimal intraguild foraging and population stability. Theor Popul Biol 58:79–94
Křivan V, Cressman R (2017) Interaction times change evolutionary outcomes: two player matrix games. J Theor Biol 416:199–207
Křivan V, Cressman R (2022) The asymmetric Hawk–Dove game with costs measured as time lost. J Theor Biol 547:111162
Křivan V, Diehl S (2005) Adaptive omnivory and species coexistence in tri-trophic food webs. Theor Popul Biol 67:85–99
Křivan V, Galanthay T, Cressman R (2018) Beyond replicator dynamics: from frequency to density dependent models of evolutionary games. J Theor Biol 455:232–248
Lach L (2005) Interference and exploitation competition of three nectar-thieving invasive ant species. Insectes Soc 52(3):257–262
Levin SA (1970) Community equilibria and stability: an extension of the competitive exclusion principle. Am Nat 104:413–423
Łomnicki A (1988) Population ecology of individuals. Princeton University Press, Princeton
Łomnicki A (2008) In: Jørgensen SK, Fath BF (eds) Competition and behavior, vol 1. Elsevier, Amsterdam, pp 695–700
Maynard Smith J, Price GR (1973) The logic of animal conflict. Nature 246:15–18
Mylius SD, Klumpers K, de Roos AM, Persson L (2001) Impact of intraguild predation and stage structure on simple communities along a productivity gradient. Am Nat 158:259–276
Okuyama T (2012) Behavioral states of predators stabilize predator–prey dynamics. Thyroid Res 5:605–610
Roeleke M, Johannsen L, Voigt CC (2018) How bats escape the competitive exclusion principle—seasonal shift from intraspecific competition drives space use in a bat ensemble. Front Ecol Evol 6:101
Taylor PD, Jonker LB (1978) Evolutionarily stable strategies and game dynamics. Math Biosci 40:145–156
Funding
This project has received funding from the European Union’s Horizon 2020 research and innovation programme under Grant Agreement No. 955708, project EvoGamesPlus (Evolutionary games and population dynamics: from theory to applications).
Author information
Authors and Affiliations
Contributions
TEG, VK and RC were involved in the conceptualization and writing—review and editing. TAR contributed to the numerical analysis and review and editing.
Corresponding author
Ethics declarations
Conflict of interest
None of the authors has any financial or personal conflicting interests.
Ethical Approval
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the topical collection “Evolutionary Games and Applications” edited by Christian Hilbe, Maria Kleshnina and Kateřina Staňková.
A Model (6) analysis
A Model (6) analysis
First, we reparametrize model (6). This reparametrization is important in stability analysis given below because it allows us to derive in Mathematica conditions for local equilibria stability analytically. This is not possible for the original parametrization of the model given in the article.
Let \(R = \alpha \tilde{R},\,N=\gamma \tilde{N}, t = \delta \tau \). Then,
Letting \(\delta = \frac{1}{C},\,\gamma =\frac{C}{\lambda },\) and \(\alpha = \frac{C}{e\lambda }\), we obtain after simplification,
Now we reparametrize with \(\tilde{r} = \frac{r}{C},\,\tilde{K} = \frac{e\lambda K}{C},\,\tilde{m}=\frac{2}{C} m\) to yield
Finally, we can drop the tildes to obtain the system
The new state variables correspond to the old ones by \(\tilde{R} = \frac{e \lambda }{C}R,\,\tilde{N} = \frac{\lambda }{C}N\).
The advantage of model reparametrization is that model (11) can be analyzed using Mathematica. First, we calculated equilibria and then, using the Routh-Hurwitz criterion we derived conditions for the theoretical local asymptotic stability using command Reduce of Mathematica 12. The results are given in Table 4.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Galanthay, T.E., Křivan, V., Cressman, R. et al. Evolution of Aggression in Consumer-Resource Models. Dyn Games Appl 13, 1049–1065 (2023). https://doi.org/10.1007/s13235-023-00496-w
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13235-023-00496-w